Навигация
Эндотоксины — липополисахариды. Они определяют антигенную специфичность бактерий; форму колоний и являются причиной эндотоксикоза
218815
знаков
4
таблицы
0
изображений
4. Эндотоксины — липополисахариды. Они определяют антигенную специфичность бактерий; форму колоний и являются причиной эндотоксикоза.
Факторы патогенности диареегенных E.coli контролируются не только хромосомными генами, но и генами, привносимыми плазмидами или умеренными, конвертирующими фагами. Особенно важную роль играют плазмиды. У E.coli обнаружены плазмиды различных категорий: Col-, R-, F-, Hly-, Ent-плазмиды (контролируют синтез энтеротоксинов); плазмиды, кодирующие синтез факторов адгезии и т. п. Все это указывает на то, что патогенные варианты кишечной палочки могут возникать в результате распространения среди них плазмид и умеренных фагов. В зависимости от наличия тех или иных факторов патогенности диареегенные E.coli разделяют на следующие категории:
энтеропатогенные E.coli (enteropathogenic — EPEC)
энтеротоксигенные E.coli (enterotoxigenic — ETEC),
энтероинвазивные E.coli (enteroinvasive — EIEC),
энтерогеморрагические E.coli (enterohaemorrhagic — EHEC).
провизорная группа энтероадгерентных E.coli (enteroadherence — ЕАЕС).
Энтеропатогенные кишечные палочки (ЭПКП). С них в 1950-х гг. начался современный этап в изучении диареегенных эшерихий. Этим объясняется универсализм термина - энтеропатогенность, который в равной мере можно распространить на все диареегенные эшерихии. ЭПКП, к которым относятся представители около 20 0-серогрупп (чаще 055, 0111, 0119, 0127, 0128), вызывают поражение тонкого кишечника. Заражение возможно и через продукты питания. Ранее заболевание именовалось «токсической диспепсией». В основе диарейного синдрома лежит перестройка внутриклеточного гомеостаза энтероцитов, которая возбуждается сигналами с клеточных рецепторов, оккупируемых адгезинами ЭПКП. Итогом является сглаживание микроворсинок, что ведет к нарушению всасывания жидкости, инициируя диарею секреторного типа, возможно образование эрозий. К числу главных адгезинов относятся закодированные в хромосоме белки наружной мембраны и производные плазмиды обязательного атрибута ЭПКП. В целом по своим патогенетическим последствиям ЭПКП-адгезия уникальна, получив название механизма «прикрепления-сглаживания» (англ. attaching-effacing). ЭПКП-диарею причисляют к энтеритам, хотя патогенетически это неверно. Поражение энтероцитов носит функциональный характер и, подобно энтеротоксигенной диарее, не сопровождается воспалительной реакцией.
Энтеротоксигенные кишечные палочки (ЭТКП) обнаружены среди представителей более 70 0-групп, чаще среди 06, 078, 0128 и 0153 (в сумме они составляют более половины ЭТКП). ЭТКП продуцируют токсины, нарушающие баланс между секрецией и всасыванием жидкости эпителиальными клетками тонкого кишечника. Избыток воды и электролитов в просвете кишечника вызывает диарею. Возможны различные формы заболевания — от легкой диареи до холероподобной интоксикации («малая холера»). Болеют дети и взрослые. Заражение возможно и через продукты питания. Распространение бактерий ограничено поверхностью слизистой оболочки, энтероциты не инвазируются и структурно не повреждаются. Об этом, в частности, свидетельствуют отсутствие воспалительной реакции в стенке кишечника и водянистые испражнения без примеси слизи и крови. Такой тип диареи называют «секреторным», противопоставляя инвазивному варианту, который наблюдается при деструкции кишечного эпителия. Примерно 25% штаммов ЭТКП образуют токсин типа LT и около 70% штаммов ЭТКП продуцируют токсин типа ST. Токсинообразованию предшествует закрепление ЭТКП на энтероцитах чаще всего тонкого отдела кишечника, где среди факторов, обеспечивающих избирательную колонизацию энтероцитов, лучше всего изучены CFA/I, CFA/II и CF/IV. Они примерно поровну распределены между ЭТКП-штаммами и представляют собой белок, структурно оформленный в виде пилей/фимбрий. Фимбрии ЭТКП животных (К88, К89, 987Р) неидентичны адгезинам «человеческих» ЭТКП. Это объясняет причину антропонозности энтеротоксигенных колиинфекций. В настоящее время считают, что ЭТКП животных не способны колонизировать тонкий кишечник человека, что справедливо и для других категорий диареегенных эшерихий. Энтеротоксины ЭТКП человека и животных структурно тоже неидентичны, хотя и близки по механизму действия. CFA закодированы в плазмидах «по соседству» с генами энтеротоксинов, и лишь одновременная экспрессия cfa- и tox-генов обеспечивает ЭТКП-вирулентность. Без колонизирующих факторов энтеротоксины патогенетически инертны точно так же, как CFA-адгезины без токсигенности. Штаммы ЭТКП, лишенных CFA составляют около 20% выделяемых культур.
Энтероинвазивные кишечные палочки (ЭИКП) вызывают колитический синдром, практически не отличимый от шигеллезной дизентерии, - диарею инвазивного типа (кровянистые испражнения с примесью слизи и лейкоцитов). Сходство с шигеллами распространяется и на механизмы, обеспечивающие инвазивность. Их контролируют плазмидные гены, гомологичные плазмидам вирулентности шигелл. Подобно шигеллам, они проникают и размножаются в эпителиоцитах нижнего отдела подвздошной и толстого кишечника, размножаясь и повреждая их, индуцируют воспаление и изъязвление слизистой оболочки. Энтероинвазивность ограничена представителями нескольких 0-групп: 028,0112,0124,0136,0143, 0144, 0152, 0164. Они отвечают за небольшой процент всех случаев бактериальной дизентерии, значительно уступая шигеллам. ЭИКП обычно поражают детей - в организованных коллективах. Взрослые страдают редко — во время пищевых вспышек. Об инвазивности штамма судят по способности вызывать кератоконъюнктивит при закапывании бактериальной взвеси в конъюнктивальный мешок морской свинки или по цитопатическому эффекту в культурах эпителиальных клеток. Таксономические сомнения устраняют за счет расширения набора биохимических тестов
Энтерогеморрагические кишечные палочки (ЭГКП). По показателям вирулентности, это, пожалуй, наиболее оснащенная разновидность эшерихии. Большинство ЭГКП относится к серогруппе 0157 (серотип 0157:Н7), реже к 026, 0111, 0145. ЭГКП колонизируют толстый кишечник (особенно слепую кишку), вызывая геморрагический колит. В отличие от шигеллезных и ЭИКП-поражений кровянистые испражнения бедны лейкоцитами. Заболевание сопровождается общей интоксикацией (тошнота, рвота), а в самых тяжелых случаях внекишечными симптомами - гемолитическая анемия, острая почечная недостаточность. Инициирующая роль принадлежит агрессивной, интиминзависимой адгезии. За ней следует продукция токсинов. Токсины разрушают клетки эндотелия мелких кровяносных сосудов, а образование сгустков крови и выпадение фибрина вызывает нарушение кровотока, кровотечение. Приводит к ишемиии и некрозу в клеточной стенке, которые оказывают местное действие (геморрагический колит). Действие продуктов данных штаммов вызывает системные эффекты (поражение почек, центральной нервной системы, внутрисосудистый гемолиз). Известно несколько цитотоксических факторов ЭГКП, которые легко обнаружить по гемолизу, хотя это лишь одно из проявлений их мембранотропной (цитолитической) активности.
Кроме гемолизинов, продуцируемых штаммами многих 0-групп, ЭГКП секретируют специфические цитолизины. Их называют «веротоксинами» (по токсическому действию в культуре клеток линии «Vero») или «шигоподобными токсинами», подразумевая сходство (прежде всего нейротропность) с токсином Shigella dysenteriae. Описаны и особые, энтерогеморрагические гемолизины. ЭГКП-цитолизины имеют плазмидную природу. Плазмидные гены необходимы и для колонизации кишечника. Подобно ЭПКП, энтерогеморрагические штаммы несут плазмиду, продукты которой включаются в патогенетически значимую адгезию.
В отличие от других колиинфекций ЭГКП-эшерихиозы являются зоонозами. Наиболее вероятным источником заражения считается крупный рогатый скот. Об этом свидетельствует ряд вспышек 0157-эшерихиоза, зафиксированных в Канаде, США и Японии. Заражение происходит при употреблении в пищу мяса (после недостаточной термической обработки), а также сырого молока.
Энтероагрегирующие (энтероадгезивные) кишечные палочки (ЭАКП). Дополнительным поводом к усложнению классификации диареегенных эшерихий явились морфологические особенности их адгезии в культурах эпителиальных клеток человека (НЕр-2 и HeLa). Один из вариантов адгезии, напоминающий кирпичную кладку (stacked-brick), обусловлен фиксацией на клеточной мембране бактериальных агрегатов. Такие штаммы выделены в самостоятельную категорию диареегенных эшерихий и считаются одним из возбудителей диарейного (особенно упорно протекающего) синдрома у детей. ЭАКП колонизируют различные отделы кишечника и продуцируют несколько цитотоксинов. Структурная основа и патогенетическая значимость ЭАКП-адгезивности неизвестны. Непонятно и то, почему, обладая широкими способностями к колонизации кишечного эпителия, ЭАКП поражают преимущественно толстый кишечник.
ВНЕШНИЕ ПРИЗНАКИ БОЛЕЗНИ (КЛИНИКА). Внешнее проявление признаков эширихиоза у людей не столь разнообразна, как при сальмонеллезе, и не имеет характерных особенностей. Время после приема зараженной бактериями пищи до проявления признаков болезни (инкубационный период) составляет 2-4 часа, редко 12-24 часа. Начинаются боли в животе, тошнота, рвота, частый стул (понос), чуть повышается температура (37,0°–37,4°С). В более тяжелых случаях - головная боль, озноб, общая слабость, у детей иногда побледнение губ, судороги. Длительность заболевания до 3-х дней. Смертельные случаи редки.
Нельзя забывать, что бактерии кишечной палочки могут попадать в продукты питания не только посредством людей, с ними контактировавших (повара, продавцы, грузчики и т.п.), а также посредством кошек, собак и сельскохозяйственных животных.
Поэтому рассмотрим внешнее проявление признаков колибактериоза у них. У больных животных наблюдаются: повышение температуры тела, с возможным ее снижением перед смертью животного, потеря аппетита, озноб, испарина, слабая подвижность, быстрая потеря веса, сильный понос кашицеобразными или водянистыми фекальными массами желтого цвета. При очень слабом проявлении болезни понос может и не проявляться, а если болезнь затягивается, то наблюдается опухание суставов. У поросят в возрасте 1-2 месяца болезнь может протекать в отечной форме, которая характеризуется появлением отеков в области глаз, повышенной возбудимостью или параличом конечностей. Диагноз ставится по результатам бактериологических исследований с выделением бактерии – кишечной палочки и определением ее антигенной формулы с помощью серологической типизации.
ПРОФИЛАКТИКА ПИЩЕВОГО ОТРАВЛЕНИЯ. Учитывая, что опасность представляют не только те животные, которые болеют и признаки заболевания ясно выражены, но и носители данной бактерии, внешне ничем себя не проявляющие, важно соблюдать меры профилактики и предупреждения заноса данного микроба в пищевые продукты. Надо в первую очередь перед убоем животного, присмотреться к его состоянию. Когда оно угнетено, вяло, с неохотой двигается, то необходимо подождать с забоем, выяснить причину подобного поведения. Надо помнить, что инструменты для убоя и разделки туши должны быть чистыми, прокипяченными. Если это не сделать, то те микробы, которые находятся на них, при разделке туши попадут на мясо, внутренние органы и будут там размножаться, накапливаясь в полученных продуктах питания. После того, как зарезали животное, необходимо как можно быстрее удалить из туши желудочно-кишечный тракт. Установлено, что уже через 40 минут после убоя, микробы, содержащиеся в кишечнике, проникают в кровь, внутренние органы и мышечную ткань. При жизни, у здорового животного, подобного не происходит, так как этому препятствует работа иммунной системы. Место убоя животного должно быть чистым, особенно от пыли и лучше будет если пол, стены или станок будут протерты слабым раствором какого-либо дезинфектанта. Если его нет, то крепким раствором поваренной соли. Это требование объясняется тем, что во внешней среде (пыль, солома и т.п.) очень много бактерий. Следующее требование, которое рекомендуют выполнять, связано с купанием, обтиранием животного перед забоем. На одном квадратном сантиметре кожного покрова может находиться от сотни миллионов до миллиарда микроорганизмов. При снятии шкуры возможен ее контакт с мясной тушей или внутренними органами, и тогда часть микроорганизмов попадет на них с последующим размножением и заражением получаемых пищевых продуктов. Для того, чтобы избежать подобного обсеменения микробами и рекомендуют обработку кожного покрова. Лучше всего применять 0,5% раствор хлорамина, но и обработка животных просто теплой водой (20-25°С) способствуют снижению содержания бактерий до 200-500 тысяч на см2 , что в сотни, тысячи раз меньше исходного количества. Еще одна рекомендация, выполнение которой предотвратит попадание микробов в мышечную ткань, а значит и в получаемые из нее пищевые продукты. Желательно проведение убоя животного в спокойных для него условиях. Не надо гонять животное по двору, бить его, подобные действия вызывают у него стресс. Доказано, что в стрессовом состоянии уже при жизни животного из кишечника в мышечную ткань, внутренние органы, кровь, проникают микроорганизмы.
Соблюдение вышеперечисленных рекомендаций позволяет в значительной степени избежать обсеменения микроорганизмами продуктов питания, получаемых из животного сырья (мясо, мясные полуфабрикаты, колбасно-копченые изделия и т.п.).
САНИТАРНАЯ ОЦЕНКА МЯСА И МЯСОПРОДУКТОВ, ПОЛУЧАЕМЫХ ОТ БОЛЬНЫХ КОЛИБАКТЕРИОЗОМ ЖИВОТНЫХ ИЛИ В СЛУЧАЕ ВЫДЕЛЕНИЯ ИЗ НИХ БАКТЕРИЙ КИШЕЧНОЙ ПАЛОЧКИ. Из 167 известных в настоящее время серовариантов кишечной палочки, около 100 серовариантов относятся к той группе, что вызывает болезнь. Этим и определяют подход к оценке мяса. Во-первых, если забили больное животное, то необходимо отправить образцы мяса для исследования в бактериологический отдел лаборатории. На основании результатов этих исследований и будет проводиться санитарная оценка. Так как ,при любом заболевании организм животного ослаблен, то может быть наслоение одной болезни на другую. При выделении сальмонеллезной палочки, все мясо направляется на проварку. В случае выделения какого-либо другого микроорганизма, вызывающего инфекционную болезнь, санитарная оценка проводится в зависимости от вида этого микроорганизма или тяжести вызываемой им болезни. Во-вторых, если выделена только бактерия кишечной палочки, то санитарная оценка мяса будет зависеть от того, какую кишечную палочку выделили. В случае выделения энтеропатогенного штамма (вызывающего отравление) мясо отправляется на проварку в течение 2,5 часов при весе каждого куска не более 2-2,5 кг, а внутренние органы утилизируют, в пищу не допускают. Готовую пищевую продукцию уничтожают. При выделении кишечной палочки, относящейся к неболезнетворным серовариантам, мясо можно направить для производства колбас или других продуктов, при приготовлении которых используют термическую (температурную) обработку, внутренние органы необходимо проваривать.
Без предварительной температурной обработки реализовывать, использовать такое мясо нельзя. Если из мышечной ткани и лимфоузлов микроорганизмы не выделены, то мясо реализуют без ограничения. Надо помнить то, что по внешнему виду ни мясо, ни мясопродукты, содержащие в себе кишечную палочку, ничем не отличаются от мяса, полученного от здорового животного. Органолептические изменения отсутствуют.
Пищевой сальмонеллез
Сальмонеллез – инфекционное заболевание млекопитающих и птиц, протекающее весьма разнообразно, но чаще всего в виде типичных токсикоинфекций. Основные пути передачи — пищевой, реже — контактный.
ИСТОРИЧЕСКАЯ СПРАВКА. В 1885 году американские микробиологи Сальмон и Смит выделили из мяса болевших свиней бактериальную культуру, вошедшую в последствии в обширную группу микроорганизмов, названных в честь одного из первооткрывателей - сальмонеллы. Поворотным пунктом в изучении вопроса о мясных отравлениях явилось открытие Гертнером во Франкенхаузене (Тюрингия) в 1888 г. бактерий, послуживших причиной отравления мясом. После употребления в пищу мяса коровы, забитой вследствие заболевания катаром кишечника (мясо было признано ветеринарным врачом доброкачественным за отсутствием в нем и в организме коровы каких-либо существенных изменений), заболело острым катаром желудка и кишок 58 человек в 25 семьях. Заболевание начиналось через 2—3 часа после употребления мяса. Наиболее тяжело заболели евшие сырое мясо мужчина, съевший 800 г, умер. Гертнеру удалось выделить возбудителя отравления как из мяса коровы, так и из селезенки умершего. Он назвал его Bact. enteritidis. Этот случай послужил основой научного исследования отравлений мясом. Уже годом позже в г. Гота подобные бактерии обнаружил Ионе в мясе коровы с тяжелым воспалением вымени. Потребление мяса этой коровы привело к массовому заболеванию, причем четверо заболевших умерли. Открытие Гертнера в дальнейшем подтвердилось и при отравлениях мясом в Гауштадте в 1891 г., в Моорзееле, Румфлетхе и Роттердаме -в 1892 г. и другими многочисленными случаями в последующие годы.Однако вскоре открылись широкие горизонты для новых исследований, и вопрос о причинах отравлений мясом, по мере. увеличения числа исследований, становился не проще и ясней, а все запутанней и трудней. 1893 г. при отравлении жареным мясом, двух вынужденно убитых коров, Флюгге и Кенше из Института гигиены в Бреславле обнаружили в мясе возбудителя, который был подобен бактериям гертнер, но не во всём тожественен им; он был назван бактерией флюгге-кенше, или бактерия бреслау. В 1898 г. при отравлении мясом в Эртрикке (Фландрия) Нобель нашел возбудителя, подобным же образом отличающегося от бактерии Гертнера; в литературе он получил название бактерии эртрикк, или бациллы де нобеля. Позднее выяснилось, что он идентичен бреславльской палочке. Новым этапом в истории вопроса явилось открытие паратифозных бактерий. В 1896 г. Ашар и Бенсод, а несколькими годами позже Шоттмюллер наблюдали у людей заболевание, протекавшее с клинической картиной тифа. Из крови и испражнений больных были выделены бактерии, которые агглютинировались сывороткой больных в довольно больших разведениях, в то время как тифозные бактерии агглютинировались этой сывороткой лишь немного сильнее, чем сывороткой крови здоровых людей. Вновь обнаруженные бактерии отличались культурально и биологически от тифозных.. торы назвали вызванное ими заболевание паратифом. Возбудитель его известен теперь под названием бактерии паратифа В. Буква В была добавлена, чтобы отличить бактерию паратифа В от бактерии паратифа А, которую открыл в 1898 г. Гвин, а подробнее описали Брион и Кайзер. Вскоре, в 1903 г., Траутман описал отравление фаршем из конины в Дюссельдорфе и отнес возбудителя к паратифозным бактериям, хотя его агглютинирующие свойства были ближе к свойствам бреславльских бактерий. С этого времени укоренился термин «паратифозная группа» или просто «паратифозные бациллы».
На сегодняшний день зарегистрировано свыше 2200 серовариантов сальмонелл, из них у человека выделено более 700. Первоначально виды сальмонелл называли чаще в соответствии с местом выделения (Moskow, London, Branderburg и др.), по имени ученых (Гертнера, Вирхова и др.) или в зависимости от заболеваний, вызываемых у животных (мышиный тиф и др.). Бактерии рода сальмонелл чаще всего встречаются в кишечнике животных и человека, а также во внешней среде.
МОРФОЛОГИЯ И КУЛЬТУРАЛЬНЫЕ СВОЙСТВА. Эти бактерии представляют собой палочки с закругленными концами, иногда овальной формы, не очень длинные. В мазках, окрашенных по методу Грама, они будут грамотрицательные. Все они подвижны, за исключением S.gallinarium, S.pullorum и не образуют спор и капсул. На питательных средах большинство сальмонелл образует небольшие, диаметром 2—4 мм, колонии. Ряд сероваров (S. abortusequi, S. abortus ovis, S. typhisuis) образует более мелкие колонии, диаметром около 1 мм. Колонии сальмонелл на питательном агаре прозрачно - голубоватого цвета. При посеве на среду Эндо они слега розоватые, прозрачные; на среде Плоскирева — бесцветные, выглядят более плотными и мутноваты. На висмут сульфит агаре колонии всегда черного цвета, с металлическим блеском. Питательная среда под колонией окрашена в черный цвет. Ряд сероваров, в частности S. рагаtyphi С, образует на этой среде светлые, зеленоватые колонии.
Ферментативные свойства сальмонелл, послужившие одним из критериев выделения подродов, различаются даже в пределах одного серовара. Большинство сальмонелл аэрогенны, но есть исключения, например S. typhi, которая никогда не продуцирует газа. Образование сероводорода и отсутствие продукции индола, как и аэрогенность, — характерные свойства сальмонелл. Однако, по данным F. Kauffmann, имеется не менее 30 сероваров, образующих индол.
УСТОЙЧИВОСТЬ этих бактерий к воздействию часто встречаемых физических и химических факторов высокая. Они могут длительное время выживать в пыли, навозе, сухом кале, почве, воде и животных кормах, сохраняя вирулентность. В воде открытых водоёмов и питьевой воде они живут 11-120 сут, в морской воде - 15-27 сут, в почве — 1-9 мес, в комнатной пыли — 80-547 сут,. В сухом кале крупного рогатого скота сальмонеллы сохраняются до 4-х лет. Сальмонеллы переживают в течение длительного времени на керамических, металлических и стеклянных предметах соответственно 70, 55 и 43 дня. Наиболее устойчива S. typhimurium, остающаяся жизнеспособной на тканях и на бумаге до года. Выдерживают 5-6 кратное замораживание и оттаивание. Выживание сальмонелл в различных пищевых продуктах неодинаково. Мясо, молоко - являются благоприятной средой для размножения. В сыре, масле, они не размножаются, но не теряют жизнеспособность. В мясе, хранящемся на холоде, эти микроорганизмы также сохраняются, а при повышении температуры до 5°С и выше начинают размножаться. В мороженом мясе живут до 3-х лет, в колбасных изделиях – до 130 дней, яйцах – до 13 месяцев, яичном порошке – до 9 месяцев, на замороженных овощах и фруктах — 0,5-2,5 мес. При температуре ниже 5°С роста сальмонелл в течение 3-х недель не наблюдают; при 5-8°С в первые два дня рост мало замечен, а после 3-4 дней идет интенсивное размножение. При 8-13°С интенсивный рост отмечается уже через 12 часов. Из замороженных яичных желтков сальмонеллы выделяются после 13-ти месяцев хранения этих продуктов при –20°C. Соление и копчение слабо действуют на них. В соленом и копченом мясе, при содержании 12-19% поваренной соли бактерии погибают через 75 дней, при 10-11%-ном содержании соли они остаются жизнеспособными даже после 80 дней. В мясном рассоле с содержанием 29% соли при 6-12°С сальмонеллы сохраняют жизнеспособность от 4 до 8 месяцев, а в свиных кишках, содержащих 22% соли, в течение 6 месяцев. Сальмонеллы чувствительны к воздействию кислот. В свинине, массивно инфицированной сальмонеллами, бактерии отмирают через 14 дней после введения в толщу мяса 10%-ной уксусной кислоты и последующей заливки его этой кислотой. Хранение сырого молока при 18-20°С и кислотности 26°Т (T- кислотность по Тернеру) сальмонеллы в нем выживают в течение 11 суток, а при снижении температуры до 5-8°С – до 20 суток. В простокваше с кислотностью 85°Т и хранящейся при температуре 0-4°С данные бактерии способны к размножению в течение 2-х суток. В простокваше с кислотностью в 130-160°Т эта способность сохраняется в течение 24-х часов. Сальмонеллезные бактерии достаточно устойчивы к воздействию высокой температуры. Нагревание при 70 °С выдерживают в течение 30 мин. Устойчивость к высокой температуре повышается, когда сальмонеллы находятся в пищевых продуктах, особенно в мясе. Мясо, зараженное сальмонеллами и заложенное в холодную воду, становится стерильным в кусках весом не более 400,0 граммов при толщине кусков 19 см и варке в течение 2,5 час; а при закладке в кипяток стерильность за тот же срок варки достигается лишь в кусках весом до 200,0 граммов, при толщине их 5,0-5,5 см. Пастеризация молока при 85°С в течение 30 минут вызывает инактивацию сальмонелл. Эти бактерии инактивируются под действием 20%-ного раствора кальцинированной соды, 5%-ного раствора щелочи, 5-6%-ного раствора нафтализола, 1%-ного раствора формальдегида, 20%-ного раствора хлорной извести.
