РЕФЕРАТ
"Регуляция иммунного ответа"
Введение
Иммунный ответ, как и все биологические функции, находится под контролем разнообразных регуляторных механизмов. Эти механизмы обеспечивают восстановление исходного, «неактивного» состояния иммунной системы, когда иммунный ответ на данный антиген более не требуется. Эффективный иммунный ответ – результат взаимодействия между антигеном и целой сетью иммунокомпетентных клеток. Характер иммунного ответа, как в количественном, так и в качественном отношении, зависит от многих факторов, в том числе от типа антигена, его дозы и пути поступления, от свойств антигенпрезентирующих клеток и генетических особенностей организма, а также от предшествующего контакта иммунной системы с данным или перекрестнореагирующим антигеном. На иммунный ответ способны влиять специфические антитела.
Антиген как фактор иммунорегуляции
Активация Т- и В-клеток происходит в результате эффективного связывания антигенного материала их антигенспецифичными рецепторами.
Рецепторы Т-клеток взаимодействуют не с нативным антигеном, а с образовавшимися в результате его процессинга пептидными фрагментами, ассоциированными с молекулами МНС класса 1 или II. На результат иммунного ответа существенно влияет природа антигена, его доза и способ введения.
Тип иммунного ответа зависит от природы антигена
Различные антигены индуцируют иммунные ответы разных типов. Полисахаридные капсульные антигены бактерий обычно вызывают только гуморальный ответ, тогда как их белковые антигены – и клеточный, и гуморальный ответы. Микроорганизмы, локализующиеся внутри клеток организма-хозяина, в частности некоторые бактерии, паразиты и вирусы, индуцируют клеточный иммунный ответ, а растворимые белковые антигены – гуморальный. Клеточный иммунный ответ вызывают и такие антигены, как кремнийсодержащие соединения.
Эффективный иммунный ответ обеспечивает элиминацию антигена из организма. После этого лимфоциты возвращаются в состояние покоя. Однако некоторые антигены могут не столь эффективно удаляться из организма, что приводит к продолжению иммунного ответа в течение длительного времени с патологическими последствиями для организма.
В больших дозах антиген может индуцировать толерантность
Введение очень высокой дозы антигена нередко вызывает развитие специфической Т-клеточной, а иногда и В-клеточной толерантности. Подобный феномен часто наблюдается в случае инъекции антигена новорожденным мышам. Долгое время причиной этого считали незрелость иммунной системы. Однако теперь установлено, что у новорожденных мышей могут развиваться и полноценные иммунные реакции; отсутствие же иммунного ответа в ряде случаев связано не с незрелостью Т-клеток, а с так называемым иммунным отклонением, при котором доминирует образование непротективных цитоки-нов II типа вместо протективных цитокинов I типа. Как установлено, Т-независимые полисахаридные антигены при введении в больших дозах индуцируют толерантность В-клеток.
В зависимости от пути поступления антигена иммунный ответ может возникнуть или отсутствовать
Как установлено, немаловажное значение для возникновения иммунного ответа имеет способ введения антигена. Антигены, введенные подкожно или внутрикожно, вызывают иммунный ответ, тогда как при внутривенной инъекции, приеме внутрь или применении в виде аэрозоля они могут индуцировать толерантность либо иммунное отклонение. Например, грызуны в случае приема овальбумина или основного белка миелина с кормом не реагируют на последующую стимуляцию соответствующим антигеном. Более того, применение ОБМ защищает животных от развития аутоиммунного заболевания – экспериментального аллергического энцефаломиелита. Этот феномен может быть использован с терапевтической целью при аллергических расстройствах; недавно проведенные исследования показали, что пероральное введение Т-клеточного эпитопа аллергена Der pi клеща домашней пыли может обеспечить толерантность к нативному антигену. Мезанизмом толерантности при этом может быть как анергия, так и иммунное отклонение.
Подобные наблюдения были сделаны и при использовании антигенов в форме аэрозолей. Эксперименты, проведенные на мышах, показали, что введение энцефалитогенного пептида интраназально в виде аэрозоля снижает интенсивность развития ЭАЭ, который возникает при последующем обычном способе введения пептида. Этот факт также может иметь значение для разработки методов лечебного воздействия, поскольку ингибировать ответ способен не только данный антиген, применяемый в виде аэрозоля, но и другие антигены, вызывающие ЭАЭ.
Наглядный пример того, как может влиять на иммунный ответ способ введения антигена, дало изучение инфекции, вызываемой у мыши вирусом лимфоцитарного хориоменингита. У мышей, примированных пептидом в неполном адъюванте Фрейнда путем его подкожного введения, развивается иммунитет к ВЛХМ. Однако, если тот же пептид введен внутрибрюшинно, животные становятся толерантными и теряют способность элиминировать вирус.
