Мембрана клетки
План Емкость мембраны Постоянная времени
Емкость в кабеле
Сенситизация и S интернейроны
Вывод
Емкость мембраны
Мембрана клетки не только проводит ионные токи, но и накапливает заряд на своей внешней или внутренней поверхности. С точки зрения теории электричества, разделение зарядов на мембране означает, что мембрана обладает свойствами конденсатора. В общем виде конденсатор состоит из двух проводящих пластин, отделенных друг от друга изолирующим материалом; в промышленных конденсаторах проводящие пластины обычно сделаны из металлической фольги, а изолирующая прослойка между ними — из пластика. В случае нервной клетки проводниками являются два слоя жидкости, находящиеся по обе стороны мембраны, а сама мембрана играет роль изолирующей прослойки. При зарядке конденсатора от батареи на одной из пластинок накапливается положительный заряд, в то время как на второй пластинке создается равный по величине запас отрицательного заряда. Емкость конденсатора (С) определяется количеством заряда (Q), запасаемым на один вольт потенциала (V), приложенного к пластинам конденсатора: С = Q/V. С измеряется в кулонах, деленных на вольт, т. е. в фарадах (Ф). Чем ближе друг к другу расположены пластины, тем более эффективно конденсатор способен разделять и накапливать заряд. Поскольку толщина мембраны клетки всего 5 нм, она способна накапливать достаточно большой заряд. Обычно емкость мембраны нервных клеток составляет 1 мкФ/см2. Преобразовав выражение, получаем Q = CV. При потенциале покоя —80 мВ, количество избыточного отрицательного заряда на внутренней стороне мембраны составит (1 · 10–6) х (80 ·10–3) = 8· 10–8 кулонов, деленных на см2, что соответствует 5 · 1011 одновалентных ионов (0,8 пмоль) на квадратный сантиметр мембраны.
Величину тока, протекающего внутрь конденсатора или из него, можно подсчитать на основе соотношения заряда и напряжения, учитывая, что ток (i, в амперах) есть скорость изменения заряда во времени, т.е. 1 ампер = (1 кулон)/(1 с). Поскольку Q = C/V, получим:
![]()
Скорость изменения заряда на конденсаторе прямо пропорциональна величине тока. Если ток постоянен, то потенциал будет меняться с постоянной скоростью dV/dt = i/C.
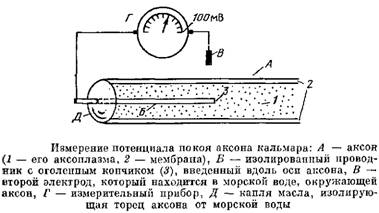
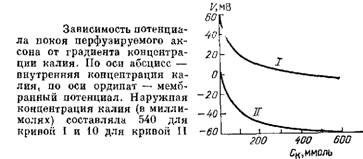
Соотношение тока и напряжения в цепи, содержащей резисторы (сопротивления) и конденсаторы (емкости), соединенные параллельно. Прямоугольный скачок тока величиной i, приложенный к резистору (R), создает скачок напряжения на резисторе величиной V = iR. Если тот же скачок тока приложить к конденсатору (С), то напряжение на конденсаторе будет накапливаться со скоростью dV/dt = i/C. Когда эти два элемента, резистор и конденсатор, соединены параллельно, то весь ток пойдет сначала на зарядку конденсатора со скоростью i/С; однако, как только на конденсаторе накопится какой-то заряд, ток потечет и через резистор. По мере нарастания тока, все большая его часть будет проходить через сопротивление, потому что скорость зарядки конденсатора будет постепенно снижаться. В конце концов весь ток будет течь через резистор, создавая на нем потенциал V = iR, a конденсатор будет полностью заряжен. По завершении скачка тока заряд из конденсатора постепенно рассеется на резисторе, а напряжение вернется к нулю. Из экспериментов на аксоне кальмара можно заключить, что гипотеза, высказанная Бернштейном в 1902 г., была близка к истине: трансмембранный градиент калия является важным, хотя и не единственным фактором, влияющим на мембранный потенциал. Чем можно объяснить отклонение экспериментальной кривой от уравнения Нернста? Оказывается, для этого достаточно снять ограничение с модели, состоящее в том, что мембрана непроницаема для ионов натрия. Мембрана реальной клетки действительно обладает натриевой проницаемостью, которая составляет от 1 до 10 % калиевой.