СЕРОЛОГИЧЕСКАЯ ХАРАКТЕРИСТИКА САЛЬМОНЕЛЛЕЗНЫХ БАКТЕРИЙ. Каждый серотип сальмонелл имеет, характерный только для него, набор определенных антигенных факторов. Этот набор, составляющий антигенную структуру серотипа, складывается из сочетания О- и Н-антигенов микроба. В соответствии с предложением Кауфмана и Уайта (1934) все подроды сальмонелл по 0-антигену делят на серологические группы, а по Н-антигену — на серотипы. 0-антиген расположен на поверхности микробной клетки и представляет собой фосфолипидо-полисахаридный комплекс, включающий 60% полисахарида, 20—30% липида и 3,5—4,5% гексозамина (углевода). Этот комплекс термостабилен, не разрушается при кипячении в течение 2,5 ч., а автоклавированием при120°С в течение 30 мин. Чувствителен к действию формалина, но резистентен к этанолу. Деление сальмонелл на серологические группы производят в соответствии с содержанием тех или иных 0-антигенов, при этом отдельные антигенные факторы обозначают арабскими цифрами. Обнаружено 65 О - антигенов Наличие у сальмонелл Н-антигена определяется жгутиками и выделение серологических типов обусловлено Н-антигенами. Они термолабильны, разрушаются нагреванием при температуре 75-100°С, под воздействием НС1, этанола и протеолитических ферментов, но устойчивы к формалину. Н – антигены у сальмонелл могут встречаться в двух серологических фазах: первой («специфическая») и второй («неспецифическая»). Антигены 1-й фазы ( их обнаружено более 80 вариантов) обозначают прописными буквами латинского алфавита, 2-й фазы ( 9 вариантов)— арабскими цифрами или прописными латинскими буквами с арабскими цифрами. Сальмонеллы, в которых Н-антигепы представлены двумя фазами, называют двухфазными в отличие от монофазных, имеющих только антигенные факторы 1-й фазы. Состав антигенов, определяющий антигенную формулу салъмонелл, не является стабильным. Это представляется тем, что благодаря развитию различных вариаций Н- и О-антигенов у салъмонелл могут возникать закономерные изменения этих антигенов. Как уже упоминалось, современная серологическая классификация сальмонелл насчитывает свыше 2200 серовариантов. Помимо указанных выше антигенов, у сальмонелл выявлены поверхностные К (Vi) -антигены – антигены вирулентности. Они представляют собой белково-полисахаридный комплекс с выраженными антигенными свойствами. Он термолабилен, чувствителен к действию НС1 и этанола, полностью разрушается при кипячении в течение 10 мин, частично инактивируется под действием фенола и при температуре 60 0C в течение 1 ч. Температурный оптимум для него 37 °С. F. Kauffmann обнаружил М-антиген (1935, 1936) и Т-антиген (1956, 1957) сальмонелл. Несмотря на многочисленность и вариабельность антигенов при серологической идентификации сальмонелл, принимают во внимание три основных антигена — О, Н, Vi. Во всех лабораториях мира используется диагностическая схема Кауфмана — Уайта, построенная на анализе этих трех антигенов. В соответствии с особенностями структуры 0-антигенов в этой схеме выделены примерно 50 0-групп. Каждая группа объединяет большее или меньшее количество сероваров, расположенных в алфавитном порядке по обозначению 1-й фазы их Н-антигена. Определение серовара сальмонелл начинается с изучения выделенной культуры, в реакции агглютинации на стекле с поливалентной (ABCDE) адсорбированной 0- сывороткой и далее с монорецепторными Н-сыворотками. После определения антигенов 1-й фазы анализируют антигены 2-й, что позволяет установить антигенную формулу серовара. При отсутствии агглютинации с поливалентной ABCDE 0-сывороткой и положительной реакции с поливалентной сывороткой редких групп, проводят дополнительные биохимические тесты с целью определения принадлежности к подродам сальмонелл. При положительном решении далее исследуют штамм с помощью моновалентных О-сывороток редких групп и Н-сывороток.
Для установления источников инфекции, эпидемиологических связей между отдельными заболеваниями, дифференциации привозных случаев от местных, определение антигенной формулы распространенных штаммов сальмонелл дополняется внутрисероваровым разделением. С этой целью определяют биовары и фаговары. Известно, что в пределах одного серовара встречается большее или меньшее число стабильных биохимических вариантов бактерий Выделение биоваров основано на ферментативной активности штамма по отношению к углеводам. В частности, у S. typhimurium имеется 25 стабильных биоваров, а биовары S. enteritidis и S. dublin имеют даже отдельные названия (denysz, chaco, essen, асага, coehi).
Важное значение для идентификации, имеет определение чувствительности штамма к сальмонеллезному 0-бактериофагу. Он лизирует 97,5% штаммов сальмонелл. Поэтому в качестве эпидемиологических маркеров используют и фаготип сальмонелл. В настоящее время существуют схемы для фаготипирования, основанные на специфичности бактериофага по отношению к микробу-хозяину. Известны типовые фаги к S. typhimurium, S. enteritidis, S. dublin и др. В настоящее время у S. typhimurium различают 120 фаготипов, причем ряд типов имеет ограниченное распространение. Особый интерес у специалистов вызывает фаготип 29 S. typhimurium. Считается, что этот фаготип распространяется только среди людей (контактно-бытовой путь), вызывает тяжелые формы сальмонеллеза, устойчив к антибиотикам. Вместе с тем ряд часто встречающихся сероваров устойчив к этому фагу (S. derby, S. tennessee, S. ariatum, S. london).
Определение антигенной формулы у выделенной из пищевого продукта бактерии играет значительную роль для выявления истинного источника сальмонелл.
БОЛЕЗНЕТВОРНОСТЬ (ПАТОГЕННОСТЬ) сальмонелл обусловлена действием 2-х типов токсических веществ, выделяемых данными бактериями. К первому типу токсинов - экзотоксинов относятся вещества, выделяемые при жизни бактерий. Второй тип токсических веществ - эндотоксины образуются в результате разрушения бактериальной клетки и представляют собой ее структуру.
Сальмонеллы способны образовать термоустойчивые токсические вещества, которые вызывают заболевания и гибель при внутривенном, внутрибрюшинном введении. Токсины могут образовываться этими бактериями в пищевых продуктах. В сыром мясе, зараженном сальмонеллами и хранившемся при 16-20°С, эти вещества накапливаются от 2-х до 7 дней; при 0‑4°С они не образуются даже в течение месяца.
У сальмонелл имеются факторы адгезии и колонизации, факторы инвазии; они имеют эндотоксин и, наконец, они, по крайней мере S.typhimurium и некоторые другие серотипы, могут синтезировать два типа экзотоксинов:
1) термолабильные и термостабильные энтеротоксины типа LT и ST;
2) шигаподобные цитотоксины.
1.Особенностью токсинов является внутриклеточная локализация и выделение после разрушения бактериальных клеток. LT сальмонелл имеет структурное и функциональное сходство с LT энтеротоксигенных E.coli и с холерогеном. Он устойчив в диапазоне рН 2,0-10,0. Токсинообразование у сальмонелл сочетается с наличием у них двух факторов кожной проницаемости:
а) быстродействующего - продуцируется многими штаммами сальмонелл, термостабилен (при 100 °С сохраняется до 4-х ч.), действует в течение 1-2 ч;
б) замедленного — термолабилен (разрушается при 75 °С в течение 30 мин), вызывает эффект (уплотнение кожи кролика) через 18-24 ч после введения.
2.Цитотоксин, продуцируемый сальмонеллами, термолабилен, его цитотоксическое действие проявляется в угнетении синтеза белка энтероцитами. Обнаружено, что отдельные штаммы сальмонелл могут одновременно синтезировать LT, ST и цитотоксин, другие — только цитотоксин.
Вирулентность сальмонелл зависит также от обнаруженной у них плазмиды с м.м. 60 МД, утрата ее значительно снижает вирулентность бактерий. Предполагается, что появление эпидемических клонов сальмонелл связано с приобретением ими плазмид вирулентности и R-плазмид.
РАСПОСТРАНЁННОСТЬ. Гастроэнтериты это заболевания более известны как сальмонеллёзы — группа полиэтилогичных острых инфекционных болезней человека, животных и птиц с фекально-оральным механизмом передачи.
Наиболее опасными источниками пищевых токсикоинфекций являются животные. Вспышки токсикоинфекций чаще всего связаны с употреблением мяса, инфицированного сальмонеллами, — до 70-75%, в том числе до 30% мяса скота вынужденного забоя. Вынужденному забою часто подвергают животных, находящихся в агональном состоянии. У ослабленных животных сальмонеллы легко проникают из кишечника в кровь, а через нее — в мышцы, обусловливая прижизненное инфицирование мяса. Большую роль в эпидемиологии сальмонеллезов играют водоплавающие птицы и их яйца, а также куры, их яйца, и другие птицепродукты. Сальмонеллы могут попасть в яйцо непосредственно во время его развития, но могут легко проникнуть и через неповрежденную скорлупу. На долю яиц и птицепродуктов приходится более 10% (но существует тенденция роста заболеваемости, обусловленной увеличением доли мяса птицы и птицепродуктов в пищевом рационе людей), на долю молока и молочных продуктов — около 10%, и на долю рыбопродуктов — около 3-5% всех вспышек сальмонеллезов. Несколько реже наблюдают поражения, вызванные употреблением инфицированных моллюсков, черепах, рыбы, овощей и фруктов. Заболевания регистрируют повсеместно, в тёплый сезон (с мая по октябрь) отмечают подъём заболеваемости.
Современная эпидемиология сальмонеллезов характеризуется постоянным ростом заболеваемости людей и животных и увеличением числа серотипов сальмонелл, вызывающих эти заболевания. С 1984 по 1988 год в Англии число случаев сальмонеллезов возросло в 6 раз. Однако специалисты ВОЗ полагают, что истинное число случаев сальмонеллезов остается неизвестным. По их мнению, выявляется не более 5-10% инфицированных лиц. Одной из основных причин роста заболеваемости сальмонеллезом, является инфицирование пищевых продуктов, при их производстве, в результате широкого распространения сальмонелл на объектах внешней среды и на обрабатывающих предприятиях, куда поступают животные, у которых сальмонеллез протекает в скрытой форме. Ухудшение экологической обстановки, загрязнение сальмонеллами различных объектов внешней среды, сточных и поверхностных вод, широкая циркуляция сальмонелл среди диких птиц и грызунов ведут к постоянному инфицированию кормов. Это одна из главных причин широкой циркуляции сальмонелл среди животных. Применение корма, содержащего переработанные побочные продукты животного происхождения и очень часто зараженного сальмонеллами - вторая причина роста сальмонеллёза.
Несмотря на постоянное увеличение числа серотипов сальмонелл, выделяемых от людей и животных, по-прежнему до 98% всех случаев сальмонеллезов обусловлено сальмонеллами групп А, В, С, D и Е, в первую очередь — S.typhimurium и S.enteritidis (до 70-80% случаев заболеваний).
Другой немаловажной особенностью современной эпидемиологии сальмонеллезов является установление роли человека как источника заражения сальмонеллами. Заражение человека от больного или бактерионосителя возможно не только через пищу, в которой сальмонеллы находят хорошие условия для размножения, но и контактно-бытовым путем. Этот способ заражения приводит к широкому распространению бессимптомного бактерионосительства. Крупная водная эпидемия сальмонеллезной инфекции в 1965г. в г. Риверсайде (США), обусловленная S.typhimurium (заболело около 16 тыс человек), показала, что заражение сальмонеллами возможно не только через пищу, но и через воду.
Сальмонеллезом болеют преимущественно молодые животные всех видов (крупный рогатый скот, свиньи и др.). В этом возрасте животные чрезвычайно восприимчивы к данной инфекции и выздоровление их не всегда заканчивается полным освобождением от возбудителей. Заболевание людей может быть вызвано различными серовариантами бактерий. Разделение сальмонелл на болезнетворных для человека и животных не имеет оснований. В настоящее время описаны болезни людей, обусловленные такими сальмонеллами, которые долго считались патогенными только для животных и птиц (S.pullorum, S. gallinarum, S. abortus). С другой стороны, все чаще описываются случаи выделения у животных чисто «человеческих» штаммов, особенно S.рaratyphi. Определенная приспособляемость отдельных типов сальмонелл к тому или иному виду имеется, но эта избирательность отнюдь не определяет абсолютной монопатогенности данного возбудителя для человека и животных.
ВНЕШНИЕ ПРИЗНАКИ БОЛЕЗНИ (КЛИНИКА). В медицинской практике наблюдают следующие клинические формы заболеваний, вызываемых сальмонеллами: гастроинтестинальную, дизентериеподобную, холероподобную, тифоподобную, гриппоподобную, септическую, нозапаразитическую и субклиническую (скрытое бактерионосительство). Инкубационный период для всех форм в среднем 12-24 ч., редко 6-8 ч., а иногда 2-3 дня. Массивное обсеменение сальмонеллами пищевого продукта обусловливает пищевую токсикоинфекцию, при которой основные симптомы связаны с поступлением возбудителя в кровь в большом количестве, его распадом и высвобождением эндотоксина. В основе сальмонеллезной диареи лежит колонизация сальмонеллами энтероцитов. После прикрепления в тонком кишечнике сальмонеллы внедряются между ворсинками и, прикрепляясь к плазмолемме энтероцитов, колонизируют ее, повреждают микроворсинки, вызывают слущивание энтероцитов и умеренное воспаление слизистой оболочки. Освобождающийся энтеротоксин вызывает диарею, а цитотоксин — гибель клеток. Сальмонеллы размножаются на плазмолемме, но не в энтероцитах, а происходит их инвазия через эпителий в подлежащие ткани слизистой оболочки, они транспортируются через нее в макрофагах, поступают в лимфу и кровь, вызывая бактериемию и генерализацию инфекционного процесса.
ГАСТРОИНТЕСТИНАЛЬНАЯ ФОРМА заболевания (встречается в 80-90% случаев) характеризуется следующими признаками: повышение температуры тела до 38-40°С, нередко с ознобом, тошнота, рвота, жидкий стул, иногда с примесью крови, слизи и зелени (при дизентериеподобной форме), боли в животе, обложенный и сухой язык, повышенная жажда, головная боль, боли в мышцах и суставах и нередко желтуха. Болезнь длится 3-7 дней, затягиваясь в отдельных случаях на более длительный срок.
ХОЛЕРОПОДОБНАЯ ФОРМА сальмонеллеза встречается редко и протекает почти со всеми признаками холеры: проффузный понос, имеющий вид «рисового отвара», обильная изнуряющая рвота, высокая температура, сильный озноб, слабость, боли в животе, судороги, резкое исхудание, цианостичность лица. Болезнь длится несколько дней. Нередко смерть наступает в первые сутки при явлениях упадка сердечной деятельности и отека легких.
ТИФОПОДОБНАЯ ФОРМА может начинаться с обычного острого гастроэнтерита и после кажущегося выздоровления через несколько дней переходит в тифоподобную форму, которая, однако часто заканчивается в более короткие сроки (8-10 дней), чем обычный брюшной тиф.
ГРИППОПОДОБНАЯ ФОРМА характеризуется тем, что явления со стороны желудочно-кишечного тракта маскируются симптомами поражения органов дыхания, причем нередко такая форма диагностируется как кишечный грипп. Заболевание протекает при следующих признаках: высокая температура (38-40°С), озноб, общее недомогание, мышечно-суставные боли в конечностях и пояснице, конъюктивит, насморк, явления фарингита и ангины. Диспептические признаки выражены слабо и отступают на второй план.
СЕПТИЧЕСКАЯ ФОРМА сальмонеллеза может протекать в виде септицемии или септикопиэмии. Нередко наблюдают местные септические процессы с локализацией очагов во внутренних органах и различных тканях: эндокардиты, перикардиты, пневмонии, холециститы, остеомиелиты и артриты (особенно у детей), абсцессы, нагноительные процессы в различных местах организма.
НОЗОПАРАЗИТИЧЕСКАЯ ФОРМА сальмонеллеза является вторичным заболеванием, наслаивающимся на первичное и возникающем в результате эндогенного проникновения сальмонелл в организм, ослабленный первичным заболеванием.
Диагноз сальмонеллеза у людей базируется на данных внешне проявляемых симптомах, эпидемиологии и серо-бактериологических исследований.
Смертность при сальмонеллезе незначительная (1-2%), зависит от тяжести болезни, возрастного состава и др.
Как уже указывалось, большую опасность представляет мясо вынужденно убитых животных. В 33-85% случаев, заболевания сальмонеллезом связано с использованием мяса от таких животных. Источником заболевания людей сальмонеллезом являются убойные животные, как больные так и скрытые бактерионосители сальмонелл. Клиническое выздоровление при сальмонеллезе не всегда совпадает по времени с освобождением организма животного от возбудителей. Переболевшие животные длительное время (до нескольких лет) являются носителями и выделителями сальмонелл. Будучи внешне совершенно здоровыми, эти животные выделяют бактерии с калом, мочой, с носовой слизью и слюной, рассеивая их во внешнюю среду. Такие животные опасны как для молодняка, так и для взрослых животных. Они представляют большую опасность и для людей, так как мясо этих животных и приготовленные из него продукты могут явиться причиной сальмонеллезных заболеваний. Кроме того, возможно заражение рабочих при убое животных и разделке туш.
У животных сальмонеллоносителей, возбудитель локализуется преимущественно в желчном пузыре, в печени, почках, реже в других органах. У таких животных, при послеубойной экспертизе, обнаружить в тканях и органах какие-либо патологические изменения не удается. Поэтому продукты убоя животных - сальмонеллоносителей могут быть по ошибке выпущены без ограничений, а в мясе и органах таких животных содержится небольшое количество бактерий, которые могут начать быстро размножаться, если будут нарушены режимы хранения мяса и мясопродуктов. В этих случаях мясопродукты могут явиться одним из серьезных источников сальмонеллеза среди людей.
Свиньи, по сравнению с другими животными чаще являются источником сальмонелл инфекции. Обладая слабой восприимчивостью к сальмонеллезной инфекции, они во многих случаях являются скрытыми носителями этой бактерии, без проявления внешних симптомов.
Среди крупного рогатого скота и среди свиней отмечается большое многообразие циркулирующих серовариантов сальмонелл с преобладанием наиболее опасного для человека вида бактерии S.typhimurium.
Сальмонеллез возможен у животных как осложнение при инфекционных, инвазионных и незаразных заболеваниях. Наиболее часто болезнь возникает у животных с желудочно-кишечными заболеваниями, с поражением органов дыхания у истощенных послеродовыми заболеваниями, поражениями суставов и вымени, плевритами, перикардитами, различными абсцессами и флегмонами, обширными травматическими повреждениями, инфицированными ранами, при инфекционных болезнях.
Сальмонеллез, возникший на фоне уже имеющегося какого-либо болезнетворного процесса, не имеет специфической клинической картины и характерных послеубойных изменений, которые затушевываются изменениями, обусловленными основным заболеванием.
Неясность клинических признаков, патологоанатомических изменений в этих случаях может быть причиной свободного выпуска в пищу мяса, неблагополучного по сальмонеллезной инфекции и опасного для здоровья людей. Для предотвращения такой опасности необходимо тщательное обследование животных перед убоем и при обнаружении клинических признаков, указывающих на возможность сальмонеллезной инфекции, убивают таких животных отдельно от здоровых или в конце общего убоя, и обязательно проводят бактериологическое исследование туш и органов. Кроме того, при послеубойной ветеринарной экспертизе, необходимо внимательно учитывать все малейшие патологоанатомические изменения в органах и туше и в неясных случаях всегда направлять их на бактериологическое исследование.
ПРИ ПРЕДУБОЙНОМ ОСМОТРЕ КРС в зависимости от длительности заболевания обнаруживают слабость, сонливость, дрожь, шаткость походки, потерю аппетита, повышение температуры или лишь незначительный подъем ее, проффузный понос (фекальные массы жидкие, желтоватого или серо-желтоватого цвета с неприятным запахом, а иногда с примесью крови, пузырьков газа и крупозными пленками), нередко конъюнктивит с обильным слезотечением и серозным истечением из носа. При подостром течении болезни у телят наблюдают симптомы пневмонии, проявляющиеся внешне сухим кашлем, ускоренным дыханием, одышкой. Видимые слизистые оболочки бледные. Телята больше лежат, часто изогнув шею и закинув голову на спину. Нередко у больных животных, особенно при хроническом течении болезни, наблюдают опухание суставов, главным образом запястного и скакательного. При сальмонеллезе у взрослых животных обнаруживают желтуху, у коров - массовые аборты.
Из послеубойных изменений в тушах крупного рогатого скота обнаруживают следующее. Слизистая оболочка сычуга воспалена и усеяна мелкими кровоизлияниями. Слизистая кишечника, особенно заднего отдела тонких кишок, тоже усеяна точечными кровоизлияниями и обильно покрыта слизью. Обнаруживают иногда точечные кровоизлияния на слизистой прямой кишки. Кишечные лимфоузлы набухшие, сочные с кровоизлияниями. В них нередко наблюдают некротические поражения в виде желтовато-белых очажков. На серозных оболочках, в лимфатических узлах туши и на слизистой мочевого пузыря обнаруживают кровоизлияния. Селезенка сильно увеличена, края закруглены, капсула сильно напряжена, пульпа вишнево-красного цвета. Почки полнокровны и усеяны многочисленными кровоизлияниями, особенно под капсулой.
САЛЬМОНЕЛЛЕЗ СВИНЕЙ вызывается различными серотипами сальмонелл.
