РЕФЕРАТ
Миграция клеток и воспаление
Введение
В норме лейкоциты, циркулирующие с кровью, мигрируют по всем тканям организма, но при этом каждая их популяция имеет свой особый характер миграции. Кроме того, путь миграции зависит от стадии дифференцировки и уровня активации клеток:
• фагоциты, в том числе нейтрофилы и моноциты, покинув костный мозг, мигрируют в те периферические ткани организма, где имеются очаги воспаления; для нейтрофилов это путешествие в одном направлении, но моноциты, превратившись в макрофаги, могут вернуться во вторичные лимфоидные ткани и функционировать там в качестве антигенпрезентирующих клеток:
• непримированные лимфоциты мигрируют из тимуса и костного мозга во вторичные лимфоидные ткани; после активации антигеном Т-клетки стремятся проникнуть в очаг воспаления, тогда как В-клетки и Т-клетки иммунологической памяти расселяются по соседним лимфоидным органам и образованиям.
• дендритные клетки, в частности кожные клетки Лангерганса, представляют собой потомство костномозговых стволовых клеток, заселившее вторичные лимфоидные ткани; захватив антиген, они могут мигрировать в регионарные лимфоузлы, чтобы презентировать его Т-клеткам CD4+. Миграция лимфоцитов позволяет каждой "горстке" лимфоцитов, специфичных к какому-либо отдельному антигену, встретиться именно с ним. Пути оттока лимфы и перемещения клеток обеспечивают встречу лимфоцитов, АПК и поступающего из инфицированных и воспаленных тканей антигена в лимфатических узлах; антигены, проникшие в кровоток, задерживаются селезенкой. Во вторичных лимфоидных тканях происходит первоначальная клональная экспансия антигенспецифичных лимфоцитов, после чего они поступают в циркуляцию через выносящие лимфатические сосуды. Дальнейшая миграция их из кровотока зависит от экспрессии на клетках эндотелия молекул адгезии; например, при появлении таких молекул на эндотелии в очаге воспаления их распознают рецепторы активированных лимфоцитов или фагоцитов и в результате там и накапливаются эти клетки. Весь комплекс реакций, возникающих в тканях в ответ на повреждение или инфекцию, назван воспалением. Для воспаления характерны три основных признака:
• усиленное кровоснабжение воспаленной области, способствующее доставке в нее лейкоцитов и растворимых компонентов плазмы;
• повышенная проницаемость капилляров и вследствие этого экссудация белков плазмы, необходимых для сдерживания инфекции;
• усиленная миграция лейкоцитов.
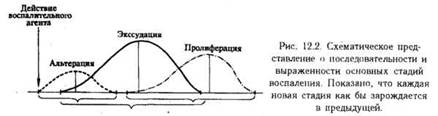
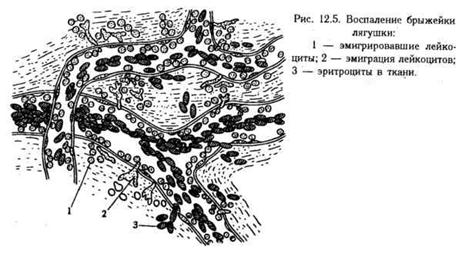
При иммунологической реакции на антиген различные популяции мигрирующих клеток появляются в ткани, как правило, поочередно. Тип клеток, присутствующих в каждый данный момент, преобладание тех или иных и время появления - все это зависит от природы антигена и от участка организма, где развертывается иммунологическая реакция. Обычно первыми в очаг острого воспаления, вызванного инфекцией, прибывают нейтрофилы, преобладающие затем в нем в течение нескольких суток. На вторые сутки в очаг начинают поступать мононуклеарные фагоциты и лимфоциты. Позже прибывают обычно Т-клетки CD8+ и немногочисленные В-клетки. Обратное развитие острой реакции зависит от того, удалось ли организму освободиться от антигена или инфекции. Если не удалось, острая воспалительная реакция переходит в хроническую, при которой в очаге мало нейтрофилов, но в значительном количестве накапливаются Т-клетки CD4+ и мононуклеарные фагоциты. Реакции на паразитарные инвазии нередко сопровождаются эозинофильной инфильтрацией. Эозинофилы вместе с базофилами и макрофагами преобладают также в инфильтрате тканей бронхиальной стенки после астматических приступов.