Антигенпрезентирующие клетки
Природа АПК, осуществляющих первоначальное представление антигена, может определять тип вызываемой им реакции – полноценный иммунный ответ или толерантность. Для эффективной активации Т-клеток необходимо присутствие на поверхности АПК костимулирующих молекул. Поэтому презентация антигена дендритными клетками или активированными макрофагами, которые экспрессируют в большом количестве антигены МНС класса II и наряду с ними костимулирующие молекулы, ведет к высокоэффективной активации Т-клеток. Кроме того, взаимодействие молекул CD40L, экспрессируемых на поверхности активированных Т-лимфоцитов, и CD40 на поверхности дендритных клеток обеспечивает интенсивную продукцию этими последними ИЛ-12, необходимого для эффективного Txl-ответа. Если же антиген презентируют Т-клеткам «непрофессиональные» АПК, которые неспособны обеспечить костимуляцию, возникает ареактив-ность или иммунное отклонение. Так, представление антигена нестимулированным Т-клеткам покоящимися В-лимфоцитами вызывает не активацию, а толерантность Т-клеток. Адъюванты могут способствовать развитию иммунного ответа тем, что они индуцируют экспрессию антигенов МНС и костимулирующих молекул с большой плотностью на поверхности АПК. Иллюстрацией этого служат результаты недавно проведенных экспериментов по изучению механизмов толерантности у новорожденных животных как более чувствительных к индукции толерантности, чем взрослые. Эти исследования показали, что резистентность к ЭАЭ, вызываемая введением ОБМ в неполном адъюванте Фрейнда, связана с развитием доминантного Тх2-ответа. В возникновении ЭАЭ участвуют Txl-клетки, а предшествующий Тх2-ответ на ОБМ предотвращает патологический Txl-ответ.
Значение дендритных клеток в индукции ответа, опосредуемого цитотоксическими Т-лимфоцитами, установлено в экспериментах с переносом новорожденным мышам-самкам клеток от мышей-самцов. Самки, получившие спленоциты, не продуцировали Тц-ответ на последующее введение З-Х-антигена мышей-самцов. В то же время перенос дендритных клеток обеспечивал развитие полноценного, З-Х-специфичного Тц-ответа.
Регуляторное влияние антител
Как установлено, антитела осуществляют регуляцию иммунного ответа по механизму обратной связи. Пассивно введенные вместе с антигеном IgM-антитела специфически усиливают иммунный ответ на данный антиген, тогда как IgG-антитела его подавляют. Первоначально это было выявлено на модели пассивной иммунизации поликлональными антителами, а затем получило подтверждение в экспериментах с использованием моноклональных антител.
Способность пассивно введенных антител усиливать или подавлять иммунный ответ учитывают при вакцинации и используют в клинической практике.
• Иммунизацию некоторыми вакцинами проводят обычно детям старше одного года, поскольку в течение по крайней мере 6 мес после рождения в крови ребенка имеется большое количество IgG-антител, полученных от матери, а присутствие таких пассивно приобретенных антител во время вакцинации может существенно снизить ее эффективность.
• В случаях резус-несовместимости введение резус-отрицательной матери антител ан-ти-RhD предотвращает первичную сенсибилизацию Кп+-эритроцитами плода, возможно в результате элиминации чужеродного антигена из крови матери.
Механизмы модуляции иммунного ответа под влиянием антител еще недостаточно полно выяснены. Предполагается, что повышение продукции бляшкообразующих клеток при действии lgM-антител может быть обусловлено двумя факторами.
• Содержащие IgM иммунные комплексы поглощаются с участием Fc-рецепторов или СЗ-рецепторов на поверхности АПК и процессируются более эффективно, чем свободный антиген.
• Содержащие IgM иммунные комплексы стимулируют образование антиидиотипических антител против IgM, которые усиливают иммунный ответ.
lgG-антитела могут подавлять синтез специфических IgG
Опосредованная IgG супрессия может осуществляться разными путями.
Блокирующее действие антител Пассивно введенные антитела связывают антиген, конкурируя с В-клетками. В этом случае эффект IgG-антител существенно зависит от их концент-
рации, а также от соотношения их аффинности к антигену с аффинностью В-клеточных рецепторов. Успешно конкурируют с антителами за антиген только те В-клетки, которые обладают высокоаффинными рецепторами, причем механизм конкуренции не зависит от Fc-фрагмента антител.
Перекрестное связывание рецепторов Антитела IgG также оказывают регуляторное действие; оно обусловлено Fc-фрагментом их молекулы. Экспериментально установлено, что иммуноглобулин способен ингибировать дифференцировку В-клеток путем перекрестного связывания антигенного рецептора с Fc-рецептором на поверхности той же клетки. В этом случае антитела могут распознавать различные эпитопы.
В дозах, недостаточных для полного подавления продукции антител, IgG повышает их среднюю аффинность в результате того, что успешно конкурировать с пассивно введенными антителами за антиген способны лишь В-клетки, обладающие высокоаффинными рецепторами. Как предполагается, регуляция по механизму обратной связи, осуществляемая антителами, играет важную роль в процессе повышения аффинности антител.
Иммунные комплексы могут усиливать или подавлять иммунные реакции
Один из механизмов модулирующего влияния антител на иммунный ответ является Fc-зависимым и связан с образованием иммунных комплексов антиген–антитело. Иммунные комплексы могут ингибировать или усиливать иммунный ответ. Активируя комплемент, иммунные комплексы могут локализоваться путем взаимодействия с CR2 на фолликулярных дендритных клетках. Это способствует иммунному ответу, поскольку обеспечивает постоянный источник антигена. Рецептор CR2 экс-прессируется также на В-клетках, и при этом известно, что косвязывание CR2 с мембранным IgM активирует В-клетки; таким образом, взаимодействие иммунных комплексов с CR2, входящим в состав В-клеточного корецеп-торного комплекса, и mlg может приводить к усилению специфического иммунного ответа.
У больных со злокачественными опухолями иммунореактивность часто бывает подавлена; предполагается, что это связано с присутствием в крови иммунных комплексов, состоящих из антител и антигенов опухолевых клеток.
