Навигация
Насосная функция сердца, ее регуляция
44285
знаков
0
таблиц
0
изображений
2. Насосная функция сердца, ее регуляция
Сердце располагается в центре грудной клетки, заключено в тонкую фиброзную околосердечную сумку, перикард, и поддерживается крупными кровеносными сосудами [1, с. 227].
Небольшое количество жидкости в полости перикарда смачивает поверхность сердца и способствует его свободным движениям во время сокращения и расслабления [1, с. 227].
Единственной функцией сердца является обеспечение энергией, которая необходима для циркуляции крови в сердечно-сосудистой системе.
Кровоток через все органы тела осуществляется пассивно и происходит только благодаря тому, что при осуществлении насосной деятельности сердца артериальное давление поддерживается на более высоком уровне, чем венозное
Насос правого сердца создает энергетический импульс, необходимый для передвижения крови через сосуды легких, а насос левого сердца обеспечивает необходимую энергию для перемещения крови через органы тела.
Кровь проходит через трикуспидальный клапан в правый желудочек, а отсюда прогоняется через клапан легочной артерии в легочное кровообращение через легочные артерии. Насыщенная кислородом венозная легочная кровь течет по легочным венам в левое предсердие и проникает через митральный клапан в левый желудочек. Отсюда кровь прогоняется через аортальный клапан в аорту для дальнейшего распределения по органам тела [1, с. 228].
Хотя в целом анатомические характеристики насоса правого сердца несколько отличаются от таковых левого сердца, тем не менее, их деятельность как насосов идентична.
Каждый насос состоит из желудочка, который является закрытой камерой, окруженной мышечной стенкой.
Клапаны имеют такое строение, чтобы кровоток мог осуществляться только в одном направлении, они пассивно открываются и закрываются, реагируя на динамику градиента давления вокруг них.
Насосная деятельность желудочка осуществляется за счет циклического изменения полости желудочков в результате ритмичного и синхронного сокращения и расслабления отдельных клеток сердечной мышцы, которые концентрически располагаются в толще стенки желудочка.
Когда мышечные клетки желудочка сокращаются, то в желудочковой ткани возникает концентрическое напряжение, которое создает постепенно нарастающее давление внутри камеры. Как только желудочковое давление превышает давление в легочной артерии (правый насос) или аорте (левый насос), кровь с силой выбрасывается из камеры через выходной клапан.
Эта фаза сердечного цикла, во время которой сокращаются клетки мускулатуры желудочка, называется систолой Так как во время систолы давление в желудочке выше, чем в предсердии, то атриовентрикулярный (АУ) клалан закрыт.
Когда мышечные клетки желудочка расслабляются, давление в желудочке падает ниже, чем в предсердии, AV клапан открывается и желудочек заполняется вновь кровью. Эта часть сердечного цикла называется диастолой.
Клапан на выходе во время диастолы закрыт, так как артериальное давление выше, чем внутрижелудочковое. После периода диастолического заполнения начинается систолическая фаза нового сердечного цикла.
3. Понятие об обмене веществ. Ассимиляция и диссимиляция. Основной обмен, условия, необходимые для его определения. Рабочий обмен. Прямая и косвенная калориметрия.
В результате обмена веществ непрерывно образуются, обновляются и разрушаются клеточные структуры, синтезируются и разрушаются различные химические соединения [3, с. 291].
В организме динамически уравновешены процессы анаболизма (ассимиляции) — биосинтеза органических веществ, компонентов клеток и тканей, и катаболизма (диссимиляции) — расщепление сложных молекул компонентов клеток.
Преобладание анаболических процессов обеспечивает рост, накопление массы тела, преобладание же катаболических процессов ведет к частичному разрушению тканевых структур, уменьшению массы тела [3, с. 291].
При этом происходит превращение энергии, переход потенциальной энергии химических соединений, освобождаемой при их расщеплении, в кинетическую, в основном тепловую и механическую, частично в электрическую энергию.