Миграция клеток
В миграции лейкоцитов выделяют две главные стадии. Первая - это прилипание циркулирующих клеток к сосудистому эндотелию с последующим проникновением между эндотелиоцитами или сквозь них. На второй стадии лейкоциты, преодолевшие эндотелий, мигрируют в направлении очага инфекции или воспаления, привлекаемые хемотаксическими стимулами. Эти процессы регулируются присутствующими на поверхности мигрирующих клеток белками, а также растворимыми сигнальными молекулами - хемокинами и другими хемоаттрактантами.
Пути клеточной миграции разнообразны и зависят не только от типа клетки, но и от стадии ее дифференцировки или уровня активации. Помимо этих факторов, существуют влияющие на миграцию особенности сосудистого эндотелия в разных участках организма. Так, венулы с высоким эндотелием, характерные для вторичной лимфоидной ткани, по строению совершенно отличны от венулнелимфоидных тканей. Эндотелий мелких сосудов в различных нелимфоидных тканях морфологически чрезвычайно разнообразен, и к тому же любая местная воспалительная реакция приводит к изменениям в экспрессии ряда его поверхностных молекул. От всех этих факторов зависит, клетки какого типа окажутся способными проникнуть сквозь эндотелиальную выстилку. Как правило, на миграцию лейкоцитов через эндотелий влияют 1) величина поверхностного заряда взаимодействующих клеток,
2) сила гемодинамического смыва в сосудистом русле и 3) экспрессия комплементарного набора молекул адгезии на поверхности как лейкоцитов, так и эндотелиальных клеток. Поэтому лейкоциты мигрируют из кровеносного русла через стенку именно венул, где поверхностный заряд эндотелиоцитов самый низкий, гемодинамический смыв незначителен а молекулы клеточной адгезии экспрессируются избирательно.
Лимфоцитарная миграция в лимфоидную ткань отличается от миграции в очаги воспаления
Для разных стадий жизненного цикла лимфоцитов характерны различные пути миграции. Например, неактивированные Т-клетки имеют тенденцию проникать через ВЭВ во вторичные лимфоидные ткани, тогда как активированные мигрируют в очаги воспаления. К тому же наблюдается избирательная миграция в определенные участки организма: лимфоциты, выделенные из пейеровых бляшек, при введении в кровь вновь локализуются в кишечнике, а лимфоциты селезенки возвращаются в нее же.
Миграция лимфоцитов из кровотока в лимфоузлы, пейеровы бляшки и лимфоидную ткань слизистых оболочек происходит через ВЭВ. Именно через стенки этих венул выходит из кровотока до 25% лимфоцитов, поступающих в лимфоузел по кровеносным сосудам. В противоположность этому через обычный эндотелий венул проникает при каждом цикле лишь ничтожная часть циркулирующих лимфоцитов. Тем не менее, эта слабая миграция крайне важна, так как позволяет лимфоцитам осуществлять "надзор" за всеми тканями организма; при развитии воспаления она многократно усиливается.
Венулы с высоким эндотелием выполняют, таким образом, особо важную роль в рециркуляции лимфоцитов. В норме ВЭВ присутствуют только во вторичных лимфоидных тканях, но могут возникать и в очагах хронического воспаления. Кроме характерной кубовидной формы, к особенностям эндотелиоцитов ВЭВ относится экспрессия различных наборов сульфатированных и обильно гликозилированных молекул межклеточной адгезии, которые связываются с циркулирующими Т-клетками, направляя их тем самым из кровотока в лимфоидную ткань. Эти молекулы адгезии отличаются от тех, которые регулируют миграцию лимфоцитов в очаг острого воспаления. При этом в разных лимфоидных тканях на эндотелии ВЭВ экспрессированы различные молекулы клеточной адгезии. Ранее эти молекулы адгезии были названы сосудистыми адрессинами; их экспрессия на эндотелии различных ВЭВ обеспечивает возвращение лимфоцитов в собственную лимфоидную ткань.