В подавляющем большинстве случаев наблюдается сальмонеллез поросят в возрасте 1,5-6 мес. Заболевание сопровождается большой смертностью (75%) заболевших, чаще в 1,5–2,5-месячном возрасте. При предубойном осмотре у больных свиней наблюдают сильное угнетение, отсутствие аппетита, повышение температуры до 41-42°С, слабость, конъюнктивит, понос со зловонным жидким калом, иногда с примесью крови, слизи, признаки колик, фиолетово-красное или синеватое цианотическое окрашивание кожи на ушах («синеухие поросята»), шее, нижней части груди, живота и пахов, затрудненное дыхание, кашель, одышку. При хроническом течении болезни отмечают резкое истощение, струпьевидную экзему на коже, опухание суставов, понос с желтовато-зеленым зловонным калом. В ряде случаев появляются признаки хронического поражения легких в виде продолжительных приступов кашля. Сальмонеллез у взрослых свиней чаще протекает хронически, без определенного симптомокомплекса. При послеубойном исследовании туш свиней наблюдают изменения в желудочно-кишечном тракте. В толстых кишках наблюдают некроз слизистой. Участки некроза превращаются в отрубевидную массу, Иногда некрозы слизистой отмечают в желудке и тонком отделе кишечника, особенно в подвздошной кишке. На слизистой оболочке толстых кишок (при хроническом течении) обнаруживают плоские язвы, дно которых покрыто хлопьевидными массами. Язвы обычно окружены валиком, состоящего из грануляционной ткани. Некротизированная поверхность слизистой толстого отдела, особенно слепой кишки, покрыта струпьевидной массой или творожистой кашицей. При хроническом течении болезни изменения в кишечнике сопровождаются увеличением лимфатических узлов и появлением в них мелких творожистых очагов желтого цвета. Легкие, преимущественно передние доли, воспалены, иногда имеются уплотненные участки с последующим творожистым перерождением; легочные лимфатические узлы претерпевают при этом те же изменения, что и узлы кишечника. Селезенка остается без изменений. Окончательный диагноз ставят на основании бактериологического исследования.
САЛЬМОНЕЛЛЕЗ ПТИЦ это собирательный термин, включающий заболевания, вызываемые S.pullorum (пуллороз), S.gallinarum (тиф птиц), S.arizonae (аризонос) и др. Иногда эти заболевания называют паратифом. Зачастую клинические признаки не всегда выражены.
Большая зараженность сальмонеллами отмечена у водоплавающих птиц. Возбудителей болезни обнаруживают не только в мясе и органах сальмонеллезных птиц, но и в их яйцах, как на яичной скорлупе, так и в содержимом яйца. Для полного освобождения яиц от сальмонелл следует варить утиные яйца в течение 13 мин., а гусиные - 14 мин. ( с момента закипания воды). Мясо, органы и яйца птиц, инфицированные возбудителями, представляют большую опасность для человека, вызывая тяжелые заболевания.
ПРОФИЛАКТИКА ПИЩЕВОГО ОТРАВЛЕНИЯ. Мясо инфицируется главным образом при жизни животного. Следовательно, одним из источников сальмонеллеза у людей являются больные животные. Однако значительную роль в заболевании играет и послеубойное инфицирование мяса. Основной причиной послеубойного обсеменения мяса сальмонеллами является убой животных-сальмонеллоносителей, которые могут обсеменить туши животных сальмонеллами в процессе убоя и переработки.
Послеубойное (экзогенное) обсеменение мяса и мясных продуктов бактериями происходит при разделке туш, обваловке, жиловке, при приготовлении фаршей, от рук и инструментов рабочих, оборудования и инвентаря, используемых при обработке мяса. Источником возбудителя инфекции могут быть инфицированные вода и лед, вспомогательные материалы, используемые в мясном производстве, жалящие насекомые(мухи, тараканы), птицы, грызуны (мыши, крысы) и наконец, бактериовыделитель – человек. Значительное обсеменение может происходить в тех случаях, когда имеют место нарушения санитарных правил хранения мяса и мясопродуктов.
САНИТАРНАЯ ОЦЕНКА МЯСА И ПОЛУЧАЕМЫХ ИЗ НЕГО ПРОДУКТОВ. Внешний вид мяса и мясопродуктов, зараженных сальмонеллами, как правило, не изменен, и не вызывает никаких подозрений. В основе санитарной оценки мяса и мясопродуктов, зараженных сальмонеллами, лежит патогенность этих бактерий для человека при попадании в пищеварительный тракт с пищей. Поэтому мясо сальмонеллезных животных нельзя выпускать из предприятия без ограничения.
По действующему ветеринарному законодательству при обнаружении сальмонелл (независимо от вида) в органах или туше животного, органы направляют на техническую утилизацию или уничтожают, а тушу проваривают или переваривают на мясные хлеба и консервы по установленному режиму. Готовые мясные изделия, обсемененные сальмонеллами, направляют на техническую утилизацию или уничтожают.
Отравления людей, вызываемые бактериями «Протей»Микроорганизмы рода Proteus так же, как и кишечная палочка, широко распространены во внешней среде, в каловых массах животных и человека, что и является источником обсеменения пищевых продуктов.
ИСТОРИЧЕСКАЯ СПРАВКА. Впервые микроорганизм этой группы был выделен и описан в 1885 году Хаузер (Hauser) при исследовании гниющего мяса. А свое название бактерия получила в честь сына Посейдона -водяного божества Протея – способного менять свой облик, так как данная бактерия меняла внешние проявления своего роста на питательных средах.
МОРФОЛОГИЯ БАКТЕРИЙ И КУЛЬТУРАЛЬНЫЕ СВОЙСТВА. Как и кишечная палочка, этот микроорганизм в мазках, окрашенных по методу Грама - грамотрицателен. Палочки размерами 1-3х0,4-0,8 мкм, капсул и спор не имеют. Протеям в наибольшей степени свойственен полиморфизм с образованием нитевидных и кокковидных форм. Протеи — факультативные анаэробы, хемоорганотрофы. Температурный оптимум — 37°С. Оксидаза-отрицательны, каталаза-положительны. Гидролизуют мочевину. Расщепляют тирозин, растут на средах KCN, восстанавливают нитраты. Обитают в кишечнике многих видов позвоночных животных, почве, сточных водах и разлагающихся органических остатках. Типовой вид — Р. vulgaris. Для бактерий характерен феномен роения (особенно выраженный на агаре меньшей плотности при температуре 20-22 °С); внесение в среду ингибиторов (соли жёлчных кислот, высокие концентрации NaCL, мочевина) обусловливает рост в неподвижной 0-форме. Подвижность бактерий используют для выделения чистой культуры посевом в конденсационную воду скошенного агара по Шукёвичу. Протеи растут на простых средах в диапазоне температур 10-43 °С (оптимальная — 35-37 °С), оптимум рН -— 7,2-7,4. Колонии протеев в 0-форме округлые, полупрозрачные и выпуклые; Н-формы дают сплошной рост. Пигментов не образуют; рост сопровождается гнилостным запахом. На среде Плоскирева формируют желтовато-розовые колонии (в зоне роста среда подщелачивается и желтеет). На висмут-сульфитном агаре через 48ч образуют серо-коричневые колонии (под ними формируется черно-коричневая редукционная зона). На агарах МакКонки и Эндо формируют бесцветные колонии. Вызывают помутнение жидких сред. Подвижность более выражена при температуре 37оC, практически все штаммы имеют жгутики, значит, в их антигенной формуле есть Н-антиген.
АНТИГЕННАЯ ХАРАКТЕРИСТИКА. Антигенная структура идентична таковой у прочих энтеробактерий, у протеев выделяют О-, Н- и К- антигены. Для серологической идентификации определяют структуру О- и Н- Аг, обозначаемых арабскими цифрами. В настоящее время антигенная формула включает в себя более 49 О-антигенов и 19 Н-антигенов, всего 110 сероваров.. В отличие от бактерий кишечной палочки, бактерии протея нельзя с твердой уверенностью разделить на болезнетворные и неболезнетворные штаммы по показаниям антигенной формулы. Хотя отмечено, что при кишечных заболеваниях людей наиболее часто выделяют палочки протея определенной серологической группы по О-антигену. В любом случае определение антигенной формулы микроорганизма, выделенного от больного человека и сравнивание ее с антигенной формулой микроорганизма того же вида, но выделенного из пищевого продукта, мяса (послужившего сырьем для приготовления этого продукта), людей, контактировавших с данным продуктом, позволяет проследить и выявить источник заражения людей.
УСТОЙЧИВОСТЬ. Данный микроорганизм выдерживает нагревание в 65°С в течение 30 минут. При 70°С он инактивируется (но в водных растворах), есть данные, что он выдерживает первоначальные режимы пастеризации молока. Замораживание слабо влияет на его жизнеспособность. При хранении мяса в режиме -13°С эти микробы выделяли из него в течение 6 месяцев. Бактерии достаточно устойчивы к высоким концентрациям хлористого натрия (поваренной соли). Они отмирали только после 17% концентрации, таким образом рассолы с более низкой концентрацией не убивают протей. Данный микроорганизм очень устойчив и к кислотному воздействию, сохраняя свои жизненные функции в диапазоне рН 3,5 - 12,0. Поэтому, попадая на мясо от здорового животного, где величина рН в пределах 5,8 - 6,2 палочка протея, в отличии от других болезнетворных микроорганизмов, может размножаться. Даже такой метод хранения пищевого продукта, как маринование т.е. [КСВ1] понижение кислотности до величин 4,5-5,0, не убивают эту бактерию.
БОЛЕЗНЕТВОРНОСТЬ (ПАТОГЕННОСТЬ). Факторы патогенности многочисленны, важнейшие из них - способность к «роению», фимбрии, бактериальные протеазы и уреаза, гемолизины и гемагглютинины. Механизм патогенности у бактерий данного рода такой же, как и у всех микроорганизмов семейства энтеробактерий (сальмонелл, кишечной палочки и т.д.). При экспериментальном введении в организм мимо желудочно-кишечного тракта (внутривенно, внутрибрюшинно) протейный эндотоксин оказывает сильное ядовитое действие. Однако при введении через рот с пищевыми продуктами, сам токсин самостоятельное токсическое действие практически не оказывает или оно не столь значительно, как при ином способе введения. Для возникновения пищевой токсикоинфекции, необходимо наряду с наличием эндотоксина, обсеменение пищевого продукта живыми бактериями.
ВНЕШНИЕ ПРИЗНАКИ БОЛЕЗНИ (КЛИНИКА). Проявление данной болезни у людей достаточно своеобразно. Инкубационный период (от момента попадания в организм до проявления болезни) укладывается в 4 - 20 часов. Болезнь может иметь внезапное начало, сопровождаться схваткообразными, режущими болями в кишечнике, тошнотой и рвотой, поносом. В фекальных и рвотных массах иногда отмечают наличие крови. В тяжелых случаях наблюдают судороги, падение сердечной деятельности, побледнение слизистых. Длительность болезни до 5 дней. Смертельный исход возможен в пределах 1,5 %. Необходимо отметить, что палочка протея в сочетании с возбудителем другого инфекционного агента, способствует повышению болезнетворного процесса. У животных этот микроорганизм вызывает воспалительные процессы в желудке, кишечнике. Достаточно часто данное заболевание у животных встречается совместно с другой инфекцией, осложняя его. Различные патологические (патология - отклонение от нормы) процессы, вызванные любой причиной (раны, маститы, бронхопневмонии и тому подобное), способствуют проникновению палочки протея из кишечника в кровь.
ПРОФИЛАКТИКА ДАННОГО ПИЩЕВОГО ОТРАВЛЕНИЯ аналогична тем мероприятиям, что и при эширихиозах.
САНИТАРНАЯ ОЦЕНКА МЯСА из которого выделена палочка протея такая же, как и при выделении сальмонеллезной палочки. Мясо рубят на куски по 2 кг, варят 2,5 часа и только после этого используют для реализации или на пищевые цели. Внутренние органы уничтожают. Органолептические изменения у пищевых продуктов, в которых содержится палочка протея, присутствуют. Появляется неприятный запах, похожий на запах тухлых яиц или плесени, возможно появление слизи. В таком случае мясо уничтожают, т.к. на пищевые цели оно непригодно.
Кишечный иерсиниозПищевое отравление человека, называемое кишечным иерсиниозом, вызывается бактериями Yersinia enterocolitica.
ИСТОРИЧЕСКАЯ СПРАВКА И РАСПОСТРАНЁННОСТЬ. За последние 50 лет отмечено расширение круга микроорганизмов, которые обуславливают пищевые отравления. Наряду с известными, хорошо изученными болезнями появляются “новые”, вызываемые микроорганизмами, расцениваемыми ранее как условно патогенные. К числу таких болезней относится кишечный иерсиниоз людей и животных, возбудителем которого является бактерия Yersinia enterocolitica.Первые сведения о возбудителе кишечного иерсиниоза получены в США, где с 1923 по 1957 год было выявлено от людей около 15 штаммов бактерий, но в то время они были отнесены к атипичным вариантам псевдотуберкулёзного микроба и не рассматривались как причина заболевания. В 1964 г. во Франции и Бельгии появились сообщения о выделении возбудителя иерсиниоза (первоначально названного Pasteurella Х ) при мезентеральном лимфадените, аппендиците, энтерите. В последующие годы бактерию выделяли и за пределами Европе в США в 1965 году, Канаде и Южной Африке в 1966, Бельгийском Конго в 1967, Бразилии в 1969, Японии 1972, Иране 1976 году (H.H.Mollaret, 1995). Одно из первых крупных зарегистрированных пищевых отравлений, вызванных данными иерсиниями, зарегистрировано в Японии в 1972 году. Тогда 180 школьников вместе с учителем были поражены заболеванием с характерными симптомами бактериального пищевого отравления. Следующая крупная вспышка подобного пищевого отравления была описана у 220 школьников США в 1976 году. Причем 36 школьников были госпитализированы, а 16-ти из них произведена апендиктотомия, прежде чем был правильно поставлен диагноз заболевания. В дальнейшем кишечный иерсиниоз, как пищевая инфекционная болезнь людей, появилась в Западной Европе – Франции, Бельгии, Голландии, Германии, Швейцарии. Позднее она регистрировалась в Швеции, Финляндии, Венгрии. По предложению W.Frederiksen бактерии вызывающие болезнь названы Yersinia enterocolitica, что и было принято Международным подкомитетом по таксономии в 1972г. В настоящее время кишечный иерсиниоз занимает второе место по значимости после сальмонеллезов( В.И. Покровский 1985), а вспышки пищевых отравлений, вызванных данным возбудителем болезни регистрируются чаще, чем бактериальная дизентерия. В России кишечный иерсиниоз занимает второе место среди пищевых зоонозов, после сальмонеллёза (Черкасский Б.Л., Подунова Л.Г., Акулова Н.К., 1995).
Опасность кишечного иерсиниоза усугубляется чрезвычайной распространённостью возбудителя Yersinia enterocolitica в природе. Как отмечают ряд исследователей (Ленченко Е.М., Куликовский А.П., Павлова И.Б., 1998; Зыкин А.Ф., 1998; Покровский В.И.,1986; Ющенко Г.В., 1984) бактерии Yersinia enterocolitica выделены практически от всех видов млекопитающих, птиц, рыб, земноводных, моллюсков и насекомых. Кроме того, Yersinia enterocolitica обнаружены в воде, почве, сточных водах, продуктах животного происхождения. Возбудители кишечного иерсиниоза широко распространены среди диких животных, в частности, среди грызунов, а также среди домашних животных. Из домашних животных наиболее часто в качестве источника иерсиниозной инфекции рассматриваются собаки. Gutman с соавт. (1975) предположили, что межсемейная вспышка энтерита, вызванного Y.enterocotitica у 16 из 21 человека (четыре семьи) в Северной Калифорнии, была связана с заболеванием собаки и ее щенков. Инфицированность собак, по данным Y. Yanagawa et al.(1978).достигает 6% . Известны многочисленные случаи выделения иерсиний от кошек, причем в большей части патогенных для человека сероваров 03 и 09 (Ващенок Г.И. и др.1983). Микроорганизмы вида Y.enterocolitica выделены у коз и оленей, однако значение этих животных в развитии эпизоотии иерсиниоза пока мало изучено. Кишечным иерсиниозом болеют и сельскохозяйственные животные. По мнению S.Christensen (1980), бактерионосительство свиней в различных хозяйствах значительно колеблется - от 6 до 49%. Эти животные бывают не только носителями иерсиний, у них регистрируют и клинически выраженное заболевание, проявляющееся диареей или генерализацией инфекции, оканчивающейся гибелью животного. Бактерионосительство крупного рогатого скота иерсиниями невелика: по данным разных авторов, она составляет от 0,7 до 7,8% (Ващенок Г.И. и др.1983). В большинстве случаев иерсинии высевают из испражнений внешне здоровых животных, а также у тех, у которых отмечали диарею. Тяжелое течение иерсиниоза, сопровождается эндокардитом, абортом и гибелью плода. Наряду с сельскохозяйственными и домашними животными источниками иерсиниозной инфекции могут служить обитатели дикой природы. Shaygani с соавт. (1979) исследовали экскременты 1426 особей различных млекопитающих, птиц, рептилий, беспозвоночных и выделили 147 штаммов Y.enterocotitica. Бадаева Е.Я и др. (1988) сообщили, что при обследовании 1200 обезьян был выделен 21 штамм Y.enterocolitica серовара 03. Особую опасность представляют маститы вызываемые иерсиниями, что сказывается на контаминации пищевых продуктов данной бактерией. Инфицированность сырого молока достигает 20%, пастеризованного 7-9%, мяса 5% (М.В. Дулатова 1989). По данным Ющенко Г.В. и др. (1969), указанные микроорганизмы выделяются из сырого молока 10 -12 % проб (молочная кухня, молокозавод), пастеризованного в 6 %, из молока от частных коров в 6,6 % проб. По данным Vidon D.,Delmas C., во Франции в 1981г при исследовании молока с помощью оригинальной обогатительной среды Yersinia enterocolitica были выявлены в 81,4% проб. Из молока и молочных продуктов Y.enterocolitica выделена в Австралии, Чехословакии. Японии, Дании, Аргентине, Германии, России. Shiemann (1978) сообщил, что микроорганизмы Y.enterocolitica были выделены в 9,2 % из 65 проб сыра. Необходимо отметить, что существующие методы пастеризации молока полностью обеззараживают его от Y.enterocolitica. Появление же бактерий в пастеризованном молоке связано с технологическими нарушениями или обусловлено вторичной контаминацией пастеризованного молока при соприкосновении с технологическим оборудованием, не прошедшим установленной санитарной обработки. В.Г. Кузнецов, В.Н. Багрянцев 1992г. в своих исследованиях показали, что в 35%, т.е. в каждой 3-й пробе пастеризованного молока, обнаруживали иерсинии. Как показали их исследования, активность бактерий сохраняется даже при разведении пробы молока в 4 раза. Выводы их исследований таковы: а) пастеризованное молоко, поступающее в продажу, в 35,8% случаев контаминировано апатогенными видами Y.enterocolitica, преимущественно биовара 1, являющегося обитателем внешней среды; б) контаминация пастеризованного молока носит вторичный характер, т.к. происходит во время розлива молока в тару, случайно загрязненную иерсиниями из окружающей среды; в) пастеризованное молоко, контаминированное иерсиниями различных биоваров и сероваров, играет роль важного фактора распространения носительства иерсиний среди населения и потенциально таит в себе опасность стать причиной заражения патогенными иерсиниями. Способность иерсиний длительное время сохраняться в продуктах животного и растительного происхождения, размножаться при низких температурах, а также неприхотливость к условиям обитания способствуют накоплению их в пищевых продуктах. Пищевой путь передачи иерсиниозной инфекции человеку является ведущим, поскольку именно с ним связано большинство спорадических групповых заболеваний и вспышек иерсиниоза. Заражение человека происходит в результате употребления в пищу сырых или недостаточно термически обработанных продуктов (мясо, молоко, фрукты, овощи). Ющенко Г.В. и др. (1969) утверждают, что частота обнаружения иерсиний с овощей и фруктов колебалась от 2,7 % до 5, 2 % и от 2,1 до 3,2 % соответственно. При этом 44 % заболевших связывали свое заболевание с употреблением сырого молока, 27,7 % - овощей и фруктов, 6,5 % - с употреблением других продуктов. Активность пищевого фактора передачи подтверждает и идентичность сероваров штаммов, выделенных от больных, из пищевых продуктов и внешней среды. Всего выделено 247 штаммов Y.enterocotitica (03 -24,3 %, 09-35,4 %). Мясо и мясные продукты так же могут служить путями распостранения иерсиниозной инфекции в случае получения их от больных животных (бактерионосителей) или вторично через различные предметы во время убоя и переработки. Leistner et al (1975) в Германии обнаружили, что 28,9 % проб куриного мяса, 34,5 % свинины, 10,8 % говядины были контаминированы Y.enterocotitica. Inoue Kurose (1975) в Японии выделили Y.enterocolitica из говядины. Schiemann (1980) в Канаде выделил Y.enterocolitica из свинины и продуктов ее переработки, причем наибольшее количество штаммов было выявлено в свиных языках и фарше. На убойных пунктах Дании Christensen (1982) выделил серовар 08 Y.enterocolitica в смывах из миндалин здоровых свиней. Seetye et al (1979) в США выделили несколько штаммов Y.enterocolitica из упакованных в вакууме кусков говядины и баранины. Бактерии были обнаружены в высушенном мясе. В Бельгии при исследовании 29 образцов свиных языков, 50 - мездры, 25-свиного мяса, 54 образцов миндалин - патогенные штаммы иерсиний были выделены соответственно в 96,5; 16; 26 и 61% случаев. Есть случаи инфицирования колбасы (М.В.Дулатова 1989), т.к. технология ее приготовления включает этап выдерживания мясного фарша в растворе поваренной соли в течении 3-х суток в холодильнике.
Большое значение, как фактору передачи иерсиниозной инфекции, ряд авторов придают и другим пищевым продуктам. Например, штаммы Y.enterocolitica выделены из устриц, салата с куриным мясом, тушеных грибов, картофеля, капусты, моркови, свеклы и фруктов.
Почва является резервуаром возбудителя, в котором он не только сохраняется, но и длительное время может размножаться. Китайские ученые в 1988г. показали, что жизнеспособная популяция кишечных иерсиний при достаточной аэрации почвы формируется к 10 дню. Отечественные исследователи установили, что почвы с высоким содержанием органических веществ, высокой влажностью и слабощелочной реакцией способствуют длительному сохранению данных бактерий. В качестве постоянно существующего резервуара и потенциального источника заражения человека и животных иерсиниями может рассматриваться вода в открытых и закрытых водоёмов и колодцев. Van Oye (1978) сообщил, что у 148 видов рыб выловленных в реке Вальдивии (Чили), были выделенны иерсинии. Таким образом, заражённая рыба способна инфицировать поедавших её людей и контаминировать воду. Установлено, что в речной воде она сохраняется более 1 года, в морской воде несколько недель, в стерильной воде 3 мес. По данным Кузнецова В.Г (1983) численность иерсиний в холодное время года в воде выше, чем в жаркое. Животноводческие фермы также являются резервуаром инфекции.
Y.enterocolitica, как и многие другие возбудители из обширной группы факультативных паразитов, могут существовать в организме теплокровных (паразитические свойства) и окружающей среде (сапрофитная фаза развития). Взаимодействие популяций иерсиний с грибами, простейшими, гельминтами и их яйцами, насекомыми, червями, личинками и нимфами членистоногих, а также наличие в почве и воде растительных субстратов способствуют размножению и длительному сохранению микроорганизмов. Признавая важное значение внешней среды как резервуара иерсиний в природе, следует указать на важную роль в распространении инфекции через млекопитающих и птиц.