Для возмещения энергозатрат организма, сохранения массы тела и удовлетворения потребностей роста необходимо поступление из внешней среды белков, липидов, углеводов, витаминов, минеральных солей и воды. Их количество, свойства и соотношение должны соответствовать состоянию организма и условиям его существования. Это достигается путем питания. Необходимо также, чтобы организм очищался от конечных продуктов распада, которые образуются при расщеплении различных веществ. Это достигается работой органов выделения [3, с. 294].
Интенсивность окислительных процессов и превращение энергии зависят от индивидуальных особенностей организма (пол, возраст, масса тела и рост, условия и характер питания, мышечная работа, состояние эндокринных желез, нервной системы и внутренних органов — печени, почек, пищеварительного тракта), а также от условий внешней среды (температура, барометрическое давление, влажность воздуха и его состав, воздействие лучистой энергии) [3, с. 294].
Для определения присущего данному организму уровня окислительных процессов и энергетических затрат проводят исследование в определенных стандартных условиях. При этом стремятся исключить влияние факторов, которые существенно сказываются на интенсивности энергетических затрат, а именно мышечную работу, прием пищи, влияние температуры окружающей среды.
Энерготраты организма в таких стандартных условиях получили название основного обмена [3, с. 295].
Энерготраты в условиях основного обмена связаны с поддержанием минимально необходимого для жизни клеток уровня окислительных процессов и с деятельностью постоянно работающих органов и систем — дыхательной мускулатуры, сердца, почек, печени [3, с. 295].
Некоторая часть энерготрат в условиях основного обмена связана с поддержанием мышечного тонуса.
Освобождение в ходе всех этих процессов тепловой энергии обеспечивает ту теплопродукцию, которая необходима для поддержания температуры тела на постоянном уровне, как правило, превышающем температуру внешней среды.
Для определения основного обмена обследуемый должен находиться:
1) в состоянии мышечного покоя (положение лежа с расслабленной мускулатурой), не подвергаясь раздражениям, вызывающим эмоциональное напряжение;
2) натощак, т. е. через 12— 16 ч после приема пищи;
3) при внешней температуре «комфорта» (18—20 °С), не вызывающей ощущения холода или жары [3, с. 295].
Основной обмен определяют в состоянии бодрствования.
Во время сна уровень окислительных процессов и, следовательно, энергетических затрат организма на 8—10 % ниже, чем в состоянии покоя при бодрствовании [3, с. 295].
Нормальные величины основного обмена человека. Величину основного обмена обычно выражают количеством тепла в килоджоулях (килокалориях) на 1 кг массы тела или на 1 м2 поверхности тела за 1 ч или за одни сутки [3, с. 296].
Для мужчины среднего возраста (примерно 35 лет), среднего роста (примерно 165 см) и со средней массой тела (примерно 70 кг) основной обмен равен 4,19 кДж (1 ккал) на 1 кг массы тела в час, или 7117 кДж (1700 ккал) в сутки. У женщин той же массы он примерно на 10 % ниже.
Интенсивность основного обмена, пересчитанная на 1 кг массы тела, у детей значительно выше, чем у взрослых. Величина основного обмена человека в возрасте 20—40 лет сохраняется на довольно постоянном уровне. В пожилом возрасте основной обмен снижается.
Согласно формуле Дрейера, суточная величина основного обмена в килокалориях (H) составляет:
H=W/K∙A0,1333 (1)
где W — масса тела, г;
А — возраст человека;
К — константа, равная для мужчины 0,1015, а для женщины — 0,1129.
Формулы и таблицы основного обмена представляют средние данные, выведенные из большого числа исследований здоровых людей разного пола, возраста, массы тела и роста.
Определение основного обмена, согласно этим таблицам, у здоровых людей нормального телосложения дают приблизительно верные (ошибка 5—8 %) величины затраты энергии. Несоразмерно высокие данные для определенной массы тела, роста, возраста и поверхности тела величины основного обмена наблюдаются при избыточной функции щитовидной железы.
Понижение основного обмена встречается при недостаточности щитовидной железы (микседема), гипофиза, половых желез.
Если пересчитать интенсивность основного обмена на 1 кг массы тела, то окажется, что у теплокровных животных разных видов, и у людей с разной массой тела и ростом она весьма различна.