Миграцию регулирует ряд факторов, как эндотелиального, так и лейкоцитарного происхождения
Миграция лейкоцитов зависит от присутствия молекул адгезии на поверхности как эндотелия, так и лейкоцитов, от подвижности самих клеток и от наличия хемотаксических агентов. Для объяснения сложного и изменчивого характера клеточной миграции необходимо учитывать множество влияющих на нее факторов. К ним относятся:
• характер активации мигрирующих лимфоцитов или фагоцитов: экспрессия молекул адгезии и их функциональная аффинность варьируют в зависимости от типа клеток и от того, активированы они антигеном, цитокинами или межклеточными взаимодействиями;
• типы молекул адгезии, экспрессируемых сосудистым эндотелием: они определяются тем, в какой анатомической области проходит сосуд и был ли эндотелий активирован цитокинами;
• присутствие специфичных хемотаксических молекул и цитокинов в тканях: для разных популяций лейкоцитов характерны различные специализированные рецепторы, поэтому каждый хемотаксический агент избирательно привлекает только определенный тип клеток.
Прежде чем рассматривать участие молекул адгезии в лейкоцитарной миграции, необходимо познакомиться с их разнообразием и межклеточным распределением.
Молекулы межклеточной адгезииМолекулы межклеточной адгезии - это связанные с плазматической мембраной белки, которые обеспечивают механическое взаимодействие клеток друг с другом. Часто это молекулы, пронизывающие мембрану и присоединенные к цитоскелету; с их помощью клетки при движении могут подтягиваться к другим клеткам или перемещаться по внеклеточному матриксу. Во многих случаях отдельная молекула межклеточной адгезии способна взаимодействовать не с одним, а с несколькими лигандами, для чего служат разные, участки связывания. Хотя связывание индивидуальных молекул адгезии со своими лигандами обычно происходит с низким сродством, авидность взаимодействия может быть довольно высокой, за счет того что молекулы адгезии расположены на поверхности клеток компактными "пятнами", или кластерами, и образуют участки многоточечного связывания.
Адгезия клеток одного типа к клеткам другого типа может изменяться в результате увеличения числа молекул адгезии на клеточной поверхности либо при изменении их аффинности и/или авидности. Существуют два механизма увеличения числа молекул адгезии на поверхности клеток: у многих клеток большие запасы этих молекул хранятся во внутриклеточных везикулах, которые способны через несколько минут после активации устремляться к поверхности цитоплазматической мембраны; другой механизм состоит в синтезе таких молекул de novo и переносе их на поверхность.
Число известных молекул межклеточной адгезии, имеющих значение для лейкоцитарной миграции, ошеломляюще велико. Тем не менее, все их можно распределить по четырем семействам структурно родственных молекул. Миграция клеток - это сложный процесс, в котором на разных стадиях принимают участие несколько наборов молекул адгезии.
Некоторые эндотелиальные молекулы адгезии относятся к иммуноглобулиновому суперсемейству.
К суперсемейству иммуноглобулинов принадлежит ряд молекул адгезии эндотелиальных клеток, в том числе молекула межклеточной адгезии 1 типа и 2 типа, молекула адгезии сосудистого эндотелия I типа и молекула адгезии ВЭВ лимфоидных образований в слизистых оболочках, названная ранее адрессином I типа - MAdCAM-l. Экспрессия этих молекул на эндотелии сосудов может быть постоянной или индуцибельной. ICAM-1 имеет пять внеклеточных доменов, из которых два Н-концевых структурно гомологичны двум внеклеточным доменам ICAM-2. В составе VCAM-1 шесть внеклеточных доменов; эти же данные присутствуют в структуре MAdCAM-l.
В адгезии лейкоцитов к эндотелию и внеклеточному матриксу участвуют интегрины.