По мнению Сомова Г.П. (1985), можно предполагать, что возбудители иерсиниозов сформировались во внешней среде. Встречаясь в процессе эволюции с первыми теплокровными, иерсиний в результате мутаций и отбора приобрели свойства, позволяющие им выжить в макроорганизме хозяина, что и привело к развитию патогенных свойств, то есть способности вызывать инфекционный процесс. При этом бактерии не утратили своих первоначальных сапрофитных свойств, управление которыми заложено в генетическом аппарате, обеспечивающем им возможность размножаться и длительное время существовать во внешней среде. Так как иерсинии являются факультативными паразитами, обладающими патогенными свойствами и сохраняющими способность существовать вне организма во внешней среде, то условия их жизнедеятельности обусловлены влиянием абиотических факторов, характерных для естественных мест обитания этих бактерий, таких, как температура окружающей среды, ультрафиолетовые лучи, повышенные концентрации натрия хлорида, рН среды и др. Наиболее важным из факторов внешней среды, определяющим жизнедеятельность микроорганизмов, является температура. К холоду иерсиний устойчивы, хорошо переносят температуру от -15 до –200С и в этих условиях могут длительно существовать. Для Y.enterocolitica важной особенностью является психротрофность - свойство довольно редкое для патогенных бактерий. То есть эти микроорганизмы при 4° С не только сохраняются, но и размножаются, поэтому иерсиниоз называют "болезнь из холодильника"
Механизм адаптации психротрофных микроорганизмов к биологически низким температурам до сих пор полностью не ясен. Вместе с тем установлено, что при понижении температуры у этих микроорганизмов наблюдается интенсивное поступление питательных веществ и расщепление их внутри клетки. Кроме того, способность микроорганизмов расти при низких температурах зависит от строения и липидного состава клеточной мембраны. В частности, в липидах клеточных мембран психротрофных бактерий содержится больше ненасыщенных кислот, чем в липидах мембран мезофилов, вследствие чего цитоплазматическая мембрана постоянно находится в жидкокристаллическом состоянии даже при низких температурах. Психрофильные свойства иерсиний способствуют возникновению и развитию эпидемического процесса, поскольку холод не только позволяет этим бактериям размножаться и накапливаться в объектах окружающей среды, но и является пусковым фактором генетико-биохимических механизмов, обеспечивающих регуляцию их вирулентности. В районах с умеренным климатом это инфекция выделяется в настоящее время значительно чаще, чем в сухих и жарких местах (инструкция Минздрава СССР. 1990). Широкое распространение иерсиниоза, многообразие клиники и трудоёмкость в постановке диагноза сделали актуальной проблему кишечного иерсиниоза в мировом масштабе. Изучение распространения иерсиниоза среди животных затруднено, так как эта болезнь в РФ не подлежит обязательному учёту как самостоятельная нозологическая единица.
КЛАССИФИКАЦИЯ, НОМЕНКЛАТУРА КУЛЬТУРАЛЬНО – МОРФОЛОГИЧЕСКИЕ СВОЙСТВА. Род Yersinia в соответствии с систематикой по Bergey’s (1994) входит в семейство Enterobacteriaceae, 5-ую секцию, 1-ый отдел Gracilicutes, царство Procaryotae. Ранее возбудители чумы и псевдотуберкулеза относили к роду Pasteurella и именовали Pasteurella pestis и Pasteurella psevdotuberculosis. J. van Loghem (1946) предложил выделить вышеуказанных бактерий в новый род - Yersinia, назвав его так в честь французского исследователя Иерсена (А. Yersin), впервые выделившего возбудителя чумы. В 1954 г. Н. Моllaret, E. Thal предложили включить род Yersinia в семейство Enterobacteriaceae. В дальнейшем в род Yersinia был включен возбудитель иерсиниоза - Yersinia enterocolitica (Pasteurella Х). В настоящее время род Yersinia состоит из 11 видов - Y.pestis, Y. psevdotuberculosis, Y. enterocolitica, Y. intermedia, Y. frederiksenia, Y. kristensenia, Y. aldovae, Y. bercovieri, Y. mollaretii, Y. rohdei, Y. ruckeri. Yersinia enterocolitica представляет собой палочки с закругленными концами от 0,8 до 1,2 мкм длиной и от 0,5 до 0,8 мкм шириной. Размеры палочек колеблются в зависимости от возраста культуры, температуры и среды выращивания. В условиях выращивания при 37С это чаще всего небольшие, одинаковые по размерам палочки (кокковидные), при 22-25С - обычные палочки средних размеров. В мазках из старых культур палочки различны по величине и форме. Yersinia enterocolitica окрашиваются всеми анилиновыми красителями (Ющенко Г.В., 1984). По Грамму – грамотрицательная, выращенная на жидкой среде окрашивается биполярно. Минимальная температура роста –1,3 0С, максимальная +44 0С (Reed, 1994). При температуре 18-20 0С бактерии подвижны. При более высокой температуре культивирования (37 0С) штаммы иерсиний неподвижны. Подвижность обеспечивается жгутиками, их число варьирует. Иерсинии - факультативные аэробы. Оптимальной для роста является рН среды 7,2-7,4, но возбудитель может расти при рН от 3.0 до 9.6 (Little,1992). Наиболее благоприятна для роста иерсиний температура 22-280С. Бактерии растут и при температуре 37-400С, но при температуре выше 440С рост прекращается. При пониженной температуре 4-14 0С возбудители растут, но темп роста по мере снижения температуры угнетается.
Иерсинии неприхотливы и нетребовательны к питательным веществам, растут на обычных питательных средах (МПА, агары Хоттингера и Мартена), средах - Эндо, ЭМС, Плоскирева, Серова, агаре Мак-Конни, SS-агаре. Хорошо растут иерсинии на шоколадном и кровяном агарах, в МПБ, 1% пептонной воде (Ющенко Г.В.,1984). Штаммы Y.enterocolitica вырабатывают фермент пенициллиназу, поэтому растут на средах с пенициллином, что является их отличительным признаком от Y. psevdotuberculosis (М.В. Дулатова, 1989). Через сутки инкубации, на агаре Эндо, при температуре 22-28 0С, вырастают мелкие (росинки), круглые, выпуклые, бесцветные, блестящие колонии. Лучшие результаты достигаются через 36-48ч. инкубации посева. На вторые сутки колонии увеличиваются в размерах и могут приобретать розоватый оттенок, иногда с красной точкой в центре. На слабощелочных средах (МПА, агары Хоттингера, Мартена) через 18-24ч. вырастают колонии размером от 0,1-0,2 до 0,5 мм, круглые, сферически выпуклые, с ровным или волнистым краем, бесцветные, блестящие, полупрозрачные мягкой консистенции с голубоватым оттенком в проходящем свете. Через 36-48ч. колонии теряют прозрачность и голубоватый оттенок.B.Nilehn (1967) выделила 5 биоваров, что было подтверждено методом нумерической таксономии. Подавляющее большинство штаммов Y. enterocolitica, имеющих клиническое значение, относятся к биоварам 2,3 и 4. В последнее время увеличивается число выделяемых от больных людей штаммов Y. enterocolitica биовара 1. Сообщения о выделении от человека Y. Enterocolitica относящихся к 5-му биовару, пока единичны.
УСТОЙЧИВОСТЬ. Иерсинии по своей устойчивости к физическим и химическим агентам во многом сходны с представителями семейства кишечных. Растворы дезинфицирующих веществ (хлорамин, карболовая кислота и алкоголь) в обычных прописях убивают микробов в течение нескольких минут. К холоду иерсинии устойчивы: хорошо переносят температуру от -200С и в этих условиях могут длительно существовать, при температуре от +4 до 140С не только сохраняются, но размножаются. В тушках цыплят, хранившихся при –18°С в течение 3-х месяцев, количество бактерий снижалось очень незначительно. Данный микроорганизм может размножаться в пищевых продуктах при температуре холодильника (+4°С), что представляет опасность для здоровья человека. В эксперименте установлено, что в свинине, хранившейся в течение 2‑х недель при температуре холодильника, количество бактерий в 1 грамме продукта увеличилось в три раза, по сравнению с первоначальными показателями. Ленченко Е.М, совместно с Штукаревой М.Ю были проведены опыты по изучению сроков выживаемости иерсиний в мясе и мясопродуктах. Установлено, что в говядине и свинине при 4°С иерсиний выживали 35 суток (срок наблюдения); при - 18°С сохранялись 75 суток. К воздействию высокой температуры микроорганизмы Y.enterocofitica менее устойчивы по сравнению с E.coli. При нагревании до 60-80° С бактерии погибают через 15-30 мин, при кипячении при 100°С - в течение нескольких секунд. При температурной обработке продуктов (варка мяса, пастеризация молока), применяемых в промышленности, Y.enterocolilica быстро погибают. По сравнению с кишечной палочкой Y.enterocolitica менее устойчива к воздействию УФ-облучения, гамма-лучей. На прямом солнечном свету иерсинии в течение 30 мин инактивируются, при рассеянном отмирают через 6-8 ч. Быстро они погибают при высыхании.
Величина кислотности продукта влияет на выживаемость бактерий. Ее рост прекращается при показателе рН 5,0. Однако сообщают, что этот микроорганизм выделили из острокислого соуса для бифштексов.
В отношении влияния на жизнеспособность иерсиний поваренной соли данные различны. Есть публикации, что концентрация в 4% угнетает рост одного из штаммов, однако бактерии другого штамма в 80% случаев выживали после контакта с поваренной солью данной концентрации. Считается, что бактерии растут при содержании NaCl - 7% (Reed, 1994). При изучении влияния различных концентраций натрия хлорида на рост и изменчивость иерсинии Ленченко Е.М. и др (1998) установили, что, на агаре с содержанием NaCI 30 г/л Y.enterocolitica способны расти, тогда как Y.pseudotuberculosis не вырастают. Таким образом, Y. enterocolitica обладает широким спектром ферментов, обеспечивающих им питательные и энергетические потребности и возможность существования в различных условиях, в том числе и в окружающей среде. (Г.В.Ющенко, 1984).
СЕРОЛОГИЧЕСКИЕ СВОЙСТВА. Учитывая, что для диагностики кишечного иерсиниоза и идентификации микроорганизмов Y. enterocolitica используют антигенную структуру бактерий остановимся на серологических свойствах данного микроорганизма. Бактерии имеют сложное антигенное строение. В соответствии с морфологией микроорганизма состоящего из жгутика фимбрий и собственно микробного тела, Y.enterocolitica имеет, жгутиковый H-антиген, соматический O-антиген и общий со всеми Enterobocteriacea K-антиген (М.В.Дулатова, 1989). По мнению Путилиной Н.Г. (1982г.) антигенный состав Y.enterocolitica представлен 16 компонентами: 12 термолабильных и 4 термостабильных. Соматический антиген сложен по своему составу, в его основе лежит липополисахарид, который является эндотоксином клетки. Различия в строении О-антигенов позволили выделить более 50 сероваров Y.enterocolitica. Наибольшее значение в патологии человека имеют серовары 03; 05; 07,8; 08; 09. При разрушении бактериальных клеток формируется эндотоксин. Для большинства штаммов Y.enterocolitica различных сероваров характерны адгезия, колонизация на поверхности кишечного эпителия и энтеротоксигенность с продукцией большого количества термостабильного энтеротоксина. Иерсинии этого вида обладают также способностью к инвазии и внутриклеточному размножению. Есть сообщение Glosnicka R. (1980), что в антигенную структуру Y. enterocolitica входят компоненты капсульного антигена, сходного с чумным капсулярным антигеном F1. Отмечено, что штаммы Y. enterocolitica, патогенные для человека и обладающие вирулентностью, способны продуцировать антигены вирулентности V и W, известные у Y. pestis и Y.psevdotuberculosis. M.Doyle c соавторами (1982) отметили, что вирулентные штаммы синтезируют присущие только им антигены. Полученная против этих антигенов сыворотка реагирует только с вирулентными штаммами. Иерсинии Y.enterocolitica сероваров 03; 09; 05,27; 08 выделяются от животных (свиней, коров, грызунов диких и синатропных), из овощей, мясных и молочных продуктов, воды небольших водоемов. Из окружающей среды (почва, грызуны, вода, рыба и т.д.) на разных территориях выделяются иерсинии всех известных сероваров, в том числе и не отмеченные в патологии человека. Антигенные связи у иерсиний серовара 09 с бруцеллами всех видов (Brucella abortus bovis, Br.suis и Br.melitensis) очень значительны. Антигенное родство выявляется в РА, РСК и других серологических реакциях. Перекрестные антитела находят в сыворотках больных бруцеллезом и иерсиниозом, что затрудняет диагностику этих заболеваний. Часто выявляемый в сыворотках больных бруцеллезом людей и животных титр антител, по указанным реакциям, к штаммам иерсиний выше, чем бруцеллам. Антигенное родство этих возбудителей связано с общими антигенными компонентами липополисахаридного комплекса, расположенного в клеточной стенке.
БОЛЕЗНЕННОСТЬ (ПАТОГЕННОСТЬ). Бактерии Yersinia enterocolitica в процессе своей жизнедеятельности и размножения выделяет токсическое вещество двух типов. Первый – термостабильный энтеротоксин, стойкий к воздействию температуры в 100-120°С в течение 20 минут. Причем, что важно, образуемый при комнатной температуре (например в пищевых продуктах) +25°С более ядовит, чем токсин, образуемый при температуре тела +37°С. Но не все штаммы этой бактерии способны продуцировать подобный токсин. Чаще всего это штаммы, выделенные из продуктов питания, особенно из молока. У штаммов, выделенных из объектов окружающей среды, подобное свойство встречается реже. В настоящее время установлено, что большинство штаммов, выделенных от больных людей, обладают энтеротоксином. Второй тип токсина - термолабильный, разрушающийся при 100°С в течение нескольких секунд. Одним из факторов, повышающих патогенность иерсиний, является их способность внедряться в клетки ткани человека или животного и размножаться в ней с последующим ее разрушением, то есть они обладают инвазией. Это свойство характерно для тех штаммов, которые также выделены от больных. Еще одним свойством, повышающим болезнетворное действие иерсиний, является их способность проявлять устойчивость к действию нормальной человеческой сыворотки крови. Таким образом, говоря о болезнетворности данных бактерий, необходимо учитывать особенности их размножения и жизнедеятельности. Проникнув в организм через ротовую полость, иерсинии попадают в желудок, где частично погибают под действием кислой среды. Бактерии, преодолевшие защитный барьер желудка, внедряются в слизистую оболочку подвздошной кишки и ее лимфоидные образования. В месте входных ворот инфекции развивается различной выраженности воспалительный процесс. По лимфатическим сосудам иерсинии проникают в мезентеральные лимфатические узлы и вызывают мезаденит. В патологический процесс может быть вовлечена слепая кишка. На фоне инфекционно-воспалительных изменений развиваются токсический и аллергический процессы, связанные с токсинемией. На этом этапе инфекционный процесс, приобретая черты локализованной формы, может завершиться. В случае прорыва лимфатического барьера кишечника возникает бактериемия, обуславливающая развитие генерализованных форм заболевания. Наблюдается токсико-аллергическое поражение многих органов и систем, в первую очередь печени и селезенки, возможно развитие полилимфаденита, полиартрита, остита, миозита, нефрита, уретрита, менингита и др. Иммунопатологические реакции у части больных иерсиниозом могут служить толчком к развитию системных заболеваний (тип коллагенозов). Заключительное звено патогенеза иерсиниоза - освобождение организма от возбудителя, ведущее к выздоровлению. Развивающийся иммунитет при иерсиниозе не является прочным, с чем связано нередкое развитие обострений и рецидивов болезни (Е.П.Шувалова 1995).
ЛАБОРАТОРНАЯ ДИАГНОСТИКА. Диагностика кишечного иерсиниоза затруднена в связи с многообразием клинических проявлений, а также появлением секундарной инфекции и возможна только лабораторными методами. Существуют бактериологические, наиболее достоверные, и иммуннобиологические методы. Они позволяют проводить как эпизоотологическую разведку, индикацию возбудителя во внешней среде, так и диагностировать заболевание у людей и животных.
Бактериологическое исследование трудоемко и основано на использовании различных добавок к средам обогащения и подращивания на холоде, далее пересев на дифференциально диагностические среды с последующим термостатированием при 280С и отбор подозрительных колоний через 42 часа. Диаметр и вид выросших иерсиний позволяют дифференцировать их от другой кишечной микрофлоры с дальнейшим использованием дифференциальных тестов. Для идентификации иерсиний используются также и другие методы: РА, РНГА, РТГА, РИФА, ИФА, ПЦР.
ВНЕШНИЕ ПРИЗНАКИ БОЛЕЗНИ (КЛИНИКА). Внешне заболевание кишечным иерсиниозом проявляется у людей очень разнообразно. При этом доминирует энтероколит, сопровождающийся кашице водянистыми испражнениями, частично с примесью слизи и крови. Часто встречается форма течения заболевания как псевдоаппендицит (мезентеральный лимфаденит). В 6% случаев исследования человеческого аппендикса обнаруживается Y.enterocolitica. Значительней, чем острая фаза, при этой инфекции является фаза осложнений, при которой могут возникать артриты, заболевания кожи и даже смертельно протекающий сепсис.(Х.Наттерман, 1987). Помимо того, что возможно его течение в виде гастроэнтероколита (воспалительный процесс в желудочно-кишечном тракте) у взрослых оно сопровождается тошнотой, поносами, болями в животе, у детей протекает в виде энтеритов. Часто появляются диффузные боли в области кишечника, локализующиеся в области пупка или правой подвздошной области. Температурная реакция различна, ее может не быть или она достигает показателя в 38-39°С. Течение болезни может проходить в форме энтероколита (одновременное воспаление слизистых оболочек толстых и тонких кишок). В этом случае развиваются более тяжелые поражения, имеющие уже другие клинические проявления. Одновременно с признаками энтероколита, или через два дня у больных появляются сильные боли в правой плеоцекальной области, повышается температура до 38-39°С. В этом случае подозревают аппендицит, и больного необходимо отправить в больницу. Довольно часто кишечный иерсиниоз протекает как генерализованная инфекция, развивающаяся после возникновения поноса и болей в животе. Повышается температура до 39°С, появляется головная боль, боль в мышцах и суставах, резкая слабость. Клинические проявления со стороны кишечного тракта могут и отсутствовать. Позднее появляется сыпь сходная с краснухой. Характерной является сыпь на ладонях с последующим шелушением. Данная инфекция может протекать как сепсис, часто встречающийся у маленьких детей и сопровождающаяся высокой температурой, лихорадкой, желтушным окрашиванием кожи, сыпью. Почти при всех формах течения иерсиниоза поражается, в той или иной степени, печень. Зачастую иерсиниозный гепатит по клинике напоминает вирусный гепатит. Возможным осложнением иерсиниоза является поражение суставов - артриты. Как правило, воспалительным явлениям в суставах предшествуют: лихорадка, понос, боли в животе. Частым осложнением при данном заболевании является узловая эритема - появление крупных болезненных узелков. Регистрируются при иерсиниозе и воспалительные явления в глазах. Возможно поражение почек, легких и плевры.
У сельскохозяйственных животных клиническое проявление данной инфекции изучено недостаточно, хотя установлено, что до 50% здоровых свиней являются носителями этой бактерии. Внешние признаки болезни у них проявляются в повышении температуры, легким поносом и через 11-15 дней вторичным повышением температуры с последующим ухудшением общего состояния организма. У коров течение болезни характеризуется поносом, возможен аборт и мастит. Причем, наряду с безсистемным носительством описаны и клинически выраженные формы заболевания, сопровождающиеся поносом, поражением суставов и зачастую приводящие к гибели животного. Животные как и люди могут заражаться иерсиниозом через корм, воду, подстилку, которая загрязнена фекальными массами грызунов, являющимися носителями данных микроорганизмов.
МЕТОДЫ ПРОФИЛАКТИКИ кишечного иерсиниоза у людей аналогичны описанным в главе о эширихиозах.
САНИТАРНАЯ ОЦЕНКА МЯСА И МЯСОПРОДУКТОВ в нашей стране не разработана, но вполне допустимо подходить к этому вопросу (учитывая слабую устойчивость бактерий к температурному воздействию и отсутствию спор, которые очень устойчивы к температуре) так же, как и при сальмонеллезной инфекции. Рубить мясо на куски по 2 кг и варить их примерно 2,5 часа, а внутренние органы уничтожить.
Камилобактериоз
Кампилобактериоз - зооантропонозная бактериальная острая инфекционная болезнь с фекально-оральным механизмом передачи возбудителя. Характеризуется сочетанием симптомов интоксикации и поражения желудочно-кишечного тракта.
ИСТОРИЧЕСКАЯ СПРАВКА. Имеются данные, что кампилобактеры выделяли от больных людей и животных еще в конце XIX - начале XX в. В 1884 г. Т.Эшерих сообщил, что он выделил спиралевидные бактерии из слизистых испражнений больных азиатской холерой во время эпидемии в Неаполе. В 1886 г. аналогичные спириллы он нашел в кишечнике больных диареей котят. Эти спириллы обнаруживали в их испражнениях, слизи толстой кишки, выводящих протоках желез; отмечено также проникновение их в клетки эпителия. Длина возбудителей 3-18 мкм, толщина в окрашенном препарата 0,5-0,8 мкм. Чаще встречались спириллы с 3-4 изгибами. О наличии у больных с клинической картиной холеры спиралевидных бактерий, не поддававшихся культивированию, сообщали и другие исследователи. Первое упоминание о вибрионоподобных возбудителях, обнаруженных у овец при эпизоотических выкидышах, которые, судя по описанию, являлись Campylobacter fetus, опубликовано в 1909 г. McFadyen, Stockman. Заболевания, вызываемые данными микроорганизмами, получившие общее название "вибриозы", описывались в первой половине XX в., главным образом у животных. Лишь в 1947 г. K. Vinzent удалось выделить бактерии данного рода из крови, длительно лихорадившей 39-летней женщины. Это было первое упоминание о потенциальной важности этого микроорганизма в заболевании людей. Важный шаг на современном этапе изучения кампилобактериоза сделала в своих трудах Е. King, которая отметила взаимосвязь кампилобактерий с энтеритами у людей. Основными источниками возбудителя инфекции являются животные и птицы, преимущественно сельскохозяйственные. Крайне редко - больные люди. Основной путь передачи возбудителей кампилобактериоза - пищевой. Заражение чаще всего происходит при употреблении в пищу инфицированного мяса: говядины, свинины, мяса птицы. Существенную роль как фактор передачи возбудителей играет молоко. С употреблением молока связано большинство пищевых вспышек кампилобактериоза в США, причем на долю этих вспышек пришлось около 80% заболеваний. Несомненное значение имеет и водный путь передачи инфекции, с которым связывают очень крупные вспышки кампилобактерного энтерита в США, Великобритании, Швеции, Канаде. Кампилобактериоз составляет от 5 до 14% всех регистрируемых случаев заболеваний с признаком расстройства работы желудочно-кишечного тракта. В Англии в 1979 году зарегистрировано 8577 случаев кампилобактериоза в то время как случаев сальмонеллеза - 12 251.
Первые работы отечественных исследователей по изучению кампилобактериоза в нашей стране относятся к 80-тым годам. В этих работах были показаны актуальность изучения кампилобактериоза и достаточно высокий уровень заболеваемости людей (до 8,7% и более). В России кампилобактериоз регистрируется во многих городах и регионах, составляя 6,5-12,2% от общего числа ОКЗ. Отмечена летне-осенняя сезонность кампилобактериоза
ЭТИОЛОГИЯ. Первоначально кампилобактеры были отнесены к семейству Vibrionaceae, но по морфологическим характеристикам они значительно отличались. В 1963 г. был выделен отдельный род Campylobacter семейства Spirillaceae. Он включил в себя С. fetus и родственные ему микроорганизмы. Хотя род Campylobacter включает в себя более 10 видов, обнаруженных как у животных, так и у человека, но наибольшее значение как возбудители острых диарей у человека имеют С. jejuni, С. coli, С. laridis, С. fetus. В подавляющем большинстве случаев кампилобактериоз, как у детей, так и у взрослых, обусловлен С. jejuni.