Роль лимфоцитов в иммунорегуляци
Т-лимфоциты оказывают очевидное положительное влияние на иммунный ответ, выполняя хелперную функцию. Кроме того, в зависимости от типа хелперных Т-клеток иммунный ответ может носить характер гуморального или клеточного. Имеется также убедительное доказательство того, что Т-клетки способны подавлять иммунный ответ.
Т-клетки CD4 могут предотвращать возникновение аутоиммунитета
При многих экспериментальных аутоиммунных, заболеваниях Т-клетки CD4+, образующиеся после введения высоких доз аутоантигенов, предотвращают последующую индукцию аутоиммунного процесса. Так, Т-клетки CD4+ предотвращают образование аутоантител к тиреоглобулину.
Установлено также, что одновременное введение мышам антител анти-С04 и иммуногенной дозы тиреоглобулина не только предотвращает развитие аутоиммунитета, но приводит к образованию по– Механизм, посредством которого Т-клетки осуществляют такого рода отрицательное влияние, не вполне ясен. Однако проведенные недавно эксперименты указывают на то, что частично или полностью подавлять иммунный ответ могут продуцируемые Тх-клетками цитокины ФЦСв, ИЛ-4и ИЛ-10.
В регуляции антителообразования участвуют различные субпопуляции Тх-клеток
Некоторые наблюдения, касающиеся регуляции синтеза IgE-антител, можно объяснить тем, что Тх-клетки CD4+ разных субпопуляций продуцируют разные цитокины. Между отдельными субпопуляциями Тх-клеток существует взаимосвязь в виде перекрестной регуляции их активности: секретируемый Txl-клетками ИФу способен ин-гибировать реактивность Тх2-лимфоцитов; ИЛ-10, образуемый Тх2-клетками, понижает экспрессию молекул В7 и ИЛ-12 антигенпрезенти-руюшими клетками, что в свою очередь ингибирует активацию Txl-лимфоцитов. Кроме того, на экспрессию высокоаффинного рецептора ИЛ-12, необходимого для развития Тх1-кле-ток, влияет баланс цитокинов. Высокоаффинный ИЛ-12Р содержит в своей структуре две цепи, вА и в2, причем обе они вместе экспрессируются только клетками Txl. Цепь в 1 имеют и Txl-, и Тх2-лимфоциты, а экспрессию в2-херр индуцирует ИФуи ингибирует ИЛ-4. На развитие субпопуляций Т-клеток оказывает также влияние ИФа, который благоприятствует формированию субпопуляций Txl-лимфоцитов даже в присутствии ИЛ-4 и в условиях нейтрализации ИЛ-12. Таким образом, преимущественная активация Txl- или Тх2-клеток может приводить к иммунному отклонению – избирательному развитию эффекторного ответа определенного типа. Такой избирательный сдвиг ответа может быть использован в терапии аутоиммунных и аллергических заболеваний.
Адоптивным переносом Т-клеток CD8+ можно вызвать резистентность и толерантность
Установлено, что Т-клетки CD8+ также регулируют иммунный ответ. В селезенке животных, у которых индуцировали толерантность к ОБМ путем перорального введения антигена, были обнаружены Т-лимфоциты CD8+, способные при адоптивном переносе создавать у реципиентов резистентность к ЭАЭ. Эти Т-клетки не только супрессируют Т-клеточный ответ на ОБМ in vitro, но могут также вызывать эффект «подавления свидетеля» в отношении других, неродственных антигенов. Предположительно данный эффект опосредован ФЦСв.
Регуляция иммунного ответа Тх2-клетками CD4+ представляет собой нормальный физиологический процесс
Роль регуляторных эффектов, опосредованных Т-клетками CD4+ или CD8+, в обычных физиологических условиях остается под вопросом. Однако обнаружение в норме у животных Т-лимфоцитов CD4+, способных предупреждать развитие аутоиммунитета, свидетельствует об их значении в поддержании нормального гомеостаза. Кроме того, для крысы и мыши установлено, что после удаления Тх2-клеток CD4+, продуцирующих в обычных условиях ИЛ-4 и ИЛ-10, нарушается регуляция иммунного ответа. Этот факт убедительно доказывает, что регуляция иммунного ответа, осуществляемая Тх2-лимфоцитами CD4, представляет собой нормальный физиологический процесс, а не артефакт. По имеющимся данным, при возникновении колита у мышей регулирующую роль в развитии патологического процесса может играть ФЦСв. Это согласуется с результатами опытов по изучению колита у мышей, лишенных гена ФЦСв методом генного нокаута.
Идиотипическая модуляция иммунного ответа
Толерантность к собственным антигенам развивается в онтогенезе. Однако в неонанальный период уникальные связывающие участки антигенспецифичных рецепторов на В- и Т-клетках слишком малочисленны, чтобы индуцировать толерантность. То же самое относится к антителам: хотя они присутствуют в сыворотке крови, толерантность развивается только к их Fc-фрагментам, поскольку лишь они имеются в достаточной концентрации; толерантности к детерминантам легких и тяжелых цепей, ответственным за связывание специфических антигенов, не возникает. Таким образом, индивидуальные Т-клеточные рецепторы и иммуноглобулины являются иммуногенными за счет этих уникальных последовательностей, называемых илиотипами. Антитела, образующиеся против антигенсвязывающих центров, способны влиять на результат иммунного ответа.