Интегрины - это обширная группа молекул межклеточной адгезии, которые присутствуют на поверхности различных клеток, в том числе лейкоцитов. Все белки, входящие в это крупное семейство, состоят из двух нековалентно связанных полипептидных цепей; обе цепи пронизывают клеточную мембрану. Семейство интегринов делят на три основные подсемейства по типу в-цепи. Тип б-цепи, соединенной с в-цепью, оказался не так важен для функциональной активности, как предполагали первоначально. В основном в-интегрины участвуют в связывании клеток со структурами внеклеточного матрикса, в2-33феГсМ33З - в адгезии лейкоцитов к эндотелию или другим клеткам иммунной системы, а в^-ЗЗфефсЗЗх - в слипании тромбоцитов с нейтрофилами в очагах воспаления или местах повреждения сосудов. Из этой простой схемы имеется, однако, несколько исключений и, кроме того, описаны дополнительные в-цепи. Каждая в-цепь может ассоциировать с одной из различных а-цепей, в результате чего образуются разнообразные молекулы адгезии. Способность интегринов связываться со своими лигандами зависит от двухвалентных катионов. Например, LFA-I в норме экспрессирован на лейкоцитах в низкоаффинном состоянии, но при повышении концентрации Mg2+ становится высокоаффинным. При повышении концентрации Са2+ интегрины на поверхности клеток перераспределяются, формируя высокоавидные "пятна".
Селетины - группа лейкоцитарных и эндотелиальных молекул межклеточной адгезии, которые связываются с углеводами
Группа селектинов включает Е-селектин, Р-селектин и L-селектин, экспрессируемые на эндотелии, тромбоцитах и некоторых типах лейкоцитов соответственно. Молекулы селектинов пронизывают мембрану и имеют ряд внеклеточных доменов, гомологичных доменам регуляторных белков комплемента, например фактору Н.
Внеклеточная часть их молекул содержит также домен, близкий по структуре рецептору для фактора роста эпидермиса, и Н-концевой домен, обладающий лектиноподобными свойствами, т.е. связывающий углеводные остатки. Соответственно этому в состав лигандов, с которыми связываются селектины, входят углеводы.
Лиганды селектинов - это углеводные компоненты различных гликопротеинов лимфоцитарной и эндотелиальной поверхности
Углеводные лиганды для селектинов встречаются в структуре разнообразных белков. Так, многочисленные О-связанные углеводные остатки, которые служат лигандами для лимфоцитарного L-селектина, содержит гликопротеин Sgp-200, экспрессируемый на эндотелии ВЭВ. L-селектины могут также связываться с углеводами в составе MAdCAM-l, который экспрессирован на эдотелии ВЭВ лимфоидных образований в слизистых оболочках. Эти гликопротеины, названные в соответствии с их функцией сосудистыми адрессинами, присутствуют главным образом на клетках ВЭВ в лимфоидных тканях, но могут быть индуцированы при хроническом воспалении и в других участках организма.
Е - и Р-селектины, экспресспруемые на активированных эндотелиоцитах, связываются с сиалированной формой углеводной детерминанты Lex, ассоциированной с белком CD 15, который присутствует на лейкоцитах многих субпопуляций. В результате этого связывания движение лейкоцитов с током крови прекращается н наступает первая фаза миграции.
Механизмы клеточной миграции
Выход лейкоцитов из сосудистою русла через эндотелий происходит в несколько этапов.
• Краевое стояние: движение лейкоцитов по венуле замедляется и они катятся по эндотелию. Это обусловлено в основном взаимодействием селектинов с углеводными лигандами.
• Активация: остановленные лейкоциты подвергаются воздействию цитокинов, хемотаксических агентов, компонентов поверхности эндотелия и внеклеточною матрикса.
Миграция лейкоцитов к очагу воспаления Нейтрофилы появляются в очаге острого воспаления на его ранней стадии, и отчасти это обусловлено индукцией цитокинами экспрессии Е-селектина на поверхности эндотелия в этой области. Стимуляция клеток эндотелия in vitro такими цитокинами, как, например, фактор некроза опухолей б или интерлейкин I индуцирует Экспрессию Е-селектина спустя 4-12 ч, а через 24 ч она прекращается; аналогичным образом in vivo эта молекула межклеточной адгезии появляется на ранней стадии воспалительной реакции. Клетки, трансфицированные геном Е-селектина, экспрессируют его в значительном количестве, приобретая при этом выраженную способность связывать нейтрофилы. Все эти данные указывают, что остановка нейтрофилов при участии Е-селектина - это первая необходимая стадия их миграции.