МОРФОЛОГИЯ КУЛЬТУРАЛЬНЫЕ СВОЙСТВА. Кампилобактеры - тонкие подвижные грамотрицательные, неспорообразующие изогнутые палочки, имеющие форму запятой или S-образные, длиной 0,5-0,8 мкм, толщиной 0,2-0,5 мкм. Характерные винтообразные движения обусловлены наличием одного или двух полярно расположенных жгутиков длиной до 15 мкм. Полиморфизм обычен среди этих клеток. Измененные формы представлены изогнутыми или спиралевидными микроорганизмами, которые вариабельны по длине и толщине. В старых культурах преобладают клетки сферической и кокковидной формы. Кампилобактеры - микроаэрофилы. Штаммы С. jejuni лучше растут при температурном оптимуме 42 оС и следующем газовом составе: O2 - 5%, СОз- 10%, N2- 85%, для создания микроаэрофильных условий в анаэростате замещают 75% воздуха смесью из 90% азота и 10% С02. Оптимальную газовую среду получают с помощью коммерческих газогенераторных пакетов (например, Campy Pak 11); удовлетворительный рост можно получить в анаэростате со свечой. В полевых условиях используют стерильные пластиковые пакеты, в которые вносят чашку, засеянную исследуемым материалом, и чашку, засеянную чистой культурой любого факультативного анаэроба (например, эшерихиями); последние будут поглощать 02 и выделять С02 в процессе роста.
С. fetus могут расти и в анаэробных условиях. Оптимум рН 7,0-7,2. Кампилобактеры плохо переносят транспортировку поэтому их следует помещать в консервант, например, тиогликолевый бульон (рН 8,5) или щелочную пептонную воду, содержащую 0,05% тиогликолата Na и 0,025% цистеина, и хранить при температуре 4 °С. Для получения более чистых культур используют посев на селективные среды, например среды Скирроу, Бутцлера или Престона, среда Сатру-ВАР. На твёрдых средах С. jejuni образуют колонии двух типов — плоские влажные слизистые сероватые «расползающиеся» колонии с неровными краями (могут быть очень мелкими и напоминать капли конденсата) либо мелкие дискретные блестящие выпуклые колонии размером до 1-2 мм. На полужидких и жидких средах образуют поверхностную голубовато-серую плёнку.
Кампилобактеры инертны к углеводам; кислых или нейтральных конечных продуктов не образуют; большинство видов редуцирует нитраты; образуют H„S; молоко не свёртывают; не проявляют гемолитическую активность. Для роста не требуют сыворотки, энергию получают от аминокислот или промежуточных продуктов цикла Крёбса. Реакции Фогеса - Проскауэра и с метиловым красным отрицательные; желатину и мочевину не гидролизуют; оксидаза-положительны; липазная активность отсутствует, пигментов не образуют.
УСТОЙЧИВОСТЬ. Кампилобактеры довольно устойчивы во внешней среде. При температуре 4°С и ниже они длительно сохраняют жизнеспособность в речной воде, мясе, молоке, фекалиях. В связи с тем, что это сравнительно недавно выявленная токсикоинфекция, устойчивость возбудителя при хранении пищевых продуктов малоизучена. Установлено, что при нагревании раствора, содержащего данную бактериальную культуру, до 60°С кампилобактерии гибнут через минуту, при пастеризации в 71-77°С через 15 секунд. В воде, хранящейся при температуре б°С, они сохраняют жизнеспособность до 1 месяца. При замораживании молока до -20°С 5-8 месяцев. В замороженном мясе сохраняются не менее 60 дней. В стерильном молоке бактерии жизнеспособны при +4С в течение 22 дней. На тушках птицы, в фарше количество бактерий не уменьшается неделями. При хранении продуктов в холодильнике, этот показатель сокращается в два раза. Поваренная соль в концентрации 1,0-2,5% при температуре -25°С достаточно быстро приводит к отмиранию бактерий, но при температуре -4°С этот процесс идет медленнее. При низких показателях кислотности (кисломолочные проекты) бактерии также быстро инактивируется
АНТИГЕННАЯ СТРУКТУРА - сложная, особенно состав термолабильных Аг. Важнейшие поверхностные Аг представлены ЛПС (0-Аг) и кислоторастворимыми низкомолекулярными веществами, белковыми фракциями, играющими ведущую роль в серотипировании. 0-Аг определяют иммунологическую специфичность кампилобактеров, выявляемую в РНГА; а антигенные различия между разными сероварами обусловлены варьированием углеводного состава внутреннего ЛПС. Недавно открыт белковый жгутиковый Аг, общий для все сероваров.
ПАТОГЕНЕЗ (БОЛЕЗНЕТВОРНОСТЬ) заболеваний обусловлен высокой адгезивной и инвазивной активностью кампилобактеров. Последние размножаются в присутствии жёлчи и быстро колонизируют верхние отделы тонкой кишки. Прикрепление бактерий к слизистой оболочке во многом обеспечивают поверхностные специфические адгезины; большое значение имеет способность кампилобактеров проникать через слизь и перемещаться вдоль эпителия (жизнеспособность в слизи сохраняют более 30 мин), т.е. жгутики — также важный фактор адгезии. Кампилобактеры легко проникают через мембрану эпителиальных клеток и межклеточные ходы, например, после внесения бактерий в культуру клеток HeLa и часовой инкубации практически во всех клетках обнаруживают бактерии. Клинический признак, подтверждающий их инвазивностъ — наличие крови и слизи в испражнениях больных. Колонизация кампилобактерами тонкой кишки приводит к развитию воспалительных изменений и отёка слизистой оболочки. После первого этапа прикрепления возбудителей к поверхности клеток происходит перфорация клеточной мембраны, и уже через 10 мин эти бактерии попадают внутрь клеток. Кампилобактеры выживают в клетках периферической крови до 6-7 дней, что способствует повышению их вирулентности. Интерстициальньй мукозный гель, выделяемый специальными клетками крипт и секреторные IgA, являются защитным барьером, препятствующим адгезии кампилобактеров. В связи с наличием такого местного иммунитета прикрепление кампилобактеров даже при высокой патогенности штамма может не состояться и заболевание не разовьется, т.е. для каждого человека существует своя инфицирующая доза. Группу повышенного риска составляют дети (особенно до 2 лет), пожилые, страдающие сопутствующими заболеваниями, пациенты с иммунодефицитами, либо получающие стероиды и цитостатики.
Определённый вклад в патогенез поражений вносят термолабильный и термостабильный энтеротоксины. По механизму действия термолабильный энтеротоксин напоминает термолабильные диареегенные токсины эшерихий и холерного вибриона (повышает уровень внутриклеточного цАМФ). Термостабильный энтеротоксин (эндотоксин) высвобождается после гибели кампилобактеров, проявляет все свойства эндотоксинов грамотрицательных бактерий. Вызывает гибель мышей после введения живых или убитых микроорганизмов.
Как и при каждом остром инфекционном диарейном заболевании, при кампилобактериозе отмечаются определенные изменения в стенке кишечника. Изменения слизистой оболочки толстой кишки носят в основном катарально-геморрагический характер и проявляются отеком слизистой и ее гиперемией, а также наличием геморрагий. Все эти явления наблюдаются равномерно по всей поверхности слизистой оболочки толстой кишки. Для кампилобактериоза нехарактерно эрозивное поражение слизистой оболочки толстой кишки.
КЛИНИЧЕСКАЯ КАРТИНА. Инкубационный период колеблется от 1 ч до 11 дней, составляя в среднем 2-5 дней. Примерно у 30-50% больных развитию характерных клинических проявлений болезни предшествует лихорадочный продромальный период с неспецифическими гриппоподобными симптомами, который может продолжаться 2~3 сут. Типичными признаками этого периода являются общая слабость, артралгии, головная боль, ознобы. Температура тела чаще всего остается в пределах 38-40°С. Болезнь может начинаться остро с одновременного развития всех симптомов. Клиническая картина кампилобактериоза полисимптоматична, что обусловлено разнообразием форм: от бессимптомного бактериовыделения до генерализованной инфекции. Наиболее часто кампилобактериоз - это острое диарейное заболевание, которое может иметь форму острого гастрита, гастроэнтерита, гастроэнтероколита, энтероколита и колита. Последние две формы отмечаются у подавляющего большинства больных в Европе, Северной Америке и Японии. Острый кампилобактерный гастрит и гастроэнтерит практически неотличимы от пищевой токсикоинфекции другой этиологии и для нашей страны не столь актуальны.
В России кампилобактериоз чаще всего протекает по типу острой дизентерии. Колитический вариант кампилобактериоза включает в себя сочетание симптомов интоксикации, болевой синдром и диарею. Боль в животе может появиться как в первые сутки заболевания, так и спустя 1-2 дня, а иногда предшествует диарее. Длительность болевого синдрома составляет от нескольких часов до 7-10 дней. Локализация бывает различной. Чаще всего боль отмечается в левой подвздошной области, однако она может быть разлитой по всему животу. Интенсивность боли тоже различная, что может быть связано с особенностью характера пациента, но иногда боль настолько сильна, что симулирует картину острого живота, в частности острого аппендицита. Диарея развивается одновременно с болевым синдромом или несколько позднее. Стул быстро становится жидким, нередко кал окрашен желчью. На 2-3-й день испражнения становятся скудными, со слизью, появляются прожилки крови. Стул в виде "ректального плевка" встречается примерно у половины больных с колитическими вариантом кампилобактериоза. Частота стула зависит от тяжести течения болезни и может быть от нескольких раз до 10 и более в сутки. Общая длительность дисфункции кишечника варьирует от 3-4 до 10-14 дней. При кампилобактерном колите редки такие симптомы, как тенезмы и ложные позывы.
Как редкие симптомы при кампилобактериозе, описаны полиморфная сыпь, появляющаяся в различные сроки заболевания, а также артриты, конъюнктивит. Чаще всего кампилобактерный колит ограничен 1-2 нед, хотя есть данные о более длительном течении, описаны рецидивы. Следует отметить, что есть наблюдения, доказывающие различия в клинических проявлениях в зависимости от вида кампилобактера. Сравнивали клинические проявления колитов обусловленных С. jejuni и С. coli. Наличие слизи и крови в фекалиях достоверно чаще встречалось при заболеваниях, вызванных С. jejuni. Энтероколиты, обусловленные С. jejuni, протекали более тяжело, чем вызванные С. coli. По другим клиническим проявлениям различий не было. Окончательный диагноз кампилобактериоза возможен только после выделения возбудителя.
ПРОФИЛАКТИЧЕСКИЕ МЕРОПРИЯТИЯ. В виду того, что заболевание у людей выявлено сравнительно недавно, специальных мер его профилактики еще не разработано. Установлено, что значительное количество больных людей заразилось от кур (или мяса кур), у которых наблюдается очень большой процент носительства этого микроорганизма. Поэтому и профилактика болезни строится на соблюдении режима личной гигиены после работы в курятнике. Что касается сельскохозяйственных животных, то если они абортировали или долго бесплодны, перегуляли, необходимо это принимать во внимание и после работы на ферме, скотном дворе обязательно соблюдать и меры личной гигиены. Необходимо помнить, что обычные дезинфектанты быстро убивают этот микроорганизм. Если убили больное животное или подозреваете, что оно является скрытым носителем данной бактерии, то необходимо отослать образцы мяса, внутренних органов в лабораторию для исследования на наличие данного возбудителя.
ВЕТЕРИНАРНО-САНИТАРНАЯ ОЦЕНКА МЯСА. При получении результатов, подтверждающих наличие кампилобактерий в мясе, его используют в пищу после проварки. Хотя именно для данной болезни ветеринарно-санитарная оценка мяса и внутренних органов не определена, но медицинские врачи рекомендуют поступать также как и при сальмонеллезе.
Ботулизм
Ботулизм - тяжелое заболевание, связанное с употреблением в пищу продуктов, содержащих или бактерии Cl.botulinum или ботулинический токсин, продуцируемый этими бактериями.
ИСТОРИЧЕСКАЯ СПРАВКА. Данная болезнь прослеживается на глубину примерно Х веков. Ее возникновение обусловлено стремлением человека к созданию запасов пищевых продуктов и разработке новых методов консервирования. В медицинской литературе описание симптомов ботулизма впервые появилось в ХVIII в. Особое внимание привлекла вспышка заболевания, вызванного употреблением в пищу колбасы в 1792 году. В начале ХIХ в. 234 человека отравились при поедании кровяной и ливерной колбас. Позднее установили, что сходное по внешним признакам заболевание возможно при употреблении в пищу копченой ветчины, соленой рыбы, заготовленных впрок овощей и фруктов.
Ван-Эрменган в 1879 году доказал, что данная болезнь связана с токсином, выделяемым бактериями особой группы. Из остатков ветчины, а также из селезенки и толстого отдела кишечника человека, съевшего этот продукт, он выделил микроорганизм, который при росте на жидкой питательной среде продуцировал токсин и при заражении этой средой морских свинок, кроликов, кошек, обезьян возникало такое же заболевание с последующей смертью подопытного животного. По предложению ученого заболевание названо ботулизмом, а микроорганизм получил название Cl. botulinum. Сейчас выделено 7 типов этого микроорганизма, вызывающих разную степень болезнетворности. Смертельный исход при этой болезни различен и зависит от многих причин. В Советском Союзе данный показатель за период с 1958 по 1968 гг. составил: от овощных и фруктовых консервов – 31%, от грибов – 16,6%, от мясных продуктов – 29,9%, от рыбы – 16,5%, от ветчины – 6% (показатель от числа заболевших ботулизмом). Ботулизм не контагиозен. Заболевание наступает лишь при употреблении пищи, содержащей возбудителя и его токсины. Поскольку C.botulinum — строгий анаэроб, наилучшие условия для его размножения и продукции токсина создаются в консервированных продуктах, куда споры могут попасть с частичками почвы. Они могут выдерживать термическую обработку консервов, а затем прорастать и продуцировать токсин, чему способствует длительное хранение консервов. Известны групповые вспышки ботулизма, например вспышка в 1933 г. в городе Днепропетровске, когда в результате отравления кабачковой икрой заболело 230 человек, 26 из них умерли. В каждой стране источником отравления служат консервы, пользующиеся наибольшим спросом: в Германии, Франции и других европейских странах — мясные консервы, колбаса, ветчина и т. п.; в США — растительные консервы; в России — рыба и рыбные консервы.
МОРФОЛОГИЯ И КУЛЬТУРАЛЬНЫЕ СВОЙСТВА Вегетативные клетки — палочки с закругленными концами размером 4-8´0,6-0,8 мкм, подвижны (перитрихи). При неблагоприятных условиях образуют эндоспоры, расположенные терминально и субтерминально. Строгие анаэробы; молодые культуры окрашиваются грамположительно, 4-5-суточные — грамотрицательно. Оптимум рН для роста 7,3-7,6, для прорастания спор 6,0-7,2.
Морфология колоний. На кровяном агаре с глюкозой образуют очень мелкие сероватые или мутные желтоватые колонии линзообразной формы (на различных средах у одного и того же штамма могут варьировать). Вокруг колонии образуются зоны гемолиза различной ширины. На печеночном агаре образуют полиморфные звездчатые колонии, на желатине — сероватые, окруженные зоной разжиженного желатина. На столбике агара можно обнаружить диссоциаты, R-формы имеют форму чечевичных зерен, S-формы — пушинок. Хорошо растут на жидких средах (обычно на бульоне Тарроцци, бульонах из гидролизатов казеина, мяса или рыбы) при условии предварительного удаления О2 из среды кипячением в течение 15-20 мин с быстрым охлаждением. Вызывают помутнение среды и газообразование, иногда имеется запах прогорклого масла, но этот признак непостоянен.
Биохимические свойства. Все типы Clositridium botulinum образуют желатиназу, лецитиназу и Н2S, проявляют широкий спектр сахаролитической активности (бактерии типов А, В, Е и F ферментируют глюкозу, левулезу, фруктозу, мальтозу и сахарозу типов С и D — глюкозу и мальтозу, тип G инертен к углеводам). Clostridium botulinum типов А и В обладают выраженными протеолитическими свойствами, разлагают свернувшийся яичный белок и гидролизуют желатин По биохимическим свойствам выделяют 4 группы бактерий Cl.botulinum.
Бактерии I группы проявляют выраженные протеолитические свойства, гидролизуют желатин и эскулин, ферментируют глюкозу и мальтозу, проявляют липазную активность на яичном агаре.
Бактерии II группы проявляют сахаролитическую активность, но лишены протеолитической.
Бактерии III группы проявляют липазную активность и гидролизуют желатин.
Бактерии IV группы гидролизуют желатин, но, в отличие от прочих возбудителей ботулизма, не проявляют сахаролитических свойств и липазной активности, что послужило основанием для предложения выделить их в отдельный вид —Clostridium argentiense.
АНТИГЕННЫЙ СОСТАВ. Серологическая идентификация Clostridium botulinum основана на выявлении токсинов, по их структуре бактерии разделяют на 8 сероваров — А, В С1 (a), С2 (b), D, Е, F и G. Антигенная структура бактерий остается малоизученной, показано наличие жгутиковых, группоспецифических (Н) и соматических, типоспецифических (О) Аг, не проявляющих токсических свойств. Оптимальная температура для токсинообразования вариабельна для бактерий типов А, В, С и D 35°С, для бактерий типов Е и F 28-30°СУСТОЙЧИВОСТЬ. Длительное сохранение C.botulinum в природе и в различных пищевых продуктах связано с их спорообразованием. Споры сохраняются в почве долгое время, а при благоприятных условиях в летнее время могут прорастать и размножаться. Они хорошо переносят низкие температуры (не погибают даже при —190 °С). В высушенном состоянии сохраняют жизнеспособность десятилетиями. Споры устойчивы и к нагреванию (особенно типа А). Споры типов А и В переносят кипячение в течение 5 ч, при 105 °С погибают через 1-2 ч, при 120 °С — через 20-30 мин. Встречаются штаммы, споры которых переносят температуру 120 °С в течение нескольких часов. Наименее устойчивы к температуре споровая форма бактерии типа Е. При 100°С она отмирает примерно через 2 минуты, при 80°С - через 40 минут. Но нельзя забывать тот факт, что теплопроводимость любого пищевого продукта иная по сравнению с водой. По данным литературы, термоустойчивость спор в консервах «Говядина тушеная» вдвое превышала термоустойчивость таких же споровых форм бактерий, но только в воде. Содержание жира повышает устойчивость спор к температуре. Неспоровая форма бактерий (вегетативная форма) под действием 80°С отмирает в течение 30 минут, а при кипячении - через 5 минут. Споры C.botulinum устойчивы ко многим бактерицидным веществам: 20%-ный раствор формалина убивает их через 24 ч; этиловый спирт — через 2 мес; 10%-ая НС1 убивает их лишь через 1 ч. При консервировании пищевых продуктов методом понижения показателей рН, то есть за счет использования кислой среды (маринады) возможна задержка или даже прекращение роста этих микроорганизмов. Но данный процесс зависит от состава пищи в консервах. Кроме того существует такая закономерность: чем более кислая среда, в которой находится продукт, тем слабее внешние признаки его порчи, если туда попала палочка Cl.botulinum. Установлено, что при показателях рН свыше 4,2 (это такие консервы как «Лечо», «Борщ без мяса», «Овощи в томатном соусе», «Капуста натуральная», «Перец, фаршированный овощами и рисом в томатном соусе» и некоторые другие) микроорганизмы не только сохраняются, но и выделяют токсин без внешних признаков порчи продукта (образование газа, помутнение жидкости). Поваренная соль является одним из немногих консервантов, влияющих на размножение и производство токсина у данного микроорганизма. Отсутствуют конкретные указания какой именно процент поваренной соли задерживает размножение этих бактерий или убивает. Все зависит от типа продукта, величины показателя рН (кислотности), температуры консервирования и хранения. Но на практике исходят из того, что образование токсинов может быть подавлено, при содержании в продукте 5-6% соли.
Токсины C.botulinum также весьма устойчивы к действию химических факторов. Они не разрушаются протеолитическими ферментами кишечного тракта. В кислой среде (рН 3,5-6,8) они более устойчивы, чем в нейтральной или щелочной среде, разрушаются от действия 2-3%-ной щелочи, но высокие концентрации NaCI в пищевых продуктах их не разрушают; в консервах токсины сохраняются длительное время. Токсины C.botulinum обладают и определенной термоустойчивостью: при 58 °С разрушаются через 3 часа; при 80 °С — через 30-60 мин; а при 100 °С — в течение 10-15 минут. В твердых субстратах данная температура разрушает его за 2 часа. В зерне токсин сохраняется в течение нескольких месяцев. Наиболее устойчивы токсины типа С, менее устойчивы токсины типов D и Е, а токсины типов А и В занимают промежуточное положение. Устойчивость токсинов к высокой температуре зависит от вида продукта, его рН и других условий. В частности, при наличии жиров, высокой концентрации сахарозы устойчивость токсинов к высокой температуре возрастает.
БОЛЕЗНЕТВОРНОСТЬ (ПАТОГЕННОСТЬ) Clostridium botulinum различна для различных видов млекопитающих; заболевания человека вызывают бактерии типов А, В, Е и F; Clostridium botulinum типов С и D вызывают заболевания животных и птиц (в редких случаях от больных животных выделяют бактерии типов А и В). Патогенность типа G для человека и животных не доказана.
Патогенность Cl.botulinum определяется токсином. Токсин, выделяемый палочкой Cl.botulinum, один из самых сильных ядов в мире. По данным Л. Месробяну (1963г.) достаточно 10 мг токсина типа «Д», чтобы убить все население Земного шара. Болезнетворная активность токсина типа «А» такова, что 1 г его достаточно для того, чтобы уничтожить 60 миллиардов мышей, вес которых составляет 1 миллион 200 тысяч тонн. Поэтому главным фактором патогенности возбудителя ботулизма являются продуцируемые им экзотоксины. Хотя они отличаются по своим антигенным свойствам, их биологическая активность одинакова. Все они являются вариантами единого нейротоксина. Установлено, что антигенная специфичность и летальная активность определяются различными детерминантами экзотоксина. Токсины всех известных типов продуцируются в виде токсических белковых комплексов (прогениторных токсинов). В зависимости от их молекулярной массы и структуры эти комплексы делят соответственно константам седиментации на 3 группы: 12 S-, 16 S- и 19 S-токсины. 12 S-токсины (М-токсины) состоят из молекулы нейротоксина, ассоциированной с молекулой нетоксического белка, который не обладает гемагглютинирующими свойствами. 16 S-токсины (L-токсины) — структуры, состоящие из М-комплекса и нетоксического белка, который отличается от белка М-комплекса и обладает гемагглютинирующими свойствами. 19 S-токсины (LL-токсины) — наиболее крупные структуры, включающие в себя нейротоксин и нетоксический белок со свойствами гемагглютинина.