Для рассмотрения роли натриевой проницаемости обратимся к модели идеальной клетки и временно исключим из поля зрения перемещение ионов хлора. Мембранный потенциал равен калиевому равновесному потенциалу, поэтому перемещение суммарного заряда через мембрану отсутствует, клетка находится в покое. Если теперь ввести в модель натриевую проницаемость, то натрий будет стремиться войти в клетку благодаря как своему концентрационному градиенту, так и мембранному потенциалу. По мере входа натрия на внутренней поверхности мембраны накапливается положительный заряд и мембрана деполяризуется. В результате ионы калия выходят из равновесия и начинают покидать клетку. С увеличением деполяризации мембраны движущая сила для входа натрия снижается, в то время как движущая сила для выхода калия возрастает. Процесс продолжается до тех пор, пока оба ионных потока не уравновесят друг друга. В этот момент изменение мембранного потенциала прекращается, поскольку какое-либо накопление заряда отсутствует. Вообще говоря, значение мембранного потенциала расположено между калиевым и натриевым равновесными потенциалами и определяется равновесием между калиевым и натриевым токами, равными по величине и направленными в противоположные стороны.
Ионы хлора также участвуют в этом процессе, однако, как мы убедились ранее, равновесный потенциал для хлора подстраивается под новое значение мембранного потенциала за счет изменения внутриклеточной концентрации этого иона. По мере того как токи катионов постепенно приходят в равновесие, внутриклеточный уровень хлора возрастает до тех пор, пока суммарный хлорный ток не станет равным нулю.
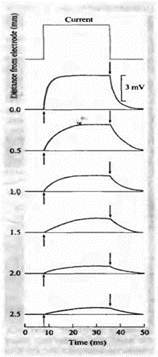
Рис. 1. Распространение потенциала вдоль аксона омара, регистрируемое с помощью поверхностного электрода.
Постоянная времениНарастание и спад потенциала происходит по экспоненциальной кривой, как показано на рис. 1В. Фаза роста описывается уравнением:
![]()
где t — время от начала импульса. Постоянная времени равна произведению RC. Это время, за которое потенциал возрастаетдо 63 % (1 - 1/e) своего максимального значения. Спад напряжения тоже экспоненциален, с той же постоянной времени. Ток через резистор, iR, должен изменяться во времени по тому же закону, что и напряжение. Следовательно, на фазе роста ток начинает расти от нуля до своего максимального значения i. Емкостной ток, наоборот, начинается с величины i и спадает до нуля по экспоненте с той же постоянной времени. По завершении импульса, поскольку внешнего источника тока нет, единственным током на резисторе будет ток, генерируемый напряжением на конденсаторе. Следовательно, ток на резисторе равен по величине емкостному току и противоположен ему по направлению, как показано на рисунке.
Вышеописанную цепь, состоящую из параллельно соединенных резистора и конденсатора, можно использовать для описания сферической нервной клетки, аксон и дендриты которой настолько малы, что их вкладом в электрические свойства клеток можно пренебречь. В эквивалентной цепочке для аксона или мышечного волокна как емкость мембраны, так и ее сопротивление распределены по всей длине волокна, как показано на рис. 7.2D. Емкость мембраны на единицу длины ст (измеряемая в мкФ/см) зависит от удельной емкости на единицу площади Сm (в мкФ/см2) по формуле сm = 2Ст, где — радиус волокна.
Постоянная времени мембраны сферической клетки или волокна не зависит от размера клетки или волокна. Причина этого в том, что увеличение радиуса (а следовательно, и площади поверхности мембраны) влечет за собой не только увеличение емкости, но и соответствующее снижение сопротивления, так что произведение двух величин не меняется. Поскольку показано, что величина Ст приблизительно одинакова для всех нервных и мышечных волокон (1 мкФ/см2), то величина τ является удобным параметром, характеризующим удельное сопротивление мембраны для данной клетки. Постоянная времени — это третий параметр, который, наряду с входным сопротивлением и постоянной длины, характеризует поведение аксона. Диапазон значений постоянной времени в различных типах нервных и мышечных клеток составляет от 1 до 20 мс.