Важную роль в миграции нейтрофилов, лимфоцитов и моноцитов выполняют также экспрессируемые на лейкоцитах Р2-интегрины LFA-I и CR3, которые связываются с эндотелиальными молекулами межклеточной адгезии из суперсемейства иммуноглобулинов. Так, LFA-1 связывается с [САМ-1 и [САМ-2 на эндотелии сосудов.
В результате прикрепления лейкоцитов и мобилизации интегринов повышается аффинность лейкоцитарных интегринов, которые взаимодействуют с эндотелиальными молекулами межклеточной адгезии и активируют клетку для миграции.
Миграция: при участии нового набора молекул адгезии лейкоциты связываются с базальной мембраной эндотелия и проникают сквозь нее.
Лизис: мигрирующие клетки выделяют ферменты, лизирующие коллаген и другие компоненты базальной мембраны эндотелия; это позволяет лейкоцитам проникнуть в ткань.
Молекулы межклеточной адгезии регулируют прилипание лейкоцитов и прохождение их через эндотелий
Для каждого типа перемещения клеток характерно участие специфического набора молекул адгезии и хемотаксических агентов.
культуре клетки эндотелия конститутивно синтезируют 1САМ-2; в связи с этим высказано предположение, что именно данный белок определяет фоновый уровень связывания лимфоцитов с эндотелием различных типов in vivo. Например, уровень экспрессии 1САМ-2 на эндотелии мозговых сосудов в норме относительно низок и этому соответствует весьма незначительная трансэндотелиальная миграция лимфоцитов. Напротив, экспрессия 1САМ-1, в норме низкая на поверхности эндотелия, может быть резко повышена цитокинами. В условиях in vitro индуцированная экспрессия ICAM-1 наблюдается в период 8-96 ч после стимуляции, что соответствует более позднему прибытию в очаг воспаления in vivo лимфоцитов и моноцитов. Роль CR3 в привлечении фагоцитов показали опыты in vivo с использованием антител анти-СРчЗ, которые, как было при этом установлено, подавляют миграцию данных клеток. У больных с дефицитом лейкоцитарной адгезии, подверженных в результате слабого накопления фагоцитов тяжелым инфекционным заболеваниям, отмечена недостаточность всех р2-интег-ринов. Следует отметить, что CR3 и LFA-1 связываются с разными участками 1САМ-1.
Нормальная миграция лейкоцитов Описанные выше молекулы, регулирующие миграцию клеток в область воспаления, необходимо отличать от молекул, ответственных за нормальную миграцию лимфоцитов. Непримированные лимфоциты экспрессируют L-селектин, обеспечивающий связывание с углеводными лигандами на эндотелии ВЭВ в лимфоидных образованиях слизистых оболочек и в периферических лимфоузлах. Останавливаясь, например, в пейеровой бляшке, они могут связываться при участии интегрина б4в7 с MAdCAM-l на поверхности эндотелия. Поскольку интегрин б4в7 обеспечивает миграцию лимфоцитов в лимфоидную ткань слизистых оболочек, а интегрин б4в - связывание с VCAM-1 на активированном эндотелии или с фибронектином во внеклеточном матрик-се, экспрессия той или другой из этих молекул означает различное направление миграции - непримированных лимфоцитов в нормальную лимфоидную ткань или активированных Т-клеток в очаг воспаления.
Взаимодействие лейкоцитов с внеклеточным матриксом Пройдя сквозь эндотелий сосуда и проникнув в ткань, лейкоциты неминуемо должны взаимодействовать с белками внеклеточного матрикса, а также с клетками ткани. Покинувшие кровеносный сосуд лимфоциты сразу теряют в результате энзиматического расщепления уже ненужный им L-селектин. Происходит смена функционального фенотипа: циркулировавшая клетка становится приспособленной к перемещению в тканях.