Клостридии ботулизма типа А могут продуцировать токсические комплексы 3 вариантов: М, L и LL, типов В, С и D — в виде L- и М-токсинов, а типов Е и F — только в виде М-токсинов. Таким образом, одна и та же культура C.botulinum может продуцировать несколько типов токсических комплексов. Роль нетоксических белков в составе токсических комплексов заключается в том, что они стабилизируют токсин и защищают его от разрушающего действия протеолитических ферментов и НС1 при пероральном заражении. Роль их в патогенезе ботулизма при парентеральном заражении остается невыясненной. Нейротоксические компоненты любого серотипа ботулинических токсинов и любого типа токсического комплекса имеют сходную структуру и биологические свойства. Они синтезируются в виде единой полипептидной цепи с молекулярной массой 150 кД (78-токсин), которая не обладает значительной токсической активностью. Эта полипептидная цепь превращается в активный нейротоксин только после ее разрезания бактериальной протеазой или протеазами кишечного тракта человека. В результате точечного гидролиза возникает структура, состоящая из двух связанных между собой дисульфидными связями цепей — тяжелой, с м,м. 100 кД (Н-цепь), и легкой, с м.м. 50 кД (L-цепь). Н-цепь ответственна за прикрепление нейротоксина к рецепторам мембраны клеток, а L-цепь осуществляет специфическое блокирующее действие нейротоксина на холинергиче-скую передачу возбуждения в синапсах. Токсины типов С1 и С2 отличаются друг от друга не только серологически, но еще и тем, что токсина С2 вегетативные формы культур не образуют. Он образуется лишь в период спорообразования, а его активация обеспечивается микробной протеазой.
Способность возбудителя ботулизма продуцировать протеолитические ферменты играет важную роль в токсинообразовании. Протеолитические группы возбудителей обеспечивают активацию прототоксинов своими эндогенными протеазами, а активация нейротоксинов, продуцируемых непротеолитическими вариантами серотипов C.botulinum, осуществляется экзогенным путем, т. е. с помощью протеаз желудочно-кишечного тракта при заражении или, in vitro — трипсином.
Помимо выраженной нейротоксической активности, различные типы C.botulinum обладают лейкотоксической, гемолитической и лецитиназной активностью. Особенность лейкотоксина заключается в том, что он подавляет фагоцитоз без разрушения лейкоцитов. Различные сроки накопления в культуральной среде, лейкотоксинов, гемотоксинов и лецитиназы при инкубации C.botulinum указывают на то, что они имеют, по-видимому, разную химическую природу.
Генетический контроль синтеза ботулинических токсинов изучен слабо. Однако получены данные, которые указывают на то, что способность C.botulinum к синтезу нейротоксинов кодируется привнесенными в ее геном генами конвертирующих профагов.
ВНЕШНИЕ ПРИЗНАКИ БОЛЕЗНИ (КЛИНИКА) обусловлены преимущественно действием нейротоксина и включают:
· классическую пищевую токсикоинфекцию, более распространенную под названием «ботулизм»;
· ботулизм новорожденных (у детей от 3 до 20 нед), для которого характерны - генерализованная гипотония и амиотрофия, развивается при заглатывании спор с последующим развитием вегетативных форм (в США ежегодно регистрируют около 70 случаев);
· раневой ботулизм (возникает при загрязнении некротизированных тканей почвой);
· неопределенно классифицируемый ботулизм (у детей старше одного года и взрослых), не связанный с указанными факторами риска (пища, рана).
Остановимся на двух первых формах течения, связанных с пищей.
ПИЩЕВОЙ БОТУЛИЗМ. Ботулизм протекает как токсикоинфекция. Организм поражается не только токсином, содержащимся в пищевом продукте, но и токсином, который образуется в пищеварительном тракте и тканях в связи с проникновением туда возбудителя. Люди чрезвычайно чувствительны к ботулиническим токсинам типов А, В, С, Е и F. Заболевания наблюдались даже тогда, когда человек брал в рот зараженный продукт, но не проглатывал его. Смертельная доза токсина для человека составляет 1 нг/кг массы тела. Ботулинический токсин быстро всасывается в желудке и кишечнике, проникает в кровь и избирательно действует на ядра продолговатого мозга и ганглиозные клетки спинного мозга. Следует отметить, что, попадая в пищеварительный тракт человека или животного, клостридии ботулизма размножаются, проникают в кровь и оттуда во все органы, продуцируя при этом токсины. Инкубационный период у людей варьирует от двух часов до 10 дней, но чаще всего он составляет 18-24 часа. Чем больше инфицирующая доза, тем короче инкубационный период и тем тяжелее протекает заболевание.
КЛИНИЧЕСКАЯ КАРТИНА ботулизма обычно складывается из сочетания различных синдромов, из которых раньше всего проявляется неспецифическая симптоматика (недомогание, общая слабость, головная боль). Могут наблюдаться жжение в желудке, тошнота, многократная рвота, понос. Через несколько часов после начала заболевания неспецифическая симптоматика сменяется специфической – первый признак офтальмоплегический: у больного нарушается аккомодация, неравномерно расширяются зрачки, появляется косоглазие, двоение в глазах, опущение век, а иногда и слепота. Эти симптомы связаны с поражением глазодвигательных нервов. В дальнейшем наступает паралич мягкого неба, языка, глотки, гортани, появляются расстройства речи, нарушается акт жевания и глотания, наблюдается выделение густой тягучей слизи. Вследствие паралича лицевых мышц, выражение лица изменяется иногда до неузнаваемости. Парезы мышц желудка и кишечника ведут за собой резкие нарушения моторной функции кишечника, появление стойкого запора и метеоризма. Отмечается сухость во рту и глотке. температура тела не соответствует частоте пульса - нормальная или ниже нормы; пульс учащен. Начинают преобладать нервно-паралитические явления, как результат расстройства деятельности нервных центров, регулирующих функцию слюнных, слезных желез, мышц носоглотки и лица. К ранним симптомам заболевания относится постепенно развивающееся расстройство зрения. Продолжительность болезни обычно 4-8 суток, реже - 3-4 недели. Смерть наступает, как правило, в результате дыхательной недостаточности от паралича дыхания и сердца при ясном сознании. Летальность составляет 35-85%. Но заболевание не всегда протекает в такой форме.
ДЕТСКИЙ БОТУЛИЗМ. Это новая форма ботулизма, обнаружена в конце 70-х годов ХХ столетия. С тех пор описано более 100 лабораторно подтвержденных случаев детского ботулизма. Из 40 случаев заболевания ботулизмом в США, зарегистрированных в 1979 году, 24 случая были связаны с детским ботулизмом. Детский ботулизм встречается у детей в возрасте от 29 дней до 8 месяцев.
По-видимому, предпосылкой появления клинических симптомов этой формы заболевания является попадание в желудочно-кишечный тракт ботулинических спор в первые недели жизни ребенка. Считают, что детский ботулизм возникает от ботулинического токсина, образуемого непосредственно в кишечном тракте ребенка в результате жизнедеятельности Cl.botulinum. Из 555 проб продуктов, домашней пыли, почвы комнатных растений, проанализированных по поводу детского ботулизма, в 9 случаях были выделены споры Cl.botulinum; ботулинический токсин во всех пробах отсутствовал. По крайней мере, в 34,7% случаях в питание госпитализированных по поводу ботулизма детей входил мед. Жизнеспособные клетки Cl.botulinum типа В были обнаружены в шести различных пробах меда (10%) от тех партий продукта, которые использовались для кормления детей. Первый симптом детского ботулизма - постоянный запор, хотя он часто остается незамеченным. У пострадавших детей отмечается вялость, умеренная слабость, затруднение в кормлении и изменившийся плач.
У сельскохозяйственных животных внешние признаки данного заболевания примерно одинаковы. Отмечается быстрая утомляемость, затруднение движения, нарушение процесса жевания и глотания, ослабление работы сердца, ослабление работы кишечника - запоры. У свиней отмечается потеря голоса, обильное слюнотечение, нарушение координации движения, частичная слепота. Прием корма также невозможен вследствие паралича жевательной мускулатуры и глотки. У птиц при ботулизме наблюдается вялость, они неохотно принимают корм и воду, с трудом передвигаются и предпочитают сидеть нахохлившись, постепенно появляются параличи мигательной перепонки, потом крыльев, ног и шеи. Птица не может держать голову и опускает ее вниз упираясь клювом в землю. Послеубойные изменения в тушах не обладают характерными признаками.
ПРОФИЛАКТИКА БОТУЛИЗМА основана на соблюдении гигиенических требований при приготовлении пищевого продукта. Необходимо знать о том, что этот почвенный микроб может попасть в пищевой продукт, приготовляемый для хранения или консервации вместе с частичками земли, но и то, что в мясные изделия он попадает вместе со специями, пряностями (перец, кардамон и т.д.), поваренной солью и другими добавками. Однако это не исключает и другой путь попадания его в мясной продукт из содержимого кишечника при разделке туши или через инструменты, используемые при той же разделке туши. Возможен и иной путь попадания микробов в мясные продукты - от больных животных или непосредственно через мясо, или через кишечное сырье, используемое для производства колбасных изделий. Поэтому и необходимо, прежде чем убить животное, поведение которого свидетельствует, что оно больное, исключить данное опасное заболевание.
Необходимо помнить, что в мясном продукте, где содержится микроб или токсин их расположение возможно гнездно, в результате чего одни люди после употребления в пищу данного продукта не заболеют, а другие заболеют. Если микроб находится в консервах, то надо знать, что тепловая обработка не уничтожает его споровую форму, и это способствует его дальнейшему прорастанию в вегетативную форму, производству и накоплению ботулинического токсина. Причем при нагревании продукта уничтожаются микробы - антагонисты (те которые препятствуют развитию этой опасной бактерии и накоплению токсина). Кроме того, из продукта при нагревании частично удаляют воздух, то есть создается питательная среда с малым содержанием кислорода, а это идеальные условия для развития и размножения бактерий Cl.botulinum. Чаще всего внешний вид продуктов, в которых образовался ботулинический токсин, изменяется. Они размягчаются, появляется неприятный запах, образуется газ. В консервах возникает бомбаж, т.е. банка вздувается под действием газа продуцируемого микробами. Такие продукты нельзя использовать в пищу или кормить ими животных.
ОЦЕНКА МЯСА от больных ботулизмом животных и пищевых продуктов, содержащих данный микроб или его токсин очень жесткая. Животные, больные ботулизмом к убою не допускаются. Мясо, мясные и другие пищевые продукты, в которых обнаружены бактерии Cl.botulinum или их токсин подлежат уничтожению.
Пищевые отравления, вызываемые кокковой микрофлорой
К данной группе отравлений относятся заболевания, причиной которых послужили токсины, вырабатываемые стафилококками или стрептококками.
ИСТОРИЧЕСКАЯ СПРАВКА. Впервые пищевые отравления, виной которых стали стафилококки, описал П. Н. Лащенков в 1901 году. Им в 1899 году в Харькове изучена вспышка пищевых отравлений после употребления орехового торта с кремом и разработана схема бактериологического и биологического исследования с выделением чистой культуры и контрольных опытах на котятах, щенках, морских свинках и кроликах. Несколько позже (1906 г.), бактериолог Яковлев доказал способность стрептококков вызывать пищевые отравления. В 1909 году Д. Филицин и в 1912 году В. Н. Косарев анализируя вспышки пищевых отравлений на Украине и в Крыму, выдвинули обоснованное предположение о стафилококковой причине некоторых из этих отравлений. За рубежом считается, что первые исследования по изучению этиологической роли стафилококков при отравлениях людей проводились американским бактериологом Барбером (1914 г.). В 1914 году появилось сообщение о коровьем молоке, которое неоднократно вызывало кишечные расстройства у потребителей. Было установлено, что непосредственно после получения молоко было безвредно, но после хранения его в течение нескольких часов в условиях комнатной температуры становилось токсичным. Кокки выделенные из молока, вызывали явления острого отравления при введении вместе с питательной средой, на которой они культивировались. Кокки, вводившиеся без питательной среды через рот щенкам и обезьянам, не давали токсического эффекта.
Это дало основание считать, что желудочно-кишечные расстройства, вызывавшиеся молоком, являются следствием образования в нем токсического вещества в результате жизнедеятельности микроорганизмов после непродолжительного нахождения в условиях комнатной температуры.
Микроорганизмы рода StaphylococcusМикроорганизмы данного рода представлены примерно 30 видами (из них 14 являются симбионтами - паразитами человека).
Данный микроорганизм был обнаружен в 1878 Р. Кохом. Первых микроорганизмов рода выделил из гноя фурункула в 1880 Л. Пастер, в 1881 г., А. Огстон предложил родовое название Staphylococcus. В 1884 из очагов гнойных поражений человека Ф. Розенбах выделил виды S. albus, S. aureus. Стафилококки являются симбионтами человека и животных, всю жизнь человек проводит в среде контаминированной стафилококками, но они не всегда ограничивают свои взаимоотношения с хозяином рамками безвредного сожительства. По мнению А. К. Акатова и В. С. Зуеваой (1983) наиболее древними видами, являются S.xylosus и особенно S.sciuri. Основанием для подобного предположения служат данные об убиквитарности названных видов, до сих пор сохранивших способность к свободному обитанию в абиотических средах - почвы и воды (Kloos, 1980). Одновременно анализ их биохимической активности свидетельствует в пользу возможности размножения S.sciuri и S.xylosus на неорганическом субстрате. Упомянутые виды также обнаруживаются на наружных покровах практически всех млекопитающих, где из них в процессе адаптации к условиям, имеющим место на коже и слизистых оболочках этих хозяев, вероятно, и генерировали прочие виды стафилококков. Поэтому то в результате подобного эволюционного процесса в настоящее время род Staphylococcus и представлен более чем тридцатью различными видами. Следующим шагом на пути эволюции стафилококков — являлось становление их патогенности, о чём свидетельствует их роль в инфекционной патологии человека и животных (Grosserode, Wenzel, 1991; Roberson et aL, 1994). В определённой степени становление патогенности стафилококков можно оценить как пример "случайного паразитизма", при котором сама патогенность может рассматриваться как функция адаптации микроорганизма к организму хозяина: чем она выше, тем больше вероятность развития инфекции (Домарадский, 1997). По мнению Д.Г. Дерябина все стафилококки по своему месту в биоценозе могут быть разделены на антропофитные и зоофитные виды. Основным хозяином первых являются человек и высшие приматы, а вторых — различные виды птиц и млекопитающих. В частности, для наружных покровов тела человека характерно присутствие S.aureus, S.epidermidis, S.capitis, S.hominis и некоторых других видов стафилококков, относительно редко обнаруживаемых у представителей животного мира (Kloos, 1986). Приматы, наряду с человеком являются хозяевами S.haemolyticus, S.warned, S.simulans, S.cochnii (Schleifer, 1986).
У животных и птиц, т е., среди зоофитных видов стафилококков, так же есть своя «специализация». Так, микроорганизмы вида S.gallinarum преимущественно обнаруживаются у птиц, a S. hyicus, S.chromogenes, S.sciuri, S.lentus, S.vitulus и S.caseolyticus у парнокопытных животных, что объясняет достаточно частое обнаружение этих видов стафилококков в молоке и мясных продуктах (Devriese, 1986). Плотоядные хищники контаминированны стафилококками таких видов, как S.intermedius, S.schleifed и S.fells, при этом первые два — типичные обитатели кожных покровов и слизистых оболочек собак и волков (Hesselbarth et a1, 1994; Igimi et a1, 1990), а последний вид специфичен для кошек (Igimi et а1., 1994).
Но столь жесткая классификация вида микроорганизма по виду хозяина вряд ли стабильна. В таблице 2 приведены суммарные данные по связке микроорганизм – хозяин. В частности, перечисление видов стафилококков, изолированных от коз (Valle et а1., 1991), включал не только такие типичные зоофитные виды, как S. chromogenes (6,6 % от общего количества выделенных культур), S.hyicus (3,8 %) S.sciuri (5,8 %) и S.caprae (5,2 %), но и виды стафилококков, устойчиво обнаруживаемых на коже и слизистых оболочках человека: S.aureus (19,7 %), S.haemolyticus (18,5 %), S.wameri (13,0 %), S.epidermidhs (9,2%). Наиболее частыми изолятами у оленей оказываются S.simulans и S.xylosus, однако достаточно часто среди них встречаются представители антропофитных видов — S.epidermidis, S.hominis, S.capitis (Laukova, 1997). В работе S. Igimi с соавторами (1994), посвященной характеристике стафилококков, изолируемых от кошек, помимо относительно специфичного для данного вида млекопитающих микроорганизма S fells (45 % от общего количества выделенных культур) также идентифицированы S.aureus (13 %), S.intermedius (10 %), S. sciuri (6 %), S.epidermidis (6 %), S.simulans (4 %) и еще шесть менее часто обнаруживаемых видов. Достаточно типичный антропофитный вид S.epidermidis в 13,8 % случаев был также изолирован A. Birgersson с соавторами (1992) с вымени коров, а виды S.h hominis и S.capitis от ослов и прочих сельскохозяйственных животных (С.С. Саторов, М.И. Орзуев, 1987). Выше приведённые данные позволяют утверждать о возможности горизонтального переноса представителей отдельных видов стафилококков между различными макроорганизмами.
Возвращаясь к вопросу о роли стафилококков в патологии человека, следует указать на вид S.aureus, который является лидирующим в инициации широкого спектра заболеваний — от "малых" кожных инфекций, тяжелых септических состояний с возможным летальным исходом до пищевых токсикозов. Существует несколько схем дифференциации среди вида S.aureus Наибольшее распространение в мировой практике нашла схема, предлагаемая V. Hajek и Е. Marsaiek (1976).
Описанные ими биотипы: А, В, С и D используются бактериологами и до настоящего времени. Штаммы биотипа А наиболее часто изолировались от людей; биотипа В — от кур, свиней и кроликов; биотипа С — от коров и овец, биотипа D — от зайцев (Акатов, Зуева, 1983). Биотип Е в дальнейшем был выделен в особый вид — S.intermed (Hajek, 1976), а биотип G и нетипируемые группы UT1 и UT2, чаще всего выделяются от крыс (Kato et al., 1995).
По степени убывания своей этиологической значимости в патологии человека виды стафилококков располагаются последующему вектору S.epidermidis (не менее 50 % от общей численности их клинических выборок) --> S.haemolyticus (< 15 %) --> S.hominis (< 10 %) —> S.simulans (< 8 %) --> S.wameri (< 6 %) --» S.capitis (< 4 %) (Schleifer, 1986 b). Спектр патологически значимых стафилококков может быть расширен за счет новых видов, среди которых следует выделить S.lugdunensis и S.schleiferi (Fleurette et al., 1989). Последний представляет достаточно значимую медицинскую проблему в некоторых европейских странах. Необходимо также указать, что некоторые зоопатогенные виды могут быть этиологически значимы и в патологии человека, как например, при выделении S.caprae при костных инфекциях (Shuttleworth et al., 1997), бактериемии и эндокардите (Vandenesch et al., 1995), S.intermedius — при стафилококковой бактериемии у иммуноскомпрометированных пациентов (Vandenesch et al., 1995 а). Последний вид микроорганизмов фиксировался и как причина гнойного процесса при инфицированных собачьих укусах (Talan et al., 1988).
Общепризнанно, что чем выше становится приспособляемость микроорганизма к существованию во внутренней среде организма хозяина, тем более утрачивается его возможность к развитию в других условиях среды обитания.
МОРФОЛОГИЯ И КУЛЬТУРАЛЬНЫЕ СВОЙСТВА МИКРООРГАНИЗМА. Эти микроорганизмы в мазках под микроскопом выглядят как шары, образуя скопления напоминающие гроздья винограда (отсюда и название staphylos – гроздь). Спор они не образуют, значит они не обладают высокой устойчивостью к температурному воздействию, однако из безспоровых форм они одни из самых устойчивых бактерий. Жгутиков у них нет, таким образом в их антигенной формуле отсутствует Н-антиген, а присутствует только соматический антиген самой микробной клетки. Грамположительны. Стафилококки неподвижны; факультативные анаэробы; хемоорганотрофы с окислительным и ферментативным метаболизмом, каталаза-положительны; содержат цитохромы, но обычно оксидаза-отрицательны, чувствительны к действию лизостафина, но не лизоцима (Schleifer, Kloos. 1975). Растут на средах содержащих до15% NaCI, температурный оптимум роста — 35-40°С; предпочтительна слабощелочная реакция среды (7,0-7,5). На плотных средах образуют мутные круглые ровные колонии кремового, желтого или оранжевого цвета. Цвет колоний обусловлен наличием липохромного пигмента, его образование происходит только в присутствии кислорода и наиболее выражено на средах, содержащих кровь, углеводы или молоко. Вызывают характерное разжижение желатина с образованием воронки, заполненной жидкостью (на 4-5 сут.) На жидких средах дают равномерное помутнение, а затем рыхлый осадок, превращающийся в тягучую массу. Восстанавливают нитраты, образуют Н2S, разлагают глюкозу, ксилозу, сахарозу, мальтозу, глицерин, маннит с выделением кислоты; уреаза-положительны; крахмал не гидролизуют; индол не образуют. По наличию коагулазы все стафилококки разделяют на две группы; среди патогенных видов коагулаза-положителен лишь S. aureus. остальные виды называют коагулаза-отрицательными. Типовой вид — S. aureus.
УСТОЙЧИВОСТЬ. Стафилококки хорошо переносят высушивание, сохраняя вирулентность; погибают при прямом воздействии солнечного света в течение 10-12 ч. Довольно устойчивы к нагреванию — при 70-80°С погибают за 20-30 мин; сухой жар убивает их за 2 ч. Менее устойчивы к действию дезинфектантов, но резистентны к воздействию чистого этанола. Кокковые микроорганизмы обладают устойчивостью к внешним воздействиям: способны расти при температуре 6,5-46°С, длительно переносят низкие температуры (-20°С), обладают выраженной солеустойчивостью, размножаются в продуктах с содержанием поваренной соли до 15 - 17%, 40% желчи. Они устойчивы и к воздействию кислой среды, сохраняясь в продуктах, где показатель кислотности достигает величины рН = 3,0 (гораздо ниже, чем в маринаде), возможен рост в пределах рН 4,2-9,3. Однако нельзя забывать, что сама по себе кокковая микрофлора не вызывает пищевых отравлений. Опасен токсин, выделяемый этими микроорганизмами, а его устойчивость значительно превышает устойчивость кокков. Токсин термоустойчив, хорошо сохраняется во внешней среде, устойчив к низким температурам, кислотам. Абсолютный спирт не разрушает его. Токсин полностью не разрушается при нагревании до 100°С в течение 30 минут; он разрушается при кипячении в течение 2 часов, а также автоклавировании при 120°С в течение 30 минут. На размножение кокков и токсинообразование в пищевых продуктах оказывают влияние концентрация соли и сахара. Содержание в продукте до 12% поваренной соли и до 30% сахара не препятствует токсинообразованию. Рост этих микроорганизмов подавляется при концентрации сахара свыше 30% и хлористого натра 20-25%; препятствует накоплению токсина и высокая кислотность продукта.
Дифференцинирование патогенных и непатогенных штаммов. Применяют коагулазный тест на наличие свертывающего фактора, положительный для 95% изолятов, а также определяют способность ферментировать маннит (разлагают); исследуют способность синтезировать термостабильную ДНКазу и агглютинировать частицы латекса или сенсибилизированные эритроциты барана. Последний тест позволяет выявлять белок А и свертывающий фактор либо оба продукта. Следует помнить о возможных ложноположительных результатах при наличии S. epidermidis и микрококков, а также ложноотрицательных, обычных для метициллин (оксациллин) резистентных штаммов S. aureus (MRSA).