Если же произвести перерасчет интенсивности основного обмена на 1 м2 поверхности тела, полученные у разных животных и людей величины различаются не столь резко [3, с. 296].
Согласно правилу поверхности тела, затраты энергии теплокровными животными пропорциональны величине поверхности тела.
Ежедневная продукция тепла на 1 м2поверхности тела у человека равна 3559 — 5234 кДж (850—1250 ккал), средняя цифра для мужчин — 3969 кДж (948 ккал).
Для определения поверхности тела R применяется формула:
R = К • масса тела (2)
Эта формула выведена на основании анализа результатов прямых измерений поверхности тела. Константа К у человека равна 12,3.
Более точная формула предложена Дюбуа:
R= W0,425 ∙ H0,725 ∙ 71,84 (3)
где W — масса тела в килограммах,
Н — рост в сантиметрах.
Результат вычисления выражен в квадратных сантиметрах.
Правило поверхности верно не абсолютно. Как показано выше, оно представляет собой лишь правило, имеющее известное практическое значение для ориентировочных расчетов освобождения энергии в организме.
Об относительности правила поверхности свидетельствует тот факт, что у двух индивидуумов с одинаковой поверхностью тела интенсивность обмена веществ может значительно различаться. Уровень окислительных процессов определяется не столько теплоотдачей с поверхности тела, сколько теплопродукцией, зависящей от биологических особенностей вида животных и состояния организма, которое обусловлено деятельностью нервной, эндокринной и других систем.
Деятельное состояние вызывает заметную интенсификацию обмена веществ. Обмен веществ при этих условиях называется рабочим обменом. Если основной обмен взрослого человека равен 1700-1800 ккал, то рабочий обмен в 2-3 раза выше [5, с. 212].
Таким образом, основной обмен исходным фоновым уровнем потребления энергии. Резкое изменение основного обмена может быть важным диагностическим признаком переутомления, перенапряжения и недовосстановления или заболевания.
Прямая калориметрия основана на непосредственном учете в биокалориметрах количества тепла, выделенного организмом.
Биокалориметр представляет собой герметизированную и хорошо теплоизолированную от внешней среды камеру.
В камере по трубкам циркулирует вода. Тепло, выделяемое находящимся в камере человеком или животным, нагревает циркулирующую воду. По количеству протекающей воды и изменению ее температуры рассчитывают количество выделенного организмом тепла [5, с. 212].
Одновременно в биокалориметр подается О2 и поглощается избыток СО2 и водяных паров.
Методы прямой калориметрии очень громоздки и сложны. Учитывая, что в основе теплообразования в организме лежат окислительные процессы, при которых потребляется О2 и образуется СО2, можно использовать косвенное, непрямое, определение теплообразования в организме по его газообмену — учету количества потребленного О2 и выделенного СО2 с последующим расчетом теплопродукции организма.
Для длительных исследований газообмена используют специальные респираторные камеры (закрытые способы непрямой калориметрии). Кратковременное определение газообмена в условиях лечебных учреждений и производства проводят более простыми не камерными методами (открытые способы калориметрии).
Наиболее распространен способ Дугласа — Холдейна, при котором в течение 10—15 мин собирают выдыхаемый воздух в мешок из воздухонепроницаемой ткани (мешок Дугласа), укрепляемый на спине обследуемого.
Он дышит через загубник, взятый в рот, или резиновую маску, надетую на лицо. В загубнике и маске имеются клапаны, устроенные так, что обследуемый свободно вдыхает атмосферный воздух, а выдыхает воздух в мешок Дугласа. Когда мешок наполнен, измеряют объем выдохнутого воздуха, в котором определяют количество О2 и СО2.
Кислород, поглощаемый организмом, используется для окисления белков, жиров и углеводов.
Окислительный распад 1 г каждого из этих веществ требует неодинакового количества О2 и сопровождается освобождением различного количества тепла. При потреблении организмом 1 л О2 освобождается разное количество тепла в зависимости от того, на окисление каких веществ О2 используется.
Количество тепла, освобождающегося после потребления организмом 1 л О2, носит название калорического эквивалента кислорода.