Емкость в кабелеКаково влияние постоянной времени на ток в кабеле? Как и в случае простой RC-цепочки (рис. 1С), нарастание и спад потенциала в ответ на скачкообразное изменение тока замедляется благодаря наличию конденсатора. Ситуация осложняется тем, что ток течет уже не через один конденсатор, но каждый сегмент цепочки является одним из резистивно-емкостных элементов, взаимодействующих между собой. Благодаря этим взаимодействиям временной ход в отдельном сегменте нельзя описать простой экспонентой, и фазы роста и спада замедляются по мере удаления от точки инъекции тока (рис. 2). Поскольку скорость нарастания потенциала зависит от расстояния между отводящим электродом и местом инъекции тока, постоянную времени спада уже нельзя рассчитать на основе простого измерения времени 63 %-ного роста потенциала, за исключением единственной точки вдоль волокна, в которой это расстояние равно постоянной длины.
Обратимся к рассмотрению движения ионов. Если в аксон инъецирован положительный ток, внутриклеточные ионы (главным образом, калия) будут распространяться вдоль волокна. Некоторая часть ионов уйдет на изменение заряда на емкости мембраны, другая часть будет протекать по мембранному сопротивлению. Одновременно с этим процессом будет происходить перемещение отрицательных ионов в противоположном направлении. Постепенно потенциал на мембране достигнет нового устойчивого значения, емкости, распределенные по мембране, будут полностью заряжены до нового уровня потенциала, а через мембрану будет протекать постоянный ионный ток. Время, необходимое для достижения нового устойчивого состояния, определяется постоянной времени.
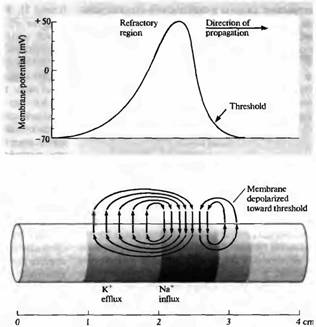
Рис. 2. Мембранный ток во время прохождения потенциала действия.
.
Механизм того, как отдельный нейрон может играть координирующую роль в поведении животного, тщательным образом на насекомых и ракообразных, у которых были описаны отдельные «командные» нейроны, способные дирижировать оркестром поведенческих реакций всего организма. У пиявки эту роль играет непарный интернейрон, присутствующий в каждом ганглии и называемый S клеткой. Этот нейрон получает возбуждающие импульсы от чувствительных клеток при касании и давлении и в свою очередь возбуждает L мотонейрон, который способствует сокращению тела (как ранее было указано). Каждая S клетка связана с S клеткой соседних сегментов посредством толстого аксона с большой скоростью проведения через электрический синапс. То, что синапс находится посередине между сегментами, можно определить путем введения в S клетку пероксидазы хрена, которая не может распространяться через щелевые контакты.
Последовательная цепь S клеток является ключевым механизмом адаптивных реакций пиявки. При повторной тактильной стимуляции прикосновением к определенному сегменту тела сначала возникает рефлекс сокращения. Ответ становится все слабее и слабее при каждом следующем прикосновении — данный эффект известен как «привыкание» (habituation). Однако после более сильного раздражения, когда активируются Ρ (давление) и N (боль) клетки, ответ на дальнейшую стимуляцию только Т клеток (при касании) вновь становится заметным. Этот процесс известен как «отвыкание» (dishabituation) Сходным же образом, если более сильный раздражитель наносится до процедуры «обучения», это вызывает сенситизацию, т. е. ответ на стимуляцию при касании будет больше обычного. Активность S клеток во время «привыкания» не меняется, однако возрастает как при сенситизации, так и при «отвыкании» В технически сложно выполнимых экспериментах, когда избирательно перерезается аксон S клетки, либо клетки убиваются инъекцией проназ, процессы «привыкания» и сокращения не изменяются, однако полностью исчезает «отвыкание» и сенситизация.
Во второй серии экспериментов аксоны S клеток были повреждены, чтобы исследовать их регенерацию. Удивительным свойством S клеток является то, что после повреждения их аксоны следуют по старому пути и заново образуют электрические соединения с соседним S нейроном с поразительной точностью. Как и ожидалось, перерыв передачи импульсов по S клеткам между отдельными сегментами приводил к исчезновению сенситизации, однако через несколько недель после полной регенерации аксонов и восстановления прежних связей данное свойство также восстанавливалось.