Идентификация штаммов с помощью типовых бактериофагов. Метод типирования патогенных стафилококков бактериофагами достаточно широко применяют в клинической эпидемиологии. Для фаготипирования используют стандартный набор из 20 бактериофагов, разделенных на 4 группы: 1-я группа включает фаги 29, 52, 52А, 79, 80, 2-я — 3А, 3С, 55, 71, 3-я — 6, 42Е, 47, 53, 54, 75, 77, 83А, 84, 85, 4-я — 42D. С помощью соответствующих бактериофагов удается типировать 60-80% изолятов: установлены особые штаммы (например, фаготипов 80 и 77), наиболее часто выделяемые при вспышках.
АНТИГЕННАЯ СТРУКТУРА. Серологические исследования (например, идентификация) не имеют принципиального значения, а результаты часто носят противоречивый характер. До настоящего времени реагенты для идентификации TSST-1 и AT к нему остаются слабо доступными.
БОЛЕЗНЕТВОРНОСТЬ (ПАТОГЕННОСТЬ). В основе токсикозов, обусловленных кокками, лежит токсическое начало, т.е. принятие с пищей готовых токсинов. Наличие кокков в пищевых продуктах, способность их размножаться, недостаточны для возникновения пищевого отравления, необходимо образование и накопления в них токсина. Токсины способны продуцировать лишь патогенные кокки, которые обладают энтеротоксигенными свойствами.
Будучи грамположительными микроорганизмами, стафилококки не обладают способностью к образованию эндотоксинов, но продуцируют значительное количество экстрацеллюлярных протеиновых факторов с токсическими свойствами — экзотоксинов. Истинные экзотоксины стафилококков не вызывают лизиса клеток, а воздействуют на их метаболизм через локализованные на цитоплазматической мембране специфические рецепторы. При этом все многообразие стафилококковых экзотоксинов может быть объединено в две неравновеликие группы – эксфолиативные токсины, вызывающие спецефическое поражение кожного покрова и энтеротоксины.
Энтеротоксины. Классический взгляд на энтеротоксины обусловлен рассмотрением их в качестве причины тяжелых пищевых отравлений, вызванных поступлением в желудочно-кишечный тракт экзопродуктов стафилококкового происхождения предварительно накопившихся в продуктах питания (Wieneke et al., 1993). Ведущим продуцентом энтеротоксинов являются микроорганизмы вида S.aureus, однако в единичных случаях этиологическая роль в возникновении вспышек пищевых отравлений может принадлежать и токсинпродуцирующим штаммам S.intermedius (Khambaty et al., 1994). При этом основные симптомы данного заболевания — тошнота, рвота, возможно диарея — развиваются у человека при потреблении чрезвычайно малых доз токсина (от 1 мкг).
При схожести клинических проявлений стафилококковых токсикозов, за их возникновение ответственно целое семейство молекул, по своим антигенным свойствам подразделяемых на несколько серологических типов: А, В, С (C l, C 2 и С З — варианты) и D. Относительно недавно были описаны также стафилококковый энтеротоксин Е (Brehm, Tranter, 1993) и энтеротоксин H (Su, Wong, 1995). На обнаружении антигенных различий между энтеротоксинами основаны и традиционные подходы к их выявлению в продуктах питания с использованием иммуноферментных и радиоиммунных методов (Park et al., 1994). В то же время все названные экзотоксины имеют ряд общих характеристик: 1) их молекулы массой от 26 до 34 kDa состоят из одной полипептидной цепи, включающей 239—296 аминокислотных остатков и имеющей одну дисульфидную связь; 2) общим является существование нескольких обусловленных конформационными изменениями изоэлектрических форм каждой из молекул; 3) для них характерна устойчивость к действию высоких температур и протеолитических ферментов, что обусловливает сохранение активности токсинов в продуктах питания при их термообработке, а также в желудочно-кишечном тракте (Marrack, Kappler, 1990).
Действие стафилококковых энтеротоксинов при пероральном введении аналогично действию энтеротоксинов, образуемых микроорганизмами иных таксономических групп, т е нарушается выход из клеток-мишеней воды и ионов хлора и поступление внутрь них ионов калия.
Накопившиеся к началу 80-х годов данные о летальных случаях септицемии, раневых, и других инфекций стафилококковой этиологии, многие из которых протекали с синдромом выраженной интоксикации по-иному заставили взглянуть на механизм действия энтеротоксинов и их патогенетическое значение при стафилококковых инфекциях. Клинические же проявления интоксикации не были связаны рвотой или диареей, но заключались в резком повышении температуры, нарушении функции печени и почек, понижении артериального давления и в ряде случаев — необратимом падении сердечной деятельности. В эти же годы сходное заболевание - синдром токсического шока (СТШ) описано у женщин в период менструального цикла (Ратгауз, Акатов, 1986), когда из влагалищного отделяемого были изолированы культур S.aureus, продуцирующие ранее неизвестный токсин — TSST, по своей молекулярной организации, сходный с прочими стафилококковыми энтеротоксинами (Kum et a1, 1993).
Изучение механизмов развития синдромов токсического шока позволило выявить у стафилококковых энтеротоксинов и TSST способность к связыванию с главным комплексом гистосовместимости на поверхности антигенпрезентирующих клеток, что в дальнейшем ведет к поликлональной стимуляции Т-лимфоцитов (Marack, Kapk 1990), а также вызывает индукцию выброса иммунокомпетентными клетками широкого спектра цитокинов (Kum et al., 1993). При этом было продемонстрировано, что активация Т-лимфоцитов совместно с цитокиновым "взрывом" и развитие шока являют патогенетически зависимыми феноменами (Bonventre et a 1993).
Таким образом, в соответствии с современными представлениями, у большинства стафилококковых экзотоксинов (СЕ, СЕВ, СЕС, TSST и др.) могут быть продемонстрированы два принципиально различных свойства: 1) энтеротоксичность - проявляющаяся в развитии рвоты и диареи при пероральном введении нескольких миллиграмов этих протеинов; 2) способность при попадании в кровеносное русле в дозе от 1 пкг/мл до 1 мкг/мл индуцировать Т-клеточную активацию и выработку ряда цитокинов (Yokomizo et al., 1995). Последнее свойство - активации Т-клеточного звена иммунитета, дало данному феномену название "суперантигенной" стимуляции. Соответственно, сами стафилококковые экзотоксины, способные качественно изменять иммунные реакции организма хозяина, были оценены как семейство структурно и функционально близких белков — суперантигенов (Johnson et al, 1991).
Открытие суперантигенов, обнаруженных помимо стафилококков у стрептококков явилось одним из наиболее ярких событий в микробиологии конца XX века, значительно изменивших представления о патогенезе заболеваний инфекционной природы.
Оптимальными условиями образования в мясе, фарше и печени токсина стафилококков, является хранение продуктов при температуре 28-37°С и рН среды 6,6-7,2; при температуре ниже 20°С и рН среды 6,5-6,0 образование токсина замедляется, а при температуре ниже 15°С и рН среды 6,0-4,5 прекращается. Установлено, что при одинаковых условиях хранения кокки и токсин накапливаются в вареном мясе и печени в 2-3 раза, а в вареном фарше в 5-7 раз быстрее, чем в цельных кусках сырого мыса и печени. Быстрота выработки токсина и его активность не всегда параллельны скорости размножения микробов. Стафилококк хорошо развивается, продуцируя токсин, на самых различных пищевых продуктах. Содержание в продукте до 12% поваренной соли и до 50% сахара не препятствует размножению стафилококков и токсинообразованию. Образование токсина происходит лучше при пониженном парциальном давлении кислорода. В лабораторных условиях для получения токсина культуру выращивают в эксикаторе при 20% СО2. Размножение стафилококков и токсинообразование могут быть и в анаэробных условиях, в частности в консервах (В.И. Тец 1958).
Развитие стафилококка на пищевых продуктах и накопление здесь энтеротоксина не вызывает заметного изменения органолептических свойств этих продуктов.
Количество токсина, способное вызвать интоксикацию, накапливается в мясном фарше, содержащем 50% белого хлеба, при комнатной температуре через 4-5 часов, а в готовых котлетах - через 3 часа. Добавление белого хлеба к фаршу в качестве наполнителя значительно ускоряет образование токсина. Пример 50% штаммов золотистого стафилококка (чаще всего фагогруппы 111) продуцируют токсины вызывающие гастроэнтерит с тяжёлой рвотой и профузным поносом. Более половины этиологически расшифрованных пищевых отравлений в развитых странах мира связаны со стафилоккоком.
Пожалуй, ни один из видов бактерий не может конкурировать со стафилококком по числу факторов патогенности. Факторами патогенности стафилококков, помимо токсинов, являются так же: микрокапсула, компоненты клеточной стенки, ферменты.
Микрокапсула защищает бактерии от комплемент-опосредованного поглощения полиморфноядерными фагоцитами, способствует адгезии микроорганизмов и их распространению по тканям. При выращивании in vitro обычно не образуется.
Компоненты клеточной стенки стимулируют развитие воспалительных реакций: усиливают синтез интерлейкина макрофагами, активируют систему комплемента. Тейхоевые кислоты облегчают адгезию к эпителиальным поверхностям. Белок А (агглютиноген А) активирует компоненты комплемента. Активация комплемента приводит к проявлению различных местных и системных реакции, например анафилаксии, феномена Артюса, угнетению активности фагоцитов и т.д.
Ферменты проявляют разнонаправленное действие: каталаза защищает бактерии от действия механизмов фагоцитоза; b-лактамаза разрушает молекулы b-лактамовых антибиотиков; липазы облегчают адгезию и проникновение в ткани. Коагулаза, существующая в 3 антигенных формах, вызывает свертывание сыворотки; сам фермент не взаимодействует с фибриногеном, а образует тромбиноподобное вещество, предположительно взаимодействующее с протромбином.
Гемолизины. Золотистые стафилококки способны одновременно синтезировать несколько подобных продуктов, в частности выделяют 4 антигенных типа гемолизинов (a-Гемолизин (a-токсин), b-Гемолизин (сфингомиелиназа), g-Гемолизин, d-Гемолизин, вызывающие полный гемолиз кровяных сред; кожные некротические реакции и гибель животных после внутривенного введения ( Классификация факторов патогенности дана в редакции В.И. Покровского, О.К. Поздеева 1998).
Микроорганизмы рода Enterococcus.Данные стрептококки впервые обнаружены в тканях человека при рожистом воспалении и раневых инфекциях (Бильрот, 1874), септицемиях и гнойных поражениях (Пастер, 1879, Огстон, 1881); в чистой культуре их выделили Феляйзен (1883) и Розенбах (1884).Экологически стрептококки делятся на сапрофитов, участвующих в процессах молочнокислого брожения и имеющие промышленное значение и симбионтов обладающих широким спектром болезнетворности. По предложению Ребекки Лэнсфилд (1933) стрептококки классифицируют по наличию специфических углеводов в клеточной стенке, выделяют 17 серогрупп, обозначаемых заглавными латинскими буквами (по специфичности белковых Аг М, Р, и Т стрептококки внутри групп разделяют на серовары) Также используют классификацию Брауна (1919), основанную на особенностях роста на агаре с кровью барана. Соответственно выделяют a- (дают частичный гемолиз и позеленение среды), b- (полностью гемолизирующие) и g- (дающие визуально невидимый гемолиз) стрептококки, основными возбудителями болезней человека являются b-гемолитические виды, большая часть которых относится к серогруппе А. Гемолизирующие энтерококки также способны вызывать пищевые отравления и дисбактериозы кишечника.
МОРФОЛОГИЯ И КУЛЬТУРАЛЬНЫЕ СВОЙСТВА, Микроорганизмы рода Enterococcus образуют овальные бактерии размером 0,6-2,0´0,6-2,5 мкм, в мазках культур, выращенных на жидких средах, располагаются парами или короткими цепочками. Спор не образуют, некоторые виды ограниченно подвижны (имеют небольшие жгутики), капсул не имеют. Факультативные анаэробы; хемоорганотрофы (метаболизм ферментативный); расщепляют различные углеводы с образованием кислоты (преимуществен молочной) без газа. Пищевые потребности сложные, каталаза-отрицательны, в редких случаях восстанавливают нитраты. Растут в интервале 10-45°С (оптимум 37°С) Типовой вид — E. faecalis. Выделение возбудителя обычно не представляет трудностей, т.к. энтерококки хорошо растут на простых средах; на кровяном агаре могут давать зоны полного (редко) или неполного гемолиза. Через 24 ч энтерококки образуют сероватые колонии диаметром 0,4-1 мм; признаками, дифференцирующими их от зеленящих стрептококков, являются способность расти на средах, содержащих 6,5% NaCI, а также способность изменять окраску лакмусового молока или молока с этиленовым синим через 4-6 ч при 37°С.
РАСПОСТРАНЕНИЕ. Широко распространены в природе, обитают в кишечнике различных позвоночных, вызывают нагноения ран, бактериемии и поражения мочевыводящей системы. Ранее микроорганизмы систематизировали как стрептококки группы D (некоторые также реагируют с антисыворотками к группе Q). У человека наиболее часто поражения вызывают E.faecalis, E.faecium и Е. Durans. Энтерококки входят в состав микрофлоры ротовой полости, кишечника и мочеполовой системы; так, E.faecium выделяют из испражнений у 25% клинически здоровых лиц. Большинство инфекций, вызванных энтерококками, носит эндогенный характер и обусловлено инвазией микроорганизмов при избыточной колонизации; также показана возможность нозокомиальной передачи микроорганизмов, частота подобных инфекций возрастает на фоне высокой частоты применения цефалоспоринов широкого спектра действия.
ВНЕШНИЕ ПРИЗНАКИ ОТРАВЛЕНИЯ (КЛИНИЧЕСКАЯ КАРТИНА), кокковых токсикозов у людей обычно проявляется сравнительно быстро после приема пищи (через 2-6 ч.) и сопровождается следующими симптомами: головной болью, головокружением, общей слабостью, тошнотой, рвотой с судорожными позывами, поносом, болями в подложечной области, иногда озноб; в тяжелых случаях упадком сердечной деятельности, судорогами, ослаблением зрения и потерей сознания с падением кровяного давления, иногда повышением температуры до 38,5°С. Выздоровление обычно наступает через 1-3 суток. Смертельных случаев почти не наблюдают. Возможно и иное проявление болезни: внезапное начало, боли в животе, жидкий стул несколько раз в сутки, нередко с кровью, иногда тошнота и рвота, головная боль, слабость. Это зависит от характера токсинов выделяемых кокковой микрофлорой.
ПРОФИЛАКТИКА БОЛЕЗНИ строится на предупреждении заноса в пищу токсинообразующих кокков. Источники инфицирования пищевых продуктов кокками весьма разнообразны. Среди них одно из видных мест занимают животные, болеющие маститом и дающие зараженное кокками молоко. Токсигенные кокки в молоке коров и овец с клинически выраженным маститом обнаруживаются в 50-59% случаев, а в 32-33% - со скрытой формой мастита и в 6-12% - у клинически здоровых животных. Наличие кокков в молоке здоровых коров может быть связано со скрытыми формами мастита или бактерионосительством, которое у коров может продолжаться от 3 недель до 16 месяцев после выздоровления.
Токсигенные штаммы стафилококков обнаруживают в органах и тканях животных убитых на мясо вследствие переболевания травматическим перикардитом, разных видов воспаления легких, маститах, эхинококкозе и фасциелезе печени.
Одним из источников инфицирования продуктов питания является человек, токсигенные кокки у которого могут находиться на слизистой носа, в зеве, в гортани при заболеваниях верхних дыхательных путей, а также на руках при наличии гнойниковых поражений. Около 50% здоровых людей являются носителями патогенных кокков и они могут заражать готовые пищевые продукты.
Описаны вспышки стафилококковой интоксикации, вызванные употреблением в пищу, молока и молочных продуктов - творога, сырковой массы, сыра, сметаны, мороженого, которые свидетельствуют, что при изготовлении различных молочных продуктов токсины могут не разрушаться и длительно сохраняться (в сыре до 6 мес.). Частой причиной кокковых отравлений являются кондитерские изделия с кремом (торты, пирожные) и мясные продукты (студни, паштеты, котлеты, колбасы, блинчики с мясом и др.), содержащие эти микроорганизмы.
ВЕТЕРИНАРНО-САНИТАРНАЯ ОЦЕНКА МЯСА И ПИЩЕВЫХ ПРОДУКТОВ, СОДЕРЖАЩИХ КОККОВУЮ МИКРОФЛОРУ. Она зависит от типа выделенных кокков. Пищевые продукты, являющиеся причиной стафилококковых интоксикаций, как правило, не обнаруживают никаких органолептических (по внешнему виду, вкусу, цвету, запаху и консистенции) отклонений от нормы. Поэтому если животное перед убоем болело маститом или у него был какой-либо воспалительный процесс, возможно гнойная рана, то необходимо после убоя послать образцы мышечной ткани и внутренних органов в бактериологическую лабораторию. Если там определят, что мясо обсеменено токсигенными кокками, то его необходимо разрубить на куски по 2-2,5 кг. и проваривать до 3-х часов или направить на мясокомбинат для изготовления колбасных хлебов (которые варятся по более высокой температуре; чем колбасы), внутренние органы, из которых выделены токсигенные кокки, уничтожаются.
Если токсигеные штаммы кокков выделили из готового пищевого продукта, то данный продукт направляют на утилизацию.
Пищевые отравления бактериями рода BacillusВызвать пищевое заболевание человека, с клиникой и патогенезом отравления, могут и бактерии рода Bacillus включающие 48 видов, это в первую очередь виды Bacillus anthracis и Bacillus cereus. Несмотря на длительную историю изучения болезней вызываемых Bacillus anthracis и Bacillus cereus, мы до настоящего времени ощущаем неполноценность своих знаний по биологии указанных микроорганизмов. В данной работе сделан акцент на сибирскую язву как на пищевую инфекцию, где токсины являются одним из факторов, вызывающих клиническое проявление болезни и заражение идёт посредством употребления пищевых продуктов контаминированных Bacillus anthracis.
Пищевые отравления, вызываемые бактериями BAC
Период активного изучения сибирской язвы (антракса) насчитывает уже около 200 лет. Исследования проводились во многих странах и в многочисленных исследовательских институтах и лабораториях. История изучения болезни пестрит громкими именами - Л. Пастер, Р. Кох, Л.С. Ценковский, Ф.А. Брауэль, Н.А. Михин, Ф.А. Тсрентьев, С.Г. Колосов, Я.Е. Коляков и многие другие.
Однако научная мысль постоянно возвращается к этому интересному объекту, продолжая открывать новые стороны этой «старой» болезни.
Что такое сибирская язва (антракс), в наше время знает, пожалуй, не только каждый врач, но и большинство простых граждан. Болезнь известна с древности. Однако широко используемые ссылки на древнегреческих авторов требуют уточнения. Так, R.Pfisterer (цит. по Ргос. Inter. Work. shop, Winchester, 1996) сообщает, что «в каждом историческом обзоре, опубликованном по сибирской язве (антраксу), говорится, что слово «антракс» использовали Гомер и Гиппократ в упоминании о болезни. Тщательное изучение оригинальных текстов показывает, что Гомер не употреблял слово «антракса при ссылке на болезнь. Это слово можно найти только один раз в его произведении «Илиада» и оно означает «горящие угли». В текстах Гиппократа «антракс» также означает, за небольшими исключениями, «горящие угли». В некоторых из этих текстов «антракс» определяет различные неспецифичсские везикулярные или «прыщавые» дерматиты без каких-либо показаний на контакт с больными животными. Дифференциация между «доброкачественным и злокачественным антраксом» сделана много позже».
Тем не менее заболевание известно с глубокой древности, со времен Галена, Цельса и Buргилия болезнь фигурирует под названием «священный огонь» (ignis sacer) или «персидский огонь» (ignis persicus). Русские врачи Колывано – Воскресенских заводов на Алтае А. Эшке (1758) и Н. Ножевщиков (1762) представили в медицинскую коллегию подробные сведения о данном заболевании, включая клинику, эпидемиологию и связь с аналогичной болезнью у животных. В 1766 г. Моран доложил о данной болезни в Академии наук в Париже, что является первой научной работой по сибирской язве за рубежем. В 1769 г. Фурнье выделил сибирскую язву в отдельную нозологическую единицу. С. С. Андриевский, изучавший заболевание во время эпидемии на Урале (1786-1788) дал ему название «сибирская язва», а в 1788г. путем самозаражения доказал единство этиологии сибирской язвы у людей и животных. Возбудитель впервые описали Поллендер, Брауэлл (1849) и Давэн (1850) Чистую культуру возбудителя описал профессор Дерптского русского ветеринарного училища Ф. Брауэль (1857-1858). В своих опытах ему удалось заразить различных животных. Кох (1876) предложил питательные среды для размножения данных микроорганизмов, а в 1881г. Пастер предложил живую вакцину для иммунопрофилактики заболевания. РАСПОСТРАНЕНИЕ. Сибирская язва — типичный зооантропоноз; среди животных наиболее восприимчивы травоядные, но отмечены случаи заболевания среди зайцев, кошек и собак. Наиболее интенсивные очаги заболевания находятся в Азии (Турция, Иран, Китай, Монголия, Индия), Южной Африке, Южной Америке (Аргентина) и Австралии; спорадические случаи регистрируют в Европе, России и США. Ежегодно сибирской язвой заболевает около 1 млн. животных и регистрируют около 40 тыс. случаев заболевания у людей. Животные заражаются при заглатывании спор во время выпаса или при поедании загрязненных кормов. У животных преобладают ангинозная, карбункулезная, кишечная и септическая формы заболевания, т.е. возбудитель проникает через микротравмы ротовой полости или стенку кишечника. Больные животные выделяют сибиреязвенные палочки с мочой и испражнениями. Болезнь быстро прогрессирует в течение 2-3 сут, а при молниеносных формах – в течение нескольких часов; летальность достигает 80%. Клинические признаки болезни (судороги, диарея с кровью) проявляются непосредственно перед гибелью животного. Человек заражается и при употреблении в пищу мяса больных животных. Пути заражения — заглатывание и проникновение в желудочно – кишечный тракт человека.
МОРФОЛОГИЯ БАКТЕРИИ, Крупная толстая палочка (5-10´1-2 мкм) с закругленными концами (при образовании цепочек — с обрезанными под прямым углом концами); неподвижна (абсолютный дифференциально-диагностический признак); легко окрашивается по Граму (грамположительна) и анилиновыми красителями. В клиническом материале располагается парами или в виде коротких цепочек, окруженных общей капсулой; на питательных средах образует более длинные цепочки; при этом морфология палочек несколько изменяется — oни слегка утолщены на концах и образуют сочленения («бамбуковая трость»). Подобные морфологические изменения еще более явно проявляются при температурной фиксации для окрашивания по Граму. Обработка культур пенициллином приводит к разрушению клеточных стенок и образованию цепочек, состоящих из протопластов («жемчужное ожерелье»). Для защиты от факторов резистентности (фагоцитов, AT) образуют капсулы, наблюдаемые только у бактерий, обитающих в живых организмах либо на средах, содержащих нативную сыворотку (например, среда ГКИ). Капсулы более устойчивы к действию гнилостной микрофлоры, чем микробные тела, и в материале из разложившихся трупов нередко можно обнаружить лишь пустые капсулы («тени» микробов). Для более быстрого обнаружения капсул, можно окрасить мазки полихромным метиленовым синим Леффлера (клетки синие, капуслы красно-малиновые). Образуют центрально расположенные эндоспоры, для чего необходимы кислород и определенная температура (12-42°С); в живом организме спор не образуют; также не образуют спор в невскрытых трупах, что опосредовано поглощением свободного кислорода в процессе гниения. Споры отличает высокая устойчивость к внешним возденействиям; в воде сохраняются до 10 лет, в почве — до 30 лет (возможно и дольше). Споры Bacillus anthracis овальной формы размером 0,8-1,0 ´ 1,5 мкм; сильно преломляют свет. Они очень легко образуются на бедных питательных средах, а при свободном доступе кислорода — даже в дистилляте или нефиксированных мазках (последнее следует иметь в виду при работе с вирулентными штаммами). На плотных средах спорообразование идет быстрее, чем на жидких; через 32-48 ч при 37°С оно бывает практически полным. Прекращается полностью при 15°С и 42-43°С. Скорость прорастания зависит от температуры (оптимум 37°С) и возраста спор; молодые споры в оптимальных условиях прорастают за 1-1,5 ч, старые – за 2-10 ч.