Зная общее количество О2, использованное организмом, можно вычислить энергетические затраты только в том случае, если известно, какие вещества — белки, жиры или углеводы, окислились в теле. Показателем этого может служить дыхательный коэффициент.
Дыхательным коэффициентом (ДК) называется отношение объема выделенного СО2 к объему поглощенного О2. Дыхательный коэффициент различен при окислении белков, жиров и углеводов. Для примера рассмотрим, каков будет дыхательный коэффициент при использовании организмом глюкозы. Общий итог окисления молекулы глюкозы можно выразить формулой:
С6Н12О6 + 6 О2 = 6 СО2 + 6 Н2О (4)
При окислении глюкозы число молекул образовавшегося СО2 равно числу молекул затраченного (поглощенного) О2.
Равное количество молекул газа при одной и той же температуре и одном и том же давлении занимает один и тот же объем (закон Авогадро— Жерара). Следовательно, дыхательный коэффициент (отношение СО2/О2) при окислении глюкозы и других углеводов равен единице.
При окислении жиров и белков дыхательный коэффициент будет ниже единицы. При окислении жиров дыхательный коэффициент равен 0,7. Проиллюстрируем это на примере окисления трипальмитина:
2 С3Н5 (С15Н31СОО)3 + 145 О2 = 102 СО2 + 98 Н2О (5)
Отношение между объемами углекислого газа и кислорода составляет в данном случае:
102 CO2/45 O2= 0,703 (6)
Аналогичный расчет можно сделать и для белка; при его окислении в организме дыхательный коэффициент равен 0,8.
При смешанной пище у человека дыхательный коэффициент обычно равен 0,85—089. Определенному дыхательному коэффициенту соответствует определенный калорический эквивалент кислорода.
Определение энергетического обмена у человека в покое методом закрытой системы с неполным газовым анализом.
Относительное постоянство дыхательного коэффициента (0,85—0,90) у людей при обычном питании в условиях покоя позволяет производить достаточно точное определение энергетического обмена у человека в покое, вычисляя только количество потребленного кислорода и беря его калорический эквивалент при усредненном дыхательном коэффициенте. Количество потребленного организмом кислорода определяют при помощи различных спирографов.
Определив количество поглощенного кислорода и приняв усредненный дыхательный коэффициент равным 0,85, можно рассчитать энергообразование в организме; калорический эквивалент 1 л кислорода при данном дыхательном коэффициенте равен 20,356 кДж, т. е. 4,862 ккал [5, с. 213].
Способ неполного газового анализа благодаря своей простоте получил широкое распространение.
Во время интенсивной мышечной работы дыхательный коэффициент повышается и в большинстве случаев приближается к единице. Это объясняется тем, что главным источником энергии во время напряженной мышечной деятельности является окисление углеводов. После завершения работы дыхательный коэффициент в течение первых нескольких минут так называемого периода восстановления резко снижается до величин меньших, чем исходные, и только спустя 30—50 мин после напряженной работы обычно нормализуется.
Изменения дыхательного коэффициента после окончания работы не отражают истинного отношения между используемым в данный момент кислородом и выделенной СО2.
Дыхательный коэффициент в начале восстановительного периода повышается по следующей причине: в мышцах во время работы накапливается молочная кислота, на окисление которой во время работы не хватало О2 (это так называемый кислородный долг).
Молочная кислота поступает в кровь и вытесняет СО2 из гидрокарбонатов, присоединяя основания. Благодаря этому количество выделенного СО2 больше количества СO2, образовавшегося в данный момент в тканях.
Обратная картина наблюдается в дальнейшем, когда молочная кислота постепенно исчезает из крови.
Одна часть ее окисляется, другая ресинтезируется в гликоген, а третья выделяется с мочой и потом. По мере уменьшения количества молочной кислоты освобождаются основания, которые до того были отняты у гидрокарбонатов.
Эти основания вновь связывают СО2 и образуют гидрокарбонаты, поэтому через некоторое время после работы дыхательный коэффициент резко падает вследствие задержки в крови СО2, поступающей из тканей.