Идиотипические детерминанты могут кодироваться гаметными генами V-областей или возникать в результате рекомбинаций и мутаций, обусловливающих образование функционально активных генов V-областей иммуноглобулинов. Иммуногенные эпитопы в самом связывающем центре или расположенные вокруг него называют идиотопами. Известным иммунологом Йерне выдвинута гипотеза о существовании в организме иммунной сети, внутри которой взаимодействия осуществляются путем распознавания идиотипов. Согласно этим представлениям, образующиеся к антигену антитела в свою очередь вызывают направленный против них антиидиотипический ответ. Эта гипотеза концептуально привлекательна, однако роль такого рола идиотипической сети в регуляции нормального иммунного ответа остается предметом горячих споров.
Идиотипические взаимодействия могут усиливать или подавлять гуморальный иммунный ответ
Получены убедительные доказательства в пользу того, что антиидиотипические антитела могут влиять на воспроизведение распознаваемых идиотипов при иммунном ответе. Так, если мышам линии C57BL/6 ввести гаптен нитрофенол, они продуцируют в основном антитела немногих определенных идиотипов, например идиотипа 146. Антиидиотипические антитела против этих антител могут усиливать или подавлять образование идиотипа 146 в ответ на последующую инъекцию З Ц вместе с белком-носителем. Наблюдаемый эффект зависит от количества вводимых антиидиотипических антител и является идиотипспецифичным, поскольку общий уровень антител против НФ изменяется мало. Наиболее важно отметить, что применявшиеся концентрации антиидиотипов не выходят за пределы нормальных физиологических значений для антител, несущих данный идиотип, и это позволяет предполагать, что илиотипическая регуляция может осуществляться in vivo. Подобные наблюдения сделаны и при исследовании других идиотипических систем.
Драматические последствия имеет введение антиидиотипических антител новорожденным животным, когда возникающий эффект может сохраняться пожизненно. Так, способность мышей отвечать на фосфорилхолин резко снижается, если в неонатальный период животные получали антиидиотипические антитела к Т15. Состояние сниженной реактивности сохраняется в течение многих месяцев. У таких мышей при последующем введении специфического антигена доминирует продукция иммуноглобулинов, не имеющих идиотипа Т15.
Нейроэндокринная регуляция иммунного ответа
Уже давно известно, что стрессовые ситуации могут служить причиной подавления иммунных функций организма, например снижения его способности преодолевать инфекции. Имеются многочисленные данные, указывающие на взаимодействие между нервной, эндокринной и иммунной системами. В общем виде два основных пути, посредством которых процессы, происходящие в центральной нервной системе, могут отражаться на иммунной функции, состоят в следующем.
• Большая часть лимфоидных тканей имеет прямую симпатическую иннервацию – как кровеносных сосудов, проходящих через лимфоидную ткань, так и непосредственно самих лимфоцитов.
• Нервная система прямо или опосредованно контролирует секрецию различных гормонов, в частности кортикостероидов, гормона роста, тироксина и адреналина.
Лимфоциты экспрессируют рецепторы для многих гормонов, медиаторов и нейропептидов, включая рецепторы для стероидов, катехолами-нов, энкефалинов, эндорфинов, вещества С и вазоактивного интестинального пептида. Степень экспрессии рецепторов и клеточная реактивность варьируют у различных популяций лимфоцитов и моноцитов, в связи с чем эффект разных медиаторов также варьирует в зависимости от условий. Однако применительно к иммунной системе особое значение имеет регуляция, опосредованная кортикостероидами, эндорфинами и энкефалинами – агентами, которые высвобождаются при стрессе и обладают иммуносупрессивным действием in vivo. Эффекты эндорфинов in vitro существенно различаются в зависимости от экспериментальной системы и дозы; в одних дозах они оказывают супрессивное влияние, в других – усиливают иммунный ответ. Однако одним из важных факторов, регулирующих иммунный ответ по механизму обратной связи, служат, несомненно, кортикостероиды. Установлено, что сами лимфоциты способны реагировать на кортико-тропин-рилизинг-гормон, синтезируя собственный АКТГ, который в свою очередь индуцирует секрецию кортикостероидов.
По имеющимся данным, кортикостероиды ингибируют продукцию цитокинов Txl-клетками, не влияя на Тх2-ответ. Кроме того, они индуцируют образование ФЦСв, который может подавлять иммунный ответ. Предполагается, что низким уровнем кортикостероидов в плазме у крыс линии Lewis обусловлена повышенная предрасположенность этих животных к возникновению различных аутоиммунных процессов: после индукции ЭАЭ спонтанное выздоровление крыс связано с повышением содержания в крови кортикостероидов; у адреналэктомированных животных выздоровления не происходит. Значение стероидов в предрасположенности к заболеванию продемонстрировано также на крысах линии PVG: в норме животные этой линии резистентны к ЭАЭ, однако становятся чувствительными к нему после адреналэктомии.
Взаимодействие между нейроэндокринной и иммунной системами не является однонаправленным. Установлено, что цитокины, в частности ИЛ-1 и ИЛ-6, действуют в обоих направлениях, играя роль модуляторов взаимодействия этих двух систем. Данные цитокины служат мощными стимуляторами продукции кортикостероидов надпочечниками благодаря своему влиянию на кортикотропин-рилизинг-гормон. Помимо того, что ИЛ-1 продуцируют макрофаги, а ИЛ-6 – Т-клетки, способностью к синтезу обоих этих цитокинов обладают нейроны и клетки глии, а также клетки, локализованные в гипофизе и надпочечниках. Это еще раз подчеркивает важную роль данных цитокинов как медиаторов двунаправленного действия при реакции организма на стресс.
