Реферат на тему:
Разделение белков по размерам с использованием DDC-Na
2009
Разделение белков по размерам с использованием DDC-Na
Электрофорез белков в простой системе удобно использовать для их разделения, но не характеристики. Электрофоретическая подвижность каждого белка в простой системе зависит одновременно и от массы белка, и от его суммарного электрического заряда, и от конфигурации, и от жесткости упаковки белковой глобулы. Вклад каждого из этих факторов неизвестен и может существенно изменяться в зависимости от условий электрофореза. Для установления строгой корреляции между каким-либо одним из перечисленных параметров и электрофоретической подвижностью белка надо исключить влияние всех остальных.
Электрофорез в ПААГ с использованием DDC-Na позволяет разделять белки, различающиеся между собой только по молекулярной массе. Для этого смесь белков в исходном препарате обрабатывают не менее, чем трехкратным избытком DDC-Na (по весу). За счет гидрофобных взаимодействий детергент одинаково связывается с подавляющим большинством белков в соотношении 1,4 мг DDC-Na на 1 мг белка.
Огромный избыток полностью диссоциированных остатков сульфокислоты, привносимый детергентом, в большинстве случаев делает несущественной роль собственного заряда белка. Благодаря электростатическому отталкиванию тесно расположенных отрицательно заряженных остатков серной кислоты, белковая цепь распрямляется и приобретает форму жесткой палочки с поперечником -1,6 m и длиной, зависящей только от числа звеньев этой цепи, а следовательно — от молекулярной массы белка.
Одновременно с обработкой DDC-Na необходимо обеспечить возможность полного развертывания белковой цепи, а для этого — разорвать все ковалентные S-S связи внутри молекулы белка. С этой целью белок перед электрофорезом обрабатывают еще и высокой концентрацией (1%) (3-меркаптоэтанола при повышенной температуре.
Электрофретическая подвижность, т. е. скорость миграции при напряженности поля 1 В/см, жесткого комплекса белок DDC-Na оказывается связанной с молекулярной массой белка (М) простым соотношением A-BIgM, где А и В — коэффициенты, зависящие от пористости геля, температуры и других условий эксперимента. Величину и' удобнее представлять в относительных единицах, выражающих отношение путей миграции белка и «лидирующего красителя» — бромфенолового синего. Обозначим это отношение Rf. Такая замена отразится только на значении коэффициентов А и В, о которых нам известно только то, что они в данных условиях опыта одинаковы для всех белков. Измененные величины коэффициентов тоже будут постоянными величинами в данном опыте. Поэтому вместо их определения пользуются методом сравнения с известными по своей массе «маркерами». Одновременно с электрофорезом изучаемой смеси белков, отдельным треком на той же пластинке, т.е. в тождественных условиях эксперимента, разделяют смесь известных маркеров. С их помощью по точкам строят зависимость lgM=f(Rf), которая, естественно, оказывается прямолинейной. Опираясь на эту зависимость, можно графически, по измеренным величинам Rf определить значения lg М, а следовательно и М для исследуемого белкА. Разумеется, вся предварительная обработка детергентом и Р-меркаптоэтанолом должна быть проведена строго одинаково для этого белка и всех маркеров.
Выбор буфера в этом варианте не играет роли, так как заряд белка определяется его комплексом с DDC-Na. Обычно используют нейтральный буфер, добавляя в него 0,1% DDC-Na, чтобы поддержать комплекс детергента с белками»
Для белков с молекулярной массой менее 12 тысяч дальтон определение М становится ненадежным. Для различных диапазонов масс белков рекомендуется использовать ПААГ различной пористости, согласно следующей таблице:
Диапазон М (тысяч дальтон) % ПААГ 12-43 15 16-68 10 36-94 7,5 57-212 5
Сам процесс электрофореза и окраски белков после его окончания производят как обычно, но DDC-Na от белка предпочтительно отмыть до окраски, вымачивая гель в 50% -ной ТХУ в течение ночи.
DDC-Na в комплексе с белком в некоторой мере препятствует окрашиванию (большой отрицательный заряд!).
Двумерный электрофорез в ПААГ
Полное разделение сложной смеси белков не всегда удается осуществить в ходе одного электрофоретического эксперимента. Всегда есть вероятность того, что в данной системе электрофореза различные белки мигрируют в одной зоне либо в силу близости их размеров, либо ввиду совпадения их электрофоретических подвижностей при выбранном значении рН, либо, наконец, в результате неблагоприятной для разделения комбинации этих параметров. Поэтому в сложных случаях фракционирования смеси белков имеет смысл использовать разделение в первом направлении как исходное для разделения во втором, перпендикулярном первому направлении при измененных условиях электрофореза.
Для этого трек первого направления (без осаждения и окраски белков в нем) вырезают и накладывают на стартовую зону пластинки второго направления (без карманов). Контакт между двумя гелями обеспечивают заливая место их соприкосновения расплавленным раствором агарозы в том же буфере. Электрофорез, естественно, ведут в направлении, перпендикулярном полоске. Каждая негомогенная полоса в ней может дать несколько пятен во втором направлении, если в новых условиях содержавшиеся в ней белки обретут различную электрофоретическую подвижность. В результате после осаждения и прокрашивания на пластинке появляется картина распределенных по всей поверхности пятен («фингерпринт»). Число пятен различных белков, которое удается зафиксировать на одной пластине может достигнуть нескольких сотен. На рис. 42 воспроизведена картина распределения пятен, полученных при двумерном электрофорезе белков из большой субъединицы одной из бактерий (в работе Mets, Bogorad Anal. Biochem. 57 200, 1974).
Извлечение белков из геля после электрофореза
Для целей аналитической идентификации белки из ПААГ (без осаждения и окраски) можно перенести на нитроцеллюлозный мембранный фильтр. Такие фильтры обладают способностью сорбировать основные белки. Перенос осуществляется путем вымывания белковых полос из геля током буфера в направлении, перпендикулярном поверхности пластинки. Фильтр накладывают непосредственно на влажный гель. Устройство для обеспечения тока буфера от геля к фильтру будет описано ниже в связи с электрофорезом ДНК.
В результате на фильтре получаются «реплики» белков, разделенных в геле. Для их идентификации можно использовать характерные реакции, иногда ферментативные или имунные (см. ниже), а также гибридизацию с мечеными радиоактивным фосфором ДНК или РНК, если эти белки in vivo связывались с ДНК или разделению подвергались рибосомальные белки, связывающиеся с РНК рибосом.
Электрофорез нуклеиновых кислот
Главное, что с точки зрения электрофореза отличает нуклеиновые кислоты от белков — это значительный по величине суммарный отрицательный заряд, обусловленный диссоциацией многочисленных остатков фосфорной кислоты в связях между нуклеозидами. рН окружающей среды мало влияет на этот заряд. Поэтому электрофорез можно вести не в буфере, а в любом подходящем ионосодержащем растворе, например в слабом растворе щелочи. Во всех случаях в жидкую среду вносят ЭДТА до концентрации 1—2 mM. Это необходимо для блокирования действия нуклеаз и предупреждения осаждения (особенно РНК) двухвалентными металлами.
Таким образом, разделение фрагментов ДНК и РНК электрофорезом приходится вести только по размеру. Однако размеры эти могут варьировать в очень широких пределах: от десятков нукле-отидных звеньев до многих сотен тысяч, а если выражать через молекулярные массы, то от нескольких тысяч до сотен миллионов дальтон.
Естественно, что для фракционирования относительно коротких фрагментов используют электрофорез в ПААГ