4. Продолговатый мозг. Классификация нервных центров. Функции продолговатого мозга, их характеристика. Особенности функционирования жизненных важных центров. Варолиев мост (мост мозга), его функциональное значение.
Продолговатый мозг (medulla oblongata) у человека имеет длину около 25 мм. Он является продолжением спинного мозга. Структурно по разнообразию и строению ядер продолговатый мозг сложнее, чем спинной. В отличие от спинного мозга он не имеет метамерного, повторяемого строения, серое вещество в нем расположено не в центре, а ядрами к периферии [5, с. 172].
В продолговатом мозге находятся оливы, связанные со спинным мозгом, экстрапирамидной системой и мозжечком — это тонкое и клиновидное ядра проприоцептивной чувствительности (ядра Голля и Бурдаха). Здесь же находятся перекресты нисходящих пирамидных путей и восходящих путей, образованных тонким и клиновидным пучками (Голля и Бурдаха), ретикулярная формация.
Продолговатый мозг за счет своих ядерных образований и ретикулярной формации участвует в реализации вегетативных, соматических, вкусовых, слуховых, вестибулярных рефлексов.
Особенностью продолговатого мозга является то, что его ядра, возбуждаясь последовательно, обеспечивают выполнение сложных рефлексов, требующих последовательного включения разных мышечных групп, что наблюдается, например, при глотании [5, с. 172].
В продолговатом мозге расположены ядра следующих черепных нервов:
- пара VIII черепных нервов — преддверно-улитковый нерв состоит из улитковой и преддверной частей. Улитковое ядро лежит в продолговатом мозге;
- пара IX — языкоглоточный нерв (п. glossopharyngeus); его ядро образовано 3 частями — двигательной, чувствительной и вегетативной. Двигательная часть участвует в иннервации мышц глотки и полости рта, чувствительная — получает информацию от рецепторов вкуса задней трети языка; вегетативная иннервирует слюнные железы;
- пара X — блуждающий нерв (n. vagus) имеет 3 ядра: вегетативное иннервирует гортань, пищевод, сердце, желудок, кишечник, пищеварительные железы; чувствительное получает информацию от рецепторов альвеол легких и других внутренних органов и двигательное (так называемое обоюдное) обеспечивает последовательность сокращения мышц глотки, гортани при глотании;
- пара XI — добавочный нерв (n. accessorius); его ядро частично расположено в продолговатом мозге;
- пара XII — подъязычный нерв (n. hypoglossus) является двигательным нервом языка, его ядро большей частью расположено в продолговатом мозге.
Рефлекторная деятельность организма во многом определяется общими свойствами нервных центров [5, с. 170].
Нервный центр — совокупность структур центральной нервной системы, координированная деятельность которых обеспечивает регуляцию отдельных функций организма или определенный рефлекторный акт. Представление о структурно-функциональной основе нервного центра обусловлено историей развития учения о локализации функций в центральной нервной системе [5, с. 170].
На смену старым теориям об узкой локализации, или эквипотенциальности, высших отделов головного мозга, в частности коры большого мозга, пришло современное представление о динамической локализации функций, основанное на признании существования четко локализованных ядерных структур нервных центров и менее определенных рассеянных элементов анализаторных систем мозга.
При этом с цефализацией нервной системы растут удельный вес и значимость рассеянных элементов нервного центра, внося существенные различия в анатомических и физиологических границах нервного центра. В результате функциональный нервный центр может быть локализован в разных анатомических структурах.
Например, дыхательный центр представлен нервными клетками, расположенными в спинном, продолговатом, промежуточном мозге, в коре большого мозга.
Нервные центры имеют ряд общих свойств, что во многом определяется структурой и функцией синаптических образований.
1. Односторонность проведения возбуждения. В рефлекторной дуге, включающей нервные центры, процесс возбуждения распространяется в одном направлении (от входа, афферентных путей к выходу, эфферентным путям).