Генетическая регуляция иммунного ответа
Хорошо известно, что разные индивиды неодинаково реагируют на один и тот же антиген. Результаты семейного анализа чувствительности к
Corynebacteriutn diphtheriae позволили предположить, что резистентность или чувствительность к инфекции может быть наследуемым признаком. В пользу этого свидетельствуют и данные, полученные на морских свинках разных линий, неодинаково резистентных к дифтерии; различия были определены как генетически детерминированные. В 1943 г. Фьёрд-Шайбель в исследованиях по селекции линий морских свинок с высокой и низкой иммунореактивностью продемонстрировал, что продукцию дифтерийного антитоксина контролирует один ген и она наследуется как доминантный менделевский признак. В этом исследовании впервые был установлен также доминантный тип наследования высокой иммунореактивности. В потомстве первого поколения, полученном от скрещивания животных с высокой иммунореактивностью, 90% особей продуцировали антитоксин, тогда как при скрещивании низкореагирующих свинок лишь в пятом поколении удалось получить 90% животных с низкой иммунореактивностью.
Способность отвечать на антиген зависит от гаплотипа МНС
Получение инбредных, или чистых, линий мышей позволило более глубоко исследовать влияние генетических факторов и окончательно доказать их роль в иммунологической реактивности. Мыши с различными гаплотипами МНС различаются по способности к гуморальному ответу на специфические антигены. Эта функция зависит от МНС-молекул класса II и специфична для каждого антигена: линия с высоким уровнем ответа на один антиген может на другие отвечать слабо. Гены МНС, как было установлено, выполняют наиболее важную роль в регуляции ответа на инфекционные агенты.
Не сцепленные с МНС гены также влияют на иммунный ответ
Достигнутые в последние годы в иммунологии значительные успехи – выяснение структуры молекул МНС классов I и II, изучение полиморфизма аминокислотных последовательностей молекул МНС и его значения для связывания пептидов, разработка молекулярных методов контроля за формированием репертуара Т-клеточных рецепторов и технологии получения трансгенных мышей – внесли вклад в выяснение того, каким образом генетические факторы влияют на иммунный ответ. К числу этих факторов относятся не только гены, сцепленные с МНС. Например, тяжелый комбинированный иммунодефицит обусловлен отсутствием гена рекомбиназы, а недостаточность адгезии лейкоцитов возникает вследствие мутаций гена, кодирующего субъединицу р2-интегрина, и вызванного ими нарушения экспрессии LFA-1, CR3 и CR4.
Иммунный ответ зависит от активации клонов лимфоцитов. Т-клетки распознают антиген только в том случае, если он презентирован им в виде комплекса антигенного пептида с антигенами МНС классов I или II. Поэтому, например, мышиные Тц-клет-ки CD8+, специфичные к гликопротеину ВЛХМ, способны лизировать лишь такие инфицированные вирусом клетки-мишени, которые получены от мышей, совместимых по МНС-антигенам класса I с донорами клеток-эффекторов. Способность к такому распознаванию развивается в онтогенезе.
Формирование репертуара периферических Т-лимфоцитов зависит как от разнообразных ау-тоантигенов, так и от способности Т-клеток связываться с собственными антигенами МНС. Способность пептида к образованию комплекса с молекулой МНС определяется последовательностью аминокислотных остатков ее пептилсвязывающего участка. Теперь мы знаем, что большая часть полиморфных аминокислотных остатков молекулы МНС локализована в ее пептидсвязывающей полости. Поэтому высокий полиморфизм аминокислотных последовательностей МНС-антигенов очень важен для связывания пептидов и вследствие этого для активации Т-клеток. Как установлено, при развитии в тимусе Т-клетки подвергаются селекции двух типов.
• Положительная селекция основана на взаимодействии ТкР с молекулами МНС на поверхности эпителиальных клеток корковой зоны тимуса.
• Отрицательная селекция происходит в результате высокоаффинного взаимодействия между ТкР и комплексом МНС–пептид, презентированным на клетках костномозгового происхождения в мозговой зоне тимуса.
пользовать этих животных для демонстрации положительной селекции. Данный ТкР идентифицируют при помощи антител, специфичных к нв8.
Различные молекулы МНС по-разному влияют на образование зрелых Т-клеток CD8+, экспрессирующих трансгенный ТкР. Положительная селекция Т-клеток CD8+ с трансгенным ТкР, содержащим \ф8-цепь, была зарегистрирована лишь у тех мышей, которые экспрессировали молекулу H-2Db. Таким образом, положительная селекция данного рецептора происходит только при экспрессии соответствующе-
Положительная селекция в тимусе означает отбор Т-клеток по способности распознавать антигены, презентированные в комплексе с собственными молекулами МНС
Первые исследования в этой области проводились по сложной схеме, включавшей тимэктомию, облучение, введение костного мозга и пересадку тимуса. Их результаты показали, что цитотоксические лимфоциты лизируют только клетки-мишени с такими же МНС-антигенами, какие экспрессируют клетки тимуса, в котором происходило развитие Т-клеток. Эти и другие данные позволяют предполагать, что созревающие Т-клетки «учатся» распознавать антигены только в контексте молекул МНС, с которыми они первоначально контактировали в тимусе.
Этот процесс прояснили эксперименты на трансгенных мышах, когда стало возможным получать таких трансгенных животных, у которых основная часть Т-клеток несет всего один антигенспецифичный рецептор. Соответствующий антиген также может экспрессироваться как продукт трансгена. Это значительно упрощает анализ, поскольку исследуемые Т-клетки составляют большую часть Т-клеточной популяции и их можно определить при помощи клонотипических, или нв-специфичных, антител. Так, можно получить трансгенных мышей, у которых Т-клетки в основном экспрессируют ТкР клона Тц CD8+, распознающий гликопротеин ВЛХМ в комплексе с молекулой H-2Db класса I, и исго МНС-гаплотипа. Подобный отбор протекает на поверхности эпителиальных клеток корковой зоны тимуса. Предполагается, что пептидами, опосредующими положительную селекцию, служат в норме присутствующие в тимусе аутопептиды.