Многие из молекул лейкоцитарной поверхности, обеспечивающих взаимодействие с внеклеточным матриксом, входят в группу вй-интегринов; они названы "очень поздними антигенами", поскольку впервые были идентифицированы на поверхности Т-клеток в поздней стадии активации. Теперь все вй-интегрины называют VLA, хотя большинство их присутствует не только на лимфоцитах. Эта группа включает рецепторы для коллагена, ламинина и фибронектина. Сам факт появления некоторых из этих молекул, означающий отдаленный результат активации лимфоцитов, свидетельствует о выполнении клетками некой программы днфференцировки, в которой взаимодействие с внеклеточным матриксом - это один из наиболее поздних этапов.
Хемотаксические молекулы стимулируют лейкоциты к миграции и определяют ее направление
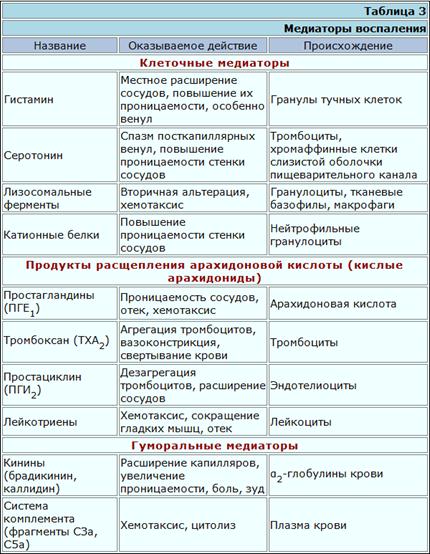
Интегрины, с помощью которых лейкоциты проникают сквозь эндотелий, присутствуют на клеточной поверхности или, прежде чем попасть на нее, сохраняются во внутриклеточных гранулах; при этом большинство из них неактивно и для функционирования нуждается в исходящем от эндотелия сигнале активации. Сигнальные молекулы могут быть продуктами самого эндотелия или осевшими на нем пептидами, которые выделяет подлежащая ткань. Многие из сигнальных молекул обладают также хемотаксическими свойствами; в том числе это С5а, лейкотриен-В4 и разнообразные низкомолекулярные цитокины, получившие общее название "хемокины".
Хемотаксическую активность клеток следует отличать от хемокинетическон: хемотаксис - это направленная миграция клеток по градиенту концентрации хемотаксических молекул, а хемокинез - беспорядочное перемещение клеток. В основе направленной миграции лежит способность клетки отвечать на градиент концентрации хемотаксического медиатора при величине этого градиента между ее ведущим и концевым полюсами не менее 0,1%. Хсмокинез связан с усилением общей подвижности клеток под действием того или иного медиатора, например гистамина.
Хемокины Это группа хемотаксических гепарин-связывающих молекул, в которую входят не менее 25 низкомолекулярных цитокинов, в частности ИЛ-8 и RANTES. Хемокины высвобождаются в очаге воспаления и могут связываться на поверхности эндотелия, взаимодействуя с сульфатными группами присутствующего на нем гепарина. Многие из этих цитокинов связывает также антиген DARC - групповой антиген крови системы Даффи. избирательно экспрессируемый на эндотелии венул. Связанные с поверхностью эндотелия хемокины могут вызывать повышение авидности интегринов на лейкоцитах в фазе "краевого стояния", остановленных при участии селектинов. Большинство хемокинов синтезируется лейкоцитами, однако ИЛ-8 и хемотаксический для макрофагов белок 1 продуцирует, например, культура клеток эндотелия, причем активация этих клеток цитокинами, способствующими развитию воспаления, усиливает синтез. Хемокины и другие хемотаксические молекулы, рассматриваемые ниже, воздействуют на клетки посредством "змеевидных" рецепторов, обладающих семью трансмембранными сегментами. Различные змеевидные рецепторы избирательно распределены среди отдельных популяций лейкоцитов, чем отчасти можно объяснить избирательность действия различных хемокинов, например тот факт, что макрофагальный воспалительный белок 1в. Эти клетки имеют рецепторы для пептидов, блокированных на Н-конце формилметионнном, в частности рецептор, связывающий трипептид. Поскольку при трансляции всех белков у прокариот в отличие от эукариот иниииаторной аминокислотой служит метионин, он и выполняет роль простого специфического сигнала присутствия бактерий, по направлению к которым должны устремиться фагоциты. Нейтрофилы и макрофаги имеют также рецепторы для С5а и лейкотриена В4. Оба эти хемоаттрактанта образуются в очаге воспаления: С5а - в результате активации комплемента, лейкотриен LTB4 - при активации разнообразных клеток, чаше всего макрофагов и тучных. Кроме того, хемотаксис фагоцитов вызывают молекулы, образуемые системой свертывания крови, прежде всего фибриновый пептид В и тромбин.