Эти примеры наглядно демонстрируют, каким образом отдельная нервная клетка способна принимать участие в таком сложном поведенческом процессе, как сенситизация. Было установлено, что на самом деле процессы сенситизации во многом обусловлены серотонином. (Необычная серия совпадений способствовала тому, что название «S клетка» стало очень подходящим для данных клеток. Изначально она была так названа, потому что ее импульс образует долгий импульс (spike) в месте контакта. Затем, обнаружив, что эти клетки очень тесно связаны друг с другом, полагали, что они могут быть синцитием. Сейчас же они называются S клетками от слов «сенситизация» и «серотонин»!) У Aplysia процесс сенситизации (в котором серотонин тоже играет важную роль) был очень тщательно изучен как на клеточном, так и на молекулярном уровне).
Вывод
Короткие сигналы распространяются на более короткие расстояния, чем длительные сигналы. В случае достаточной длительности сигнала, в течение которого потенциал успевает достигнуть своего максимального значения, емкость заряжается полностью, и пространственное распределение потенциала определяется сопротивлениями мембраны и цитоплазмы: Vx = V0ε–x/. Для коротких импульсов, таких как синаптический потенциал, ток прекращается еще до того, как емкость успевает полностью зарядиться. Это выражаетсяв уменьшении расстояния, на которое потенциал распространяется вдоль волокна. Другими словами, эффективная постоянная длины для коротких сигналов меньше, чем для длительных. Кроме того, форма коротких сигналов искажается по мере их перемещения по волокну, а их амплитуда снижается за счет «закругления» пика, достигаемого все позднее и позднее
Литература
1. Пенроуз Р. НОВЫЙ УМ КОРОЛЯ. О компьютерах, мышлении и законах физики.
2. Грегори Р. Л. Разумный глаз.
3. Леках В. А. Ключ к пониманию физиологии.
4. Гамов Г., Ичас М. Мистер Томпкинс внутри самого себя: Приключения в новой биологии.
Похожие работы
... полюсам ядра. Ядерная мембрана разрушается; одновременно специальные белки объединяются (агрегируют), формируя микротрубочки в виде нитей. Центриоли, расположенные теперь на противоположных полюсах клетки, оказывают организующее воздействие на микротрубочки, которые в результате выстраиваются радиально, образуя структуру, напоминающую по внешнему виду цветок астры («звезда»). Другие нити из ...
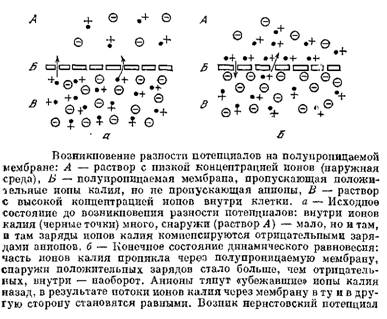
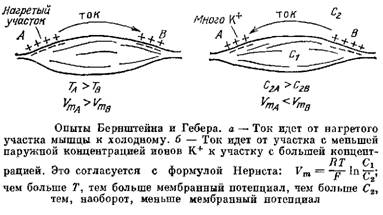
... много, наружу. Вместе с ними будет выноситься наружу их положительный заряд. Внутрь через мембрану будет проходить мало ионов, так как снаружи мало калия. В результате на клеточной мембране будет возникать разность потенциалов: снаружи клетки — «плюс», а внутри — «минус». Эта разность потенциалов будет тормозить движение новых положительных заряженных ионов калия наружу, и увеличивать поток этих ...
... , и эти два органоида, как отмечено выше, представляют единый аппарат синтеза и транспортировки образующихся в клетке белков. Комплекс Гольджи. Комплекс Гольджи – органоид клетки, названный так по имени итальянского ученого К. Гольджи, который впервые увидел его в цитоплазме нервных клеток (1898) и обозначил как сетчатый аппарат. Сейчас комплекс Гольджи обнаружен во всех клетках растительных и ...
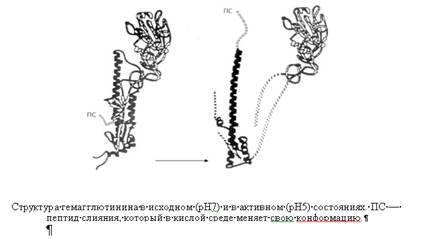
... , вирус Эбола и другие, способны ассоциироваться с "плотами" клеточной мембраны. Также филовирусы Эбола и Марбурга проникают в клетки путем кавеолин-опосредованного эндоцитоза, при котором белок кавеолин связан с микродоменами плазмалеммы. Механизм проникновения вирусов путем эндоцитоза, независимого от клатрина и кавеолина и опосредованного липидными площадками плазмалеммы - "плотами", главным ...
0 комментариев