Т-клетки, распознающие аутоантигены, подвергаются отрицательной селекции
Отрицательная селекция путем клональной делеции продемонстрирована с использованием моноклональных антител, специфичных к нв-це-пям мышиных ТкР. Таким способом были идентифицированы и подсчитаны Т-клетки, несущие ТкР с данной нв-цепью, и этот анализ показал, что у мышей, экспрессирующих 1-Е, Т-клетки \ф17а+ делетируются в тимусе. Присутствие Т-клеток Y4317a+CD4+CD8+, но не зрелых Т-клеток нв173+004+ или vpi7a+CD8.1+, позволило предположить, что делеция происходит на стадии дважды положительных Т-клеток в процессе их созревания. При этом было установлено, что для делеции необходима экспрессия наряду с 1-Е и эндогенного лиганда, а не только одного 1-Е.
Суперантигены могут вызывать полную делецию отдельных серий Т-клеточных рецепторов
У мышей ряда линий в тимусе происходит делеция целых популяций Т-клеток, несущих определенные ТкР. Так, у мышей, экспрессирующих минорный, стимулирующий лимфоциты антиген Mls-la и некоторые МНС-молекулы класса II, делетируются Т-клетки нв6+ и нв8.1+. Способностью целых семейств Т-лимфоцитов распознавать антигены Mis объясняется интенсивный пролиферативный ответ, наблюдаемый при совместном культивировании несовместимых по Mis клеток. В ответе на некоторые антигены участвуют все Т-клетки, экспрессирующие определенные нв-цепи.
Вызывающие такой сильный ответ антигены, как экзогенные, так и эндогенные, названы суперантигенами. Они связываются не в пептид-связывающей полости антигенов МНС, а непосредственно с молекулами МНС класса I и II и в-цепями ТкР. Примером экзогенного суперантигена может служить стафилококковый энтеротоксин В. Pea-
гирующие на этот антиген Т-клетки мышей имеют ТкР, содержащий нв3- или \ф8-цепи. Mis-антигены относятся к классу эндогенных суперантигенов. Однако к настоящему времени установлено, что наличие Mis-антигенов в действительности определяется присутствием эндогенных вирусов опухоли молочной железы мыши, кодируемых длинным З'-терминальным повтором в геноме мышей данной линии. Существует множество различных М MTV, и мыши каждой линии имеют в геноме лишь некоторые из них. Эти эндогенные суперантигены, экспрессированные в тимусе, вызывают делецию Т-клеток, несущих ТкР с цепью нв3, нв6 или нв8.1. Для изучения того, каким образом селекция в тимусе на ранних стадиях развития организма может повлиять на способность взрослого животного формировать ответ, были использованы также трансгенные мыши, описанные в предыдущем разделе. При инфицировании ВЛХМ в неонаталь-ный период у мышей развивалась толерантность и они становились носителями вируса. Анализ созревающих Т-клеточных популяций у этих неона-тально инфицированных мышей показал значительное снижение числа Т-клеток CD4+CD8+ – возможное последствие клональной делеции Т-клеток на ранней стадии онтогенеза. Таким образом, формирование репертуара периферических Т-лимфоцитов происходит в результате как положительной, так и отрицательной селекции. и соответственно оба эти типа отбора влияют на иммунореактивность.
Сцепленные с МНС гены регулируют ответ на инфекционные агенты
Определенную роль в развитии ответа на инфекционные агенты, а также на аутоантигены выполняют, как установлено, гены, сцепленные с МНС. В некоторых случаях эту функцию несут гены, входящие в МНС.
Чувствительность мышей к инвазии Trichinella spiralis определяется локусом 1-Е
Первое наблюдение, свидетельствующее о том, что гены, сцепленные с МНС, могут влиять на иммунный ответ против паразитарной инвазии, было сделано на модели трихинеллеза, вызываемого Trichinella spiralis. При заражении Trichinella spiralis мышей ряда рекомбинантных линий обнаружено, что резистентность или восприимчивость животных зависит от локуса 1-Е: мыши, экспрессируюшие 1-Е, восприимчивы к инвазии. На ответ влияет также один сцепленный с МНС ген. Он не входит в состав МНС, а находится в неравновесии по сцеплению с МНС. Этот ген, названный Ts-2, картирован вблизи гена ФНО.
Локус 1-Е влияет также на чувствительность к Leishmania donovani
Субрайон 1-Е влияет также на чувствительность мышей к Leishmania donovani. В исследованиях на мышах, конгенных по комплексу Н-2, установлено, что экспрессирующие 1-Е мыши не обладают резистентностью к висцеральной форме лейшманиоза. Прямое участие продукта 1-Е в определении чувствительности к данной инвазии обнаружено по действию антител анти-Й–Е, которые усиливали элиминацию паразитов из организма. Кроме того, перенос трансгена 1-Е мышам, лишенным этого локуса, делал животных неспособными уничтожать лейшманий, локализующихся в печени и селезенке, как это происходит у мышей исходной линии.