КУЛЬТУРАЛЬНЫЕ СВОЙСТВА. Bacillus anthracis хорошо растет на обычных питательных средах: бактерии можно даже выращивать на сыром или вареном картофеле, настое соломы, экстрактах злаков и бобовых. Температурный оптимум на агаре 35-36°С, оптимум рН 7,0. На жидких средах растет в виде ватных хлопьев, взвешенных комком, не вызывает помутнения среды. Дает характерный рост при посеве уколом в желатин («перевернутая елочка»); позднее верхний слой желатина разжижается, образуя воронку. На твердых средах образует крупные шероховатые серовато-белые колонии (R-формы) диаметром 2-3 мм, типичными считают характерные волокнистые колонии («голова Медузы» или «львиная грива»), образованные переплетающимися цепочками бактерий. Рост вирулентных штаммов на плотных средах и желатине настолько характерен, что может служить диагностическим признаком. На свернувшейся (30 мин при 80°С) лошадиной сыворотке растет в виде гладких прозрачных S-колоний, тянущихся за петлей. Палочка сибирской язвы — аэроб или факультативный анаэроб; в анаэробных условиях (либо при значительном варьировании температурного режима) образует единичные, мелкие колонии. При культивировании в микроаэрофильных условиях всегда образует гладкие (S), слизистые (М) или смешанные (SM) колонии. Возбудитель образует кислоту без газа на средах с глюкозой, фруктозой, мальтозой и декстрином. Слабое или отсроченное образование газа наблюдают на средах с глицерином и салицином (не у всех штаммов). Кислотообразования не находят на средах с арабинозой, рамнозой, маннозой, галактозой, раффинозой, лактозой, инсулином, маннитом, дульцитом, сорбитом и инозитом. Гидролизует крахмал; образует ацетилметилкарбинол и лецитиназу. Bacillus anthracis очень медленно и слабо коагулирует жидкую желточную среду; нередко изоляты вообще лишены такой способности. Положительный результат можно ожидать не ранее 5-7 сут инкубирования при 37°С; в то же время сапрофитные бациллы разлагают ее за 6-10 ч. В отличие от сапрофитов, палочки сибирской язвы лишены фосфатазы и не разлагают фосфаты, содержащиеся в питательной среде. Молоко свертывают за 3-5 сут; затем сгусток медленно пептонизируется и разжижается; выделяется аммиак и (в связи с окислением тирозина) накапливается бурый пигмент.
АНТИГЕННАЯ СТРУКТУРА. Выделяют 3 группы основных Аг Bacillus anthracis, две из которых (капсульные Аг и токсин) кодируются плазмидами; отсутствие одной или обеих делает бактерии авирулентными. Капсульные Аг заметно отличаются по химической структуре от прочих капсулярных Аг бактерий; представлены полипептидами. Соматические Аг представлены полисахаридами клеточной стенки.
ПАТОГЕННОСТЬ Bacillus anthracis прямо зависит от капсуло- и токсинообразования; штаммы, не проявляющие подобные свойства, обычно авирулентны. Капсула защищает бактерии от действия вне- и внутриклеточных продуктов фагоцитов и препятствует поглощению бактерий; на начальных этапах заболевания капсула — важнейший фактор вирулентности. Способностью к капсулообразованию также обладают некоторые вакцинные штаммы (например, Пастера или второй вакцинный штамм Ценковского). Долгое время считали, что патологические изменения и гибель наступают в результате действия какого-то гемолитического яда, однако выделить его не удавалось. В 1954 г. появились первые сообщения о выделении сибиреязвенного токсина (in vitro, in vivo). Smith и др. писали, что Вас. anthracis продуцирует токсин in vitro. В 1969 г. почти одновременно были опубликованы работы американских и английских ученых о выделении трех типов токсина. Английские ученые условно назвали их факторами 1, II, III, американские соответственно EF (фактор отека), РА (защитный или протективный антиген) и LF (фактор гибели). Эти факторы различаются серологически. Например, EF и РА синергически вызывают отек у морских свинок и кроликов после подкожной инъекции, а LF и EF — гибель всех животных, восприимчивых к сибирской язве.
Прогресс в области биохимических исследований токсина Bac.anthracis и механизма его патогенетического воздействия на животные клетки во многом связан с работами S. Lеppla (1982-1989). Автору удалось получить в препаративных количествах все 3 компонента токсина в высокоочищенном состоянии, что позволило использовать их как для характеристики функциональной активности, так и для разработки эффективных средств диагностики.
Механизм действия сибиреязвенного (антракс) токсина. До последнего времени оставался не вполне ясным вопрос о механизме взаимодействия PA, LF и EF с животными клетками, расшифровка которого могла бы во многом способствовать пониманию интимной природы патогенетического действия сибиреязвенного (антракс) токсина. Существенные успехи в этом направлении были достигнуты с началом использования культур эукариотических клеток. В 1982 и 1984 гг. S. Leppla опубликовал результаты изучения биохимической природы EF сибиреязвенного микроба. Автору удалось показать, что EF Bac. anthracis вызывает морфологические изменения в монослое культуры клеток ряда клеточных линий. В 1986 г. A. Friedlander показал, что добавление РА и LF к монослою перитонеальных макрофагов приводит к его деструкции при отсутствии таковой в случае использования отдельно взятых РА и LF. Отмеченный факт послужил основанием для выдвижения предположения, что LF, так же как и EF, обладает ферментативной активностью, точка приложения которой, однако, остается невыясненной. Интересно, что, как и в случае с отечным токсином, для реализации цитотоксического действия летального токсина требуются ионы «Са».
Необходимо также отметить, что из 3 указанных компонентов токсина лишь иммунизация РА предохраняла лабораторных животных от гибели при их заражении вирулентными штаммами Bac.anthracis, что позволило использовать в ряде стран препараты на его основе в качестве химической сибиреязвенной вакцины. Данные ряда исследований позволяют отнести сибиреязвенный (антракс) токсин к категории известных бифункциональных (бинарных) токсинов, механизм действия которых в определённой мере укладывается в рамки так называемой А-В-структурной модели, описанной для токсинов Cl. tetani,V. cholerac, C. Diphtheriae (K.Sandvig et al., 1982), экзотоксина А и цитотоксина псевдомонад (D.Fizgerald et al., 1982; T.Sasaki et al., 1985), шига-токсина, а-токсина стафилококков, энтеротоксинов Cl рerfringens (Ю.В.Езепчук.1985; I. Freer, 1988). В качестве В-субъединицы (от англ. binding - связывание) в данном случае выступает протеолитически активированный РА, а в качестве субъединицы А (каталитической, активной) - LF или EF.
Полученные данные свидетельствуют о том, что цитотоксическое действие летального токсина Bac.anthracis реализуется на уровне внутриклеточной мишени, природа которой остается неизвестной (K.Sandvig et al., 1986).
Несмотря на разнообразие и неоднородность изменений, вызываемых обоими компонентами сибиреязвенного токсина, в их основе лежат единые регуляторные механизмы. R.Liddington et al. (1998) провели кристаллографическое изучение летального токсина Bac.anthracis. Оказалось, что она не имеет сходства с другими бактериальными токсинами и определяет новый структурный класс. Летальный фактор является каталитическим компонентом летального токсина Bac.anthracis.
Ph.Hanna (1998) указывает, что после проникновения спор антракса в организм происходит их размножение, рост вегетативных клеток и экспрессия летального токсина и других факторов вирулентности. Летальный токсин Bac.anthracis (Leix) является ответственным за большинство патологий, наблюдаемых при системной сибиреязвенной инфекции. Введение стерильного Leix подопытным животным проявляется в виде шока и внезапной смерти, как это происходит в течении активных бактериальных инфекций. При заражении в организме сразу накапливается Leix в высокой концентрации. Разрушение бактерий при применении антибиотиков чаще всего не приносит успеха. Считается, что Leix секретируется в кровяное русло и свободно циркулирует по организму, проникая в клетки хозяина. Непосредственно в цитоплазме летальный токсин нарушает нормальные гомеостатические функции. Макрофаги являются уникально чувствительными типами клеток, которые, по-видимому, служат жизненными глобальными медиаторами патологий, индуцируемых токсинами. От взаимодействия протективного антигена и летального фактора токсина Bac.anthracis зависит цитотоксичность этого токсина. По данным Y. Sigh и S.Lеppla (1998), в процессе цитотоксичности РА связывается с поверхностным клеточным рецептором и LF транслоцируется в цитозоль клеток. В другой работе те же авторы пишут, что главную роль, которую играет токсин в патогенезе антракса, является то, что токсин эволюционировал в эффективную систему для переноса его двух каталитических белковых, компонентов - фактора отека и летального фактора в цитозоль клеток хозяина. А.С.Степанова и S.Leppla (1996) считают, что взаимодействие летального токсина и фагоцитов животного играет центральную роль в вызывании серьезных симптомов сибирской язвы и летальности.
По данным этих авторов, макрофаги являются мишенью для сложного воздействия обоих факторов вирулентности. Летальный токсин и капсула, при определенных условиях, оказывается, действуют главным образом внутриклеточно, значительно снижая жизнеспособность макрофагов. Эти данные показывают решающую роль бактериального фагоцитоза на начальных стадиях патогенеза сибирской язвы, где фагоциты могут служить в качестве резервуара для аккумуляции, размножения и рассеивания микроорганизма по телу, играя, таким образом, роль в установлении и последующем прогрессировании болезни. На конечных этапах болезни аккумулированные и освобожденные токсины играют первичную роль.
По мнению Т. Koehler, R.Blaustein, A.Fmkelstem, R.Collier (1990), РА компонент антракс токсина играет центральную роль в проявлении интоксикации. Он взаимодействует с мембранным рецептором; подвергается протеолитической обработке; является посредником между эндоцитозом и последующей транслокацией других белков токсина, EF и LF в цитозоль. Путь, по которому токсин, связанный с клеточной поверхностью, достигает цитоплазмы, не известен. За последние годы обнаружено, что токсин Вас. anthracis играет решающую роль в патологии и гибели макроорганизма, независимо от способа введения.
Пищевые отравления, вызываемые бактериями BAC.CEREUSВызвать пищевое отравление человека могут и бактерии Bacillus cereus.
ИСТОРИЧЕСКАЯ СПРАВКА. С середины шестидесятых годов в литературе появились данные о пищевых отравлениях, вызываемых бактериями рода Baillus и, в частности, Bac.сеreus. Описано несколько случаев отравления, причиной которого послужил жареный рис, содержащий указанные бактерии. Позднее наблюдали случаи вспышек пищевого отравления после употребления студня, содержащего бактерии того же вида. В 1979 году было описано заболевание 28 пациентов госпиталя, употребляющих в пищу мясо индейки, из которого впоследствии микробиологи выделили данный микроорганизм. В обзоре Foot Technol за 1988 год Bac.cereus вошел в список 10 основных инфекционных агентов, вызывающих кишечные заболевания людей в Северной Америке. По мнению W. Sperber (1991г.) бактерии Bac.сereus входят в группу из 4-х наиболее опасных микроорганизмов – источников пищевого отравления людей.
МОРФОЛОГИЯ И КУЛЬТУРАЛЬНЫЕ СВОЙСТВА. Bacillus cereus (от лат. cera — воск, свеча) широко распространена в природе, морфологически сходна с Bacillus anthracis (характерно расположение микробных тел в виде штакетника). Bac.cereus подвижная бактерия, значит обладает Н-антигеном, по которому различают более 20 серологических вариантов. По 0-антигенам в группе B.cereus-В.thiiringiensis выделено 13 серотипов. Микроорганизм не обладает капсулой, но образует споры - устойчив к воздействию высокой температуры. Грамположительный.
Температурный оптимум роста 30°С, оптимум рН 7-9,5. На МПА образует «распластанные» колонии с неровными краями; на КА колонии «распластанные», зернистые, с широкой зоной гемолиза, на яичном агаре образует широкую зону преципитации белого цвета (эффект бактериальной лецитиназы), зона быстро растет и через несколько суток инкубации охватывает всю поверхность чашки. Колонии, выросшие на агаре, имеют характерный восковидный вид. На жидких средах бацилла образует белый хлопьевидный осадок, нежную пленку на поверхности и вызывает помутнение бульона. Следует отметить высокую протеолитическую активность микроорганизма — разжижает желатин в течение 1- 4 суток (80% штаммов); гемолизирует эритроциты барана, все штаммы образуют лецитиназу и ацетилметилкарбинол, расщепляют до кислоты глюкозу и мальтозу, а часть штаммов — также и сахарозу, глицерин, лактозу, галактозу, инулин, дульцит и декстрин
УСТОЙЧИВОСТЬ. Данный микроорганизм выделяют практически из всех пищевых продуктов, муки и теста, мясных продуктов, из практически всех видов специй (лавр, перец, сухой чеснок и т.д.). Бактерии широко распространены в природе. Основная среда обитания это почва, оттуда микроб попадает в воздух и воду. Вегетативная форма бактерий отмирает при воздействии температуры в 70°С. При температуре холодильника, бактерии не размножаются, при низких температурах (до –20°С) сохраняет жизнеспособность до 4-х месяцев. Споровая форма бактерий выдерживает режимы пастеризации, температуру в 100°С выдерживает 1 час, в 125°С – 10-13 минут. Микроб может сохраняться в широком диапазоне показателя кислотности - от величины рН равной 4,0 и выше, то есть процессы консервирования методом маринования на него не действуют. Эти бактерии хорошо переносят значительные концентрации нитритов, сахара, коптильных жидкостей, поваренной соли (прекращая свой рост только при 15% ее концентрации).
БОЛЕЗНЕТВОРНОСТЬ (ПАТОГЕННОСТЬ), ПРОЯВЛЕНИЕ ВНЕШНИХ ПРИЗНАКОВ БОЛЕЗНИ. Пищевые отравления, причиной которых является Bac.cereus возникают при употреблении продуктов питания животного, растительного и смешанного происхождения. Bacillus cereus вызывает два типа пищевых отравлений (гастроэнтеритов); интоксикацию опосредует энтеротоксин, образуемый вегетирующими формами, прорастающими из спор устойчивых к определенным термическим режимам обработки пищевых продуктов. Бациллы образуют токсины только in vivo, во время прорастания спор.
Первый тип отличает укороченный инкубационный период (около 4-5 ч.), характерны изнуряющие диарея и рвота. Заболевание развивается при употреблении пищи, обсемененной большим количеством микроорганизмов Часты случаи отравлений в связи с употреблением жареного риса, содержащего проросшие споры Bacillus cereus, эти случаи не менее часто ошибочно связывают с отравлениями стафилококковым энтеротоксином. Подобные отравления можно считать токсикозами, связанными не столько с активностью токсина, сколько с действием метаболитов, накапливающихся в пищевых продуктах
Второй тип отравлений характеризует более продолжительный инкубационный период (около 17 ч); патогенез полностью опосредован действием энтеротоксина, больные жалуются на схваткообразные боли в животе, диарею; этот комплекс симптомов часто и ошибочно принимают за пищевые отравления, вызванные клостридиями. Патогенез обоих типов отравлении в большей или меньшей степени связан с действием энтеротоксина. В материалах семинара, состоявшегося в 1990 году, опубликована работа Ю.В. Езепчука и А.Р. Битцаева «Структурное сходство токсинов Bac.cereus и Bac.anthracis». Авторы считают, что существует структурное и функциональное сходство между диареегенным - летальным токсином (DLT) Bac.cereus и экзотоксином Bac.anthracis. DLT обладает тремя типами биологической активности: диареегенным, летальным и васкулярным, увеличивающим проницаемость. Однако механизмы действия токсина Bac.cereus остаются до конца не изученными. Считается, что патогенность B.cereus связана с ее способностью синтезировать и секретировать два экзотоксина. Один из них состоит из трех белковых компонентов, обладает, как выше указывалось, диареегенной, летальной активностью и повышает проницаемость сосудов (диареегенно-летальный токсин). Второй токсин — цереолизин — вызывает цитолитический и летальный эффект и также нарушает проницаемость кровеносных сосудов. Попадая в пищевые продукты, патогенные варианты B.cereus размножаются в них и продуцируют экзотоксины. Под влиянием протеолитических и других ферментов, выделяемых B.cereus, в продуктах накапливаются различные ядовитые вещества (птомаины). Все это вместе взятое и приводит к развитию пищевого отравления. Заражение чаще всего происходит при употреблении зараженных B.cereus растительных продуктов и молока (40-55%), а также мясных (25%) и других продуктов.
При бактериологической диагностике таких пищевых отравлений необходимо обращать внимание на количественное содержание B.cereus в продуктах (106 и более клеток в 1 г). Серологическое подтверждение диагноза дают обнаружение в сыворотке антител к B.cereus и нарастание их титра. Длительность болезни 3-6 дней. Смертельный исход редок.
ПРОФИЛАКТИКА ЗАБОЛЕВАНИЯ. В настоящее время не установлено, что у животных есть болезнь, вызываемая этим микроорганизмом. Ученые считают, что в пищевые продукты бактерии попадают из внешней среды: с кожного покрова животных, из воды, с частичками почвы, со специями. Отравления чаще всего обусловлены мясными блюдами, колбасами, консервами. В консервах Bac.cereus начинает размножаться и вырабатывать токсин при температуре от 20°С и выше. При этом на поверхности продукта появляется налет серого цвета. В сырых продуктах (мясо, мясные полуфабрикаты) быстрое размножение микроба начинается при температуре 17°С. При хранении готовой колбасы в температурном режиме холодильника (4-6°C) эти бактерии не размножаются, но при комнатной температуре (от 20°С и выше) начинается их быстрый рост с выделением токсина. На этих фактах и основывается профилактика данного заболевания. Она включает в себя соблюдение чистоты при забое животного, в первую очередь дезинфекция его кожного покрова, использование чистых инструментов при разделке туши, чистой воды, контроль условий хранения пищевых продуктов.
ОЦЕНКА ПИЩЕВЫХ ПРОДУКТОВ обсемененных данным микроорганизмом в нашей стране не разработана. Диагностическим признаком считают обнаружение в подозрительных продуктах более 105 бактерий в 1 г/мл продукта либо 102-103 бактерий в 1 г/мл каловых и рвотных масс или промывных вод. В консервах наличие Bac.сеreus как и в сырье для консервирования не допускается.
заключение
В данном издании описаны основные и наиболее распространенные в нашей стране пищевые отравления, причиной которых послужили больные животные или скрытые носители инфекционного агента. Всех их объединяет два фактора: первый - наличие общих симптомов характерных для отравления (рвота, понос, слабость, головокружение) с определенными, характерными для каждого из них особенностями; второй - причиной заболевания послужили микроорганизмы находящиеся в продуктах полученных от животных. Эти животные могут выглядеть как здоровые или переболеть достаточно давно, но являются микробоносителями. Не обязательно животные должны переболеть или болеть инфекционными заболеваниями. Возможна любая причина ослабления их организма (будь то рана или простуда). В этом случае на первую причину, в результате ослабления организма будет наслаиваться «секундарная инфекция» – «вторичная» инфекция, которая зачастую и является фактором вызывающим отравление. Поэтому при убое подобных животных и необходимо проведение ветеринарно-санитарной экспертизы с дополнительным бактериологическим исследованием полученного мяса. В зависимости от результатов микробиологических исследований, ветеринарный врач подскажет владельцу туши, что делать с мясом или продуктом, полученным из этого мяса.
Список литературы
1. Дерябкин Г.Д. Стафилококки: экология и патогенность. Екатеринбург 2000.
2. Мазохина – Поршнякова Н.Н. Подавление возбудителей ботулизма в пищевых продуках. М. Агропромиздат.1989.
3. Никифоров В.Н, Никифоров В.В Ботулизм М Медицина 1985.
4. Ленченко Е.М., Куликовский А.В, Павлова И.Б. Иерсиниоз М.1998.
5. Энтеробактерии. Под редакцией В.И. Покровского М. Медицина1985.
6. Бакулов И.А., Смирнов А.М, Васильев Д.А. Токсикоинфекции, пищевые инфекции и токсикозы микробного происхождения. М. МСХиП РФ 1995.
7. Бакулов И.А., Гаврилов В.А, Селивёрстов В.В. Сибирская язва.
8. Далин М.В, Фиш Н.Г. Белковые токсины микробов М.Медицина 1980.
9. Месорбяну Л, Пэунеску, Физиология бактерий. Бухарест. 1963.
10. Чайка Н.А., Хазенсон Л.Б., Бутцлер Ж.П. Кампилобактериоз. М. Медицина, 1988.
[КСВ1] Понижение?
Похожие работы
... для него в обычных условиях цикла развития. При перенесении токсикоинфекции имеет место бактерионосительство, в некоторых случаях довольно длительное. II. Историческая справка Впервые этиологическая роль Bacillus cereus при пищевых отравлениях была изучена и описана Наugе в 1950 г. Сначала источником пищевых отравлений, обусловленных Bac. cereus, считали кулинарные изделия, содержащие ...
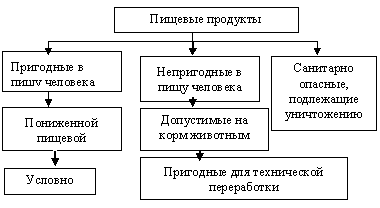
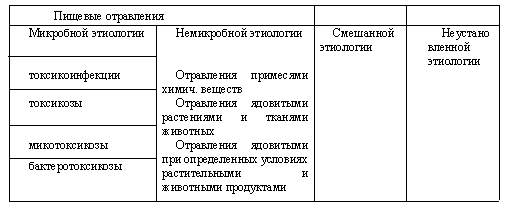
... для токсинообразования условия не всегда создаются. Этим обстоятельством и объясняется разрыв между широкой распространенностью возбудителей бактериотоксикозов в природе и относительной редкостью вызываемых ими пищевых отравлений. Профилактика бактериотоксикозов имеет некоторые особенности. Так, для предупреждения ботулизма, источниками которого могут быть все виды консервов, рыба, употребляемая ...
... заболевшими одного общего блюда. 4. Кратковременное течение заболевания (за исключением ботулизма). 5. Вспышка локализована, когда выявляется причинный пищевой продукт и исключается фактор передачи. Классификация пищевых отравлений 1. По характеру этиологического фактора: 1.1. Микробные. 1.2. Немикробные. 1.3. Неуточненной этиологии. 2. По патогенезу 2.1. Отравления микробной ...
... часть живота для вытеснения воздуха, попадающего в желудок. Искусственное дыхание не прекращают до восстановления у пострадавшего самостоятельного ритмичного и достаточно глубокого дыхания. Поражение электрическим током Действие электрического тока на организм человека имеет сложный характер, однако наиболее опасно для жизни поражение внутренних органов и, прежде всего, воздействие на ...
0 комментариев