2. Иррадиация возбуждения. Особенности структурной организации центральных нейронов, огромное число межнейронных со единений в нервных центрах существенно модифицируют (изменяют) направление распространения процесса возбуждения в зависимости от силы раздражителя и функционального состояния центральных нейронов. Значительное увеличение силы раздражителя приводит к расширению области вовлекаемых в процесс возбуждения центральных нейронов — иррадиации возбуждения.
3. Суммация возбуждения. В работе нервных центров значительное место занимают процессы пространственной и временной суммации возбуждения, основным нервным субстратом которой является постсинаптическая мембрана. Процесс пространственной суммации афферентных потоков возбуждения облегчается наличием на мембране нервной клетки сотен и тысяч синаптических контактов. Процессы временной суммации обусловлены суммацией ВПСП на постсинаптической мембране.
4. Наличие синаптической задержки. Время рефлекторной ре акции зависит в основном от двух факторов: скорости движения возбуждения по нервным проводникам и времени распространения возбуждения с одной клетки на другую через синапс.
При относительно высокой скорости распространения импульса по нервному проводнику основное время рефлекса приходится на синаптическую передачу возбуждения (синаптическая задержка). В нервных клетках высших животных и человека одна синаптическая задержка примерно равна 1 мс. Если учесть, что в реальных рефлекторных дугах имеются десятки последовательных синаптических контактов, становится понятной длительность большинства рефлекторных реакций — десятки миллисекунд.
5. Высокая утомляемость. Длительное повторное раздражение рецептивного поля рефлекса приводит к ослаблению рефлекторной реакции вплоть до полного исчезновения, что называется утомлением. Этот процесс связан с деятельностью синапсов — в последних наступает истощение запасов медиатора, уменьшаются энергетические ресурсы, происходит адаптация постсинаптического рецептора к медиатору.
6. Тонус. Тонус, или наличие определенной фоновой активности нервного центра, определяется тем, что в покое в отсутствие специальных внешних раздражений определенное количество нервных клеток находится в состоянии постоянного возбуждения, генерирует фоновые импульсные потоки. Даже во сне в высших отделах мозга остается некоторое количество фоновоактивных нервных клеток, формирующих «сторожевые пункты» и определяющих некоторый тонус соответствующего нервного центра.
7. Пластичность. Функциональная возможность нервного центра существенно модифицировать картину осуществляемых рефлекторных реакций. Поэтому пластичность нервных центров тесно связана с изменением эффективности или направленности связей между нейронами.
8. Конвергенция. Нервные центры высших отделов мозга являются мощными коллекторами, собирающими разнородную афферентную информацию. Количественное соотношение периферических рецепторных и промежуточных центральных нейронов (10:1) предполагает значительную конвергенцию («сходимость») разномодальных сенсорных посылок на одни и те же центральные нейроны. На это указывают прямые исследования центральных нейронов: в нервном центре имеется значительное количество поливалентных, полисенсорных нервных клеток, реагирующих на разномодальные стимулы (свет, звук, механические раздражения).
Конвергенция на клетках нервного центра разных афферентных входов предопределяет важные интегративные, перерабатывающие информацию функции центральных нейронов, т. е. высокий уровень интеграционных функций. Конвергенция нервных сигналов на уровне эфферентного звена рефлекторной дуги определяет физиологический механизм принципа «общего конечного пути» по Ч. Шеррингтону.
9. Интеграция в нервных центрах. Важные интегративные функции клеток нервных центров ассоциируются с интегративными процессами на системном уровне в плане образования функциональных объединений отдельных нервных центров в целях осуществления сложных координированных приспособительных целостных реакций организма (сложные адаптивные поведенческие акты).
10. Свойство доминанты. Доминантным называется временно господствующий в нервных центрах очаг (или доминантный центр) повышенной возбудимости в центральной нервной системе. По А.А.Ухтомскому, доминантный нервный очаг характеризуется такими свойствами, как повышенная возбудимость, стойкость и инертность возбуждения, способность к суммированию возбуждения.