Некоторые HLA-гаплотипы обеспечивают защиту против Plasmodium falciparum
Изучение ассоциации HLA-гаплотипов с тяжелой формой анемии у больных малярией, показало, что распространенный среди населения Западной Африки и редко встречающийся в других расовых группах гаплотип DRB1*1302-DQB1*0501 обеспечивает защиту от летального исхода при малярии, вызываемой Plasmodium falciparum. Установлено, что молекулы DRBP1302 и DRB1*1301, различающиеся одним аминокислотным остатком в в-цепи, связывают разные пептиды и это различие, очевидно, влияет на иммунный ответ против этого возбудителя малярии.
Сцепленные с МНС гены существенно влияют на чувствительность к аутоиммунным заболеваниям Ассоциации с МНС-генами
Инсулин-зависимый сахарный диабет – аутоиммунное заболевание, при котором в-клет-ки поджелудочной железы разрушаются клетками иммунной системы, – ассоциирован с антигенами HLA-DR3 и HLA-DR4. Наибольший риск развития заболевания существует в случае гете-розиготности HLA-DR3/DR4. В действительности ИЗСД ассоциирован с генами DQ, хотя вследствие неравновесия по сцеплению первоначально была описана ассоциация с DR. С помощью молекулярно-генетических методов эту ассоциацию удалось изучить более детально, и оказалось, что у европеоидов заболевание первично ассоциировано с геном DQB 1*0302. Ревматоидный артрит в разных этнических группах ассоциирован главным образом с HLA-DR4 или HLA-DR1 и лишь в небольшом числе случаев с HLA-DQ. Механизмы, лежащие в основе повышенной чувствительности к указанным заболеваниям, пока неясны. Одной из возможных причин ее связи с гаплотипом может быть различие в репер-туарах генов класса II, возникающее как результат положительной и отрицательной селекции. Не исключено также, что вызывающие иммунный ответ эпитопы на поверхности бактерий или вирусов связываются преимущественно с теми или иными молекулами МНС. Подтверждением этой гипотезы могут служить данные, полученные при анализе аминокислотных последовательностей, образующих пептидсвязывающие полости молекул HLA-DR4 и HLA-DR1: ассоциированные с чувствительностью или резистентностью субтипы HLA-DR содержат различно заряженные аминокислотные остатки.
Ассоциации с МНС-областью
Другим примером неравновесия по сцеплению служит ассоциация аутоиммунного процесса у мышей F1 с гаплотипом H-2Z родительской линии NZW. Четко установлено, что заболевание ассоциировано не с самими МНС-генами, а с геном ФНОа, тесно сцепленным с МНС. Аллель ФНОа у мышей линии NZW детерминирует низкий уровень продукции ФНОа. Если концентрация цитокина возрастает, животные приобретают устойчивость к волчаночному нефриту.
Ассоциации с генами, регулирующими процессинг антигенов
Недавно были идентифицированы и другие сцепленные с МНС гены, способные влиять на иммунный ответ. Эти гены имеют отношение к протеолитическому расщеплению антигенов с образованием антигенных фрагментов и к их транспорту. Данные гены отличаются полиморфизмом, имеющим функциональное значение. Так, например, у крысы различные аллельные формы локуса cim определяют эффективность образования комплексов молекул МНС класса I с пептидами, а она в свою очередь влияет на возможность распознавания МНС-молекул этого класса как аллоантигенов. Таким образом, возможно, что установленные ассоциации некоторых заболеваний с МНС обусловлены генами подобного типа, регулирующими протеолиз антигенов и транспорт антигенных пептидов к молекулам МНС, которые презентируют их клеткам иммунной системы.
Многие не-МНС-гены также модулируют иммунные ответы
Иммунный ответ регулируют также некоторые гены, расположенные вне области МНС. Однако в целом эти гены отличаются меньшим полиморфизмом по сравнению с МНС, в связи с чем их вклад в определение чувствительности к заболеваниям не столь велик, как генов МНС. Тем не менее не-МНС-гены могут играть определенную роль при аутоиммунных болезнях, аллергии и
• повышение экспрессии МНС-антигенов класса II.
Недавно проведенные исследования позволили идентифицировать ген Nramp, потенциально способный функционировать в качестве гена Beg у мыши. Поскольку Nramp кодирует мембранный белок, гомологичный известным транспортным белкам, было высказано предположение, что его экспрессия может влиять на транспорт NO2 в фаголизосомы и тем самым на уничтожение внутриклеточных микроорганизмов. Однако пока экспериментально не доказано, что этот ген кодирует продукт Beg, данное предположение остается чисто гипотетическим.
Путем направленного отбора получены две линии мышей Biozzi, различающиеся уровнем реактивности после иммунизации чужеродными эритроцитами. Одним из факторов, определяющих неодинаковую способность мышей этих линий отвечать на эритроцитарные антигены, служат генетически детерминированные различия в активности макрофагов. Иммунные реакции на паразитарные инвазии у мышей высоко- и низкоотвечающих линий также существенно различаются, и эти различия не обязательно коррелируют с количеством продуцируемых антител к эритроцитам барана.
Эозинофилы играют важную роль в реакции организма на паразитарные инвазии. Установлено, что степень эозинофилии, развивающейся при инвазии, генетически детерминирована, и у мы-
шей разных линий этот показатель значительно варьирует. Подобное же явление наблюдается у морской свинки и овны: найдена выраженная корреляция между резистентностью к инвазии, вызываемой нематодами, и интенсивностью эо-зинофилии.