Клетки, прибывшие в очаг воспаления первыми, способны в результате активации вызвать следующую волну лейкоцитарной миграции. Так, активированные моноциты выделяют ИЛ-8, который может вызвать хемотаксис нейтрофилов и базофилов. Подобно этому, активация макрофагов приводит к метаболизированию арахидоновой кислоты с образованием и выделением лейкотриена В4.
Воспаление
Воспаление - это реакция организма на внедрение инфицирующего агента, введение антигена или физическое повреждение тканей. Помимо усиления клеточной миграции, описанного выше, воспаление вызывает приток различных растворимых молекул из плазмы крови. В противоположность лейкоцитам, которые мигрируют через эндотелий венул, молекулы плазмы крови попадают в воспалительный экссудат главным образом из капилляров, где кровяное давление выше. Этот процесс обеспечивается двумя механизмами:
• усилением кровенаполнения капилляров в области воспаления и
• увеличением проницаемости капилляров. Проницаемость капилляров повышается вследствие втягивания клеток эндотелия и, возможно, также усиления транспорта везикул сквозь эндотелий. Это обеспечивает поступление в очаг воспаления более крупных молекул, чем те, которые обычно могут проникать сквозь эпителий. Таким образом в очаг воспаления поступают антитела, компоненты комплемента и другие ферментные системы плазмы крови.
Воспаление регулируется хемокинами, ферментными системами плазмы, цитокинами, а также продуктами метаболизма тучных клеток, тромбоцитов и лейкоцитов
Развитие воспалительного процесса происходит при участии 1) хемокинов,
2) продуктов активации ферментных систем плазмы и 3) вазоактивных медиаторов, выделяемых лейкоцитами. Воспалительные реакции разного типа регулируются различными медиаторами. Немедленный ответ зависит от быстродействующих вазоактивных аминов и продуктов кининовой системы. Позднее привлечение и активация лейкоцитов происходят под действием вновь синтезированных медиаторов, таких как лейкотриены.
Достигая очага инфекции или воспаления, лейкоциты ранней волны миграции выделяют медиаторы, которые обеспечивают дальнейшее накопление и активацию клеток. Однако роль главного регулятора воспалительных реакций, инициированных иммунной системой, как и иммунного ответа вообще, принадлежит самому антигену. Поэтому очаг хронической инфекции или аутоиммунных реакций существенно отличается по клеточному составу инфильтрата от очагов воспаления, быстро освобождаемых от антигена.
Ферментные системы плазмы Существенная роль в гемостазе и регуляции воспаления принадлежит четырем главным ферментным системам плазмы крови: системе свертывания, системе фибринолиза, системе кининов и системе комплемента. Система комплемента опосредует многообразные взаимодействия между иммунным ответом и воспалением. К кининовой системе относятся медиаторы брадикинин и лизилбрадикинин. Брадикинин - это функционально весьма сильный вазоактивный нонапептид, вызывающий увеличение просвета венул и сосудистой проницаемости, а также сокращение гладких мышц. Он образуется в результате активации фактора Хагемана, относящегося к системе свертывания крови, тогда как для образования каллидина необходимы активация плазминовой системы или участие ферментов, выделяемых поврежденными тканями.