В доминантном очаге устанавливается определенный уровень стационарного возбуждения, способствующий суммированию ранее подпороговых возбуждений и переводу на оптимальный для данных условий ритм работы, когда этот очаг становится наиболее чувствительным. Доминирующее значение такого очага (нервного центра) определяет его угнетающее влияние на другие соседние очаги возбуждения. Доминантный очаг возбуждения «притягивает» к себе возбуждение других возбужденных зон (нервных центров). Принцип доминанты определяет формирование главенствующего (активирующего) возбужденного нервного центра в тесном соответствии с ведущими мотивами, потребностями организма в конкретный момент времени.
11. Цефализация нервной системы. Основная тенденция в эволюционном развитии нервной системы проявляется в перемещении, сосредоточении функции регуляции и координации деятельности организма в головных отделах ЦНС.
Этот процесс называется цефализацией управляющей функции нервной системы. При всей сложности складывающихся отношений между старыми, древними и эволюционно новыми нервными образованиями стволовой части мозга общая схема взаимных влияний может быть представлена следующим образом: восходящие влияния (от нижележащих «старых» нервных структур к вышележащим «новым» образованиям) преимущественно носят возбуждающий стимулирующий характер, нисходящие (от вышележащих «новых» нервных образований к нижележащим «старым» нервным структурам) носят угнетающий тормозной характер. Эта схема согласуется с представлением о росте в процессе эволюции роли и значения тормозных процессов в осуществлении сложных интегративных рефлекторных реакций.
Продолговатый мозг выполняет ряд следующих функций:
Похожие работы
... и утрачивается к началу полового созревания. 36. Учение Павлова о типах ВНД. Общин типы ВНД. Пластичность ВНД. Учет типов ВНД в индивидуальном обучении и воспитании Среди вопросов физиологии ВНД человека особое значение для совершенствования учебно-воспитательного процесса и разработки естественнонаучных теорий воспитания и обучения имеет учение о типах ВНД. Это связано с тем, что именно ...
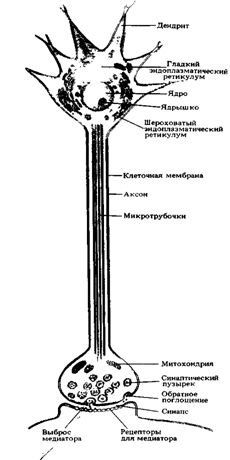
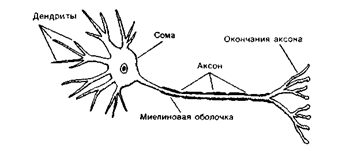
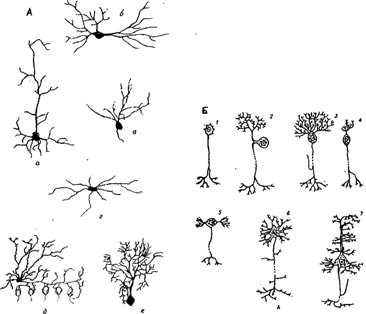
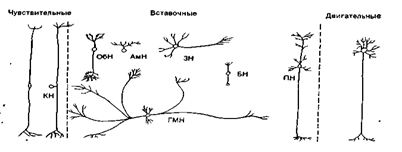
... мозгу (1), где через вставочный нейрон передаются на эфферентные волокна (эфф. нерв), по которым доходят до эффектора. Пунктирные линии - распространение возбуждения от низших отделов центральной нервной системы на ее вышерасположенные отделы (2, 3,4) до коры мозга (5) включительно. Наступающее вследствие этого изменение состояния высших отделов мозга в свою очередь воздействует (см. стрелки) на ...
... также следует учитывать при применении некоторых ЛС (например, снотворных длительного действия). Обилие лекарственных веществ, известных современной медицине, отнюдь не означает, что каждый препарат обладает индивидуальным механизмом действия. Понимание механизма действия важно не только для фармаколога, занятого поиском совершенных препаратов, но и помогает правильно использовать их в клинике. ...
... условным, так как в организме любой из уровней не является автономным, низшие уровни соподчиняются – высшим. 5. Особенности функционирования вегетативной нервной системы при интенсивных нагрузках Вегетативная нервная система осуществляет регуляцию деятельности всех висцеральных систем организма, принимает участие в гомеостатических реакциях организма, выполняет адаптационно-трофическую ...
0 комментариев