Не сцепленные с МНС гены влияют на развитие аутоиммунных заболеваний
Крупным успехом в изучении генетических основ аутоиммунной патологии стало проведенное недавно картирование локусов, регулирующих предрасположенность к инсулин-зависимому сахарному диабету. Работа была проведена в основном на мышах линии NOD, у которых спонтанно развивается аутоиммунное заболевание, сходное с ИЗСД человека. У этих мышей картированы по меньшей мере 15 генетических локусов, и только один из них оказался сцепленным с МНС в хромосоме 17. Предполагается, что этот ген непосредственно кодирует молекулы МНС класса П. Другие гены картированы в разных хромосомах, однако их природа и роль в резистентности или предрасположенности к заболеванию пока неизвестны.
У мышей, несущих встроенный ген Ipr, возникает лимфопролиферативное заболевание с характерными клиническими признаками. Животные продуцируют антитела против ДНК и ревматоидный фактор; у них образуются циркулирующие иммунные комплексы и развивается гломеруло-нефрит. Наблюдается также лимфаденопатия с экспансией Т-клеток CD4~CD8~ на периферии. Эти Т-клетки не являются моноклональными, но имеют различную структуру ТкР. Первоначально для объяснения причин этого синдрома было высказано предположение о дефекте отрицательной селекции, однако трудно было представить, каким образом ген, подобный Ipr, может опосредовать такой эффект. Позднее выяснилось, что у мышей, несуших ген Ipr, имеется дефект молекулы CD95, или Fas, ген которой расположен в хромосоме 19. Fas представляет собой трансмембранную молекулу, принадлежащую к суперсемейству рецепторов ФНО; взаимодействуя со своим лигандом CD95L, или FasL, она индуцирует запрограммированную клеточную гибель – апоптоз. Молекула CD95L входит в семейство факторов некроза опухолей и роста нервов. Дефект Fas-антигена, возникающий вследствие мутации Ipr, приводит к отсутствию апоптоза. Однако этот дефект, по-видимому, не затрагивает процесс отрицательной селекции и формирование нормального репертуара зрелых моноположительных Т-клеток в тимусе. Очевидно, Fas служит лишь одним из лигандов, опосредующих апоптоз. В настоящее время предполагается, что этот дефект ведет к экспансии дважды отрицательных Т-клеток на периферии и ускорению аутоиммунного синдрома. Нарушение апоптоза происходит также у В-клеток, приводя к накоплению периферических аутореактивных В-лимфоцитов.
В других исследованиях было установлено, что ген gld, присутствие которого обусловливает возникновение аутоиммунного заболевания, сходного с наблюдаемым у мышей Ipr/lpr, кодирует дефектный FasL. Таким образом, у мышей gld/gld, т.е. не экспрессирующих функциональный лиганд, также наблюдается недостаточность апоптоза периферических В- и Т-клеток и развитие аутоиммунитета. Ген gWлокализован у мыши в хромосоме 1 и, таким образом, служит примером гена, влияющего на иммунную функцию, но не сцепленного с МНС. Синдромы, сходные с описанными выше, встречаются у человека; они также связаны с дефектами активности и функции Fas.
Похожие работы
... иммунной системы, влияя не только через гормоны соответствующих периферических эндокринных желез, но и прямо на эти клетки. Влияние гормонов гипофиза на иммунную систему было рассмотрено выше в разделе "Нейропептиды и регуляция иммунного ответа". Схема основных путей взаимодействия нейроэндокринной и иммунной систем в целостном организме. Антиген вызывает активацию антиген-чувствительных кл
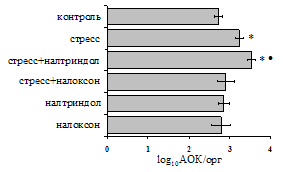
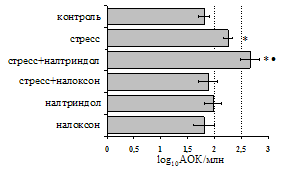
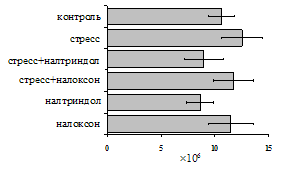
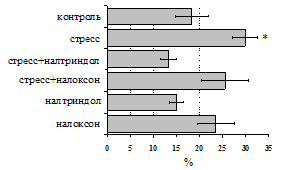
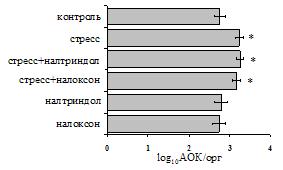
... , в то же время их основная биологическая роль может заключаться в ограничении опасной для организма стрессиндуцированной активации иммунных процессов. ВЫВОДЫ 1. Подтверждена интегральная роль эндогенной опиоидной системы в нейроэндокринной регуляции иммуногенеза в норме, введении глюкокортикоидов и катехоламинов и в условиях стрессорного воздействия. Основные компоненты эндогенной опиоидной ...
... приспособительных реакций у подростков очень важно как в аспекте изучения межсистемных связей в организме, так и в чисто прикладных целях. Это определяется необходимостью прогнозировать состояние иммунной реактивности подростка, выбора оптимальных условий для вакцинации, адекватной терапии заболеваний. В последние годы большое внимание уделяется циркадному, инфрадианному и годичному биоритмам. ...
... комплексов, которые активируют макрофаги, фиксируются на эндотелии и активируют систему комплемента с развитием локального воспаления и повреждением эндотелиоцитов [246]. Двойственный характер влияния иммунной системы на развитие экспериментального атеросклероза и пострансплантацион-ной васкулопатии установлен и при определении роли Т-лимфо-цитов и СВ40-сигнальной системы в развитии иммунного ...
0 комментариев