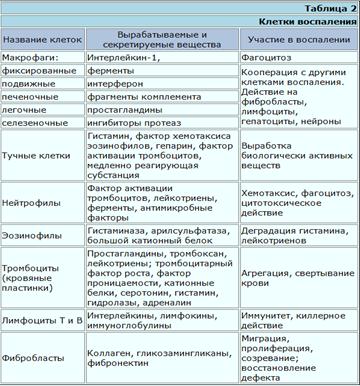
Вспомогательные клетки воспаления К ним относятся тучные клетки, базофилы и тромбоциты; все эти клетки служат важным источником вазоактивных медиаторов - гистамина и 5-гидрокситриптамина, вызывающих вазодилатацию и увеличение проницаемости сосудов. Многие из провоспалительных эффектов СЗа и С5а обусловлены их способностью вызывать высвобождение содержимого гранул из тучных клеток. Об этом свидетельствует факт подавления данных эффектов антигистаминными препаратами. Кроме того, тучные клетки и базофилы могут стать непосредственной причиной воспаления, вызванного специфическим иммунным ответом, так как IgE сенсибилизирует их для дегрануляции при встрече с антигеном. Тучные клетки служат также важным источником медленнореагирующих медиаторов воспаления, в том числе лейкотриенов, простагландинов и тромбоксанов.
Тромбоциты, как и тучные клетки, могут быть активированы продуктами иммунной системы - иммунными комплексами или фактором активации тромбоцитов, выделяемым нейтрофилами, базофилами и макрофагами. Предполагается, что этот механизм важен в реакциях гиперчувствительности II и III типов.
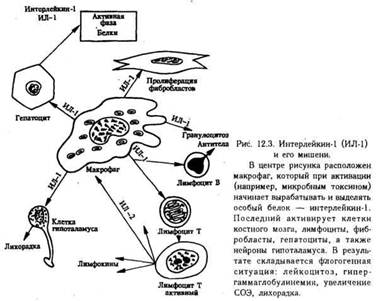
ЦитокиныПодобно другим медиаторам, цитокины служат для межклеточной сигнализации при развитии воспалительного процесса. На его начальных стадиях местные тканевые клетки могут выделять такие цитокины, как ИЛ-1 и ИЛ-6. Как только в очаге воспаления появляются лимфоциты и мононуклеарные фагоциты, они могут, активируясь под действием антигена, выделять свои собственные цитокины, которые, воздействуя на эндотелий местных сосудов, дополнительно усиливают клеточную миграцию. Другие цитокины, например ИЛ-8, могут оказывать хемотаксическое или активирующее действие на прибывающие клетки.
Похожие работы
... кПа (8—11 атм) при воспалении (Шаде). Ацидоз обусловливает набухание элементов соединительной ткани. Повышение осмотического давления усиливает экссудацию и местный отек. Этим объясняются главные признаки воспаления — появление припухлости и боли, наличие которой тоже в значительной степени объясняется натяжением ткани, возникающим при припухлости. Пролиферация и завершение процесса. На этой ...
... - в 14,7 раза, адгезия моноцитов к эндотелиальному монослою возросла на 48 % [163]. Одним из важнейших факторов, запускающим при воспалении трансэндотелиальную миграцию моноцитов, является активирующее действие нейтрофилов на клетки эндотелия. В результате этого стимулируется продукция и высвобождение эндоте-лиоцитами МСР-1 и повышается хемотаксическая активность моноцитов, что способствует их ...
... зависит от вида и степени гормонального дисбаланса. Страдают в начале клетки-мишени, ферментные системы которых находятся под регулирующим действием соответствующих гормонов. При рассмотрении патофизиологии острого повреждния клетки следует отдельно остановиться на роли лизосомального аппарата. Существует много причин, приводящих к недостаточности функции лизо- - 23 -сом: угнетение ...
... агглютинация тромбоцитов изменения сосудистой стенки, снижение тромборезистентности сосудистой стенки. В эндотелии сосудов синтезируется простациклин, который предотвращает адгезию и агрегацию тромбоцитов. В воспалении происходит повреждение сосудистой стенки и количество простациклина уменьшается, начинаются процессы адгезии и агглютинации тромбоцитов. Из тромбоцитов выделяются тромбоксаны, ...
0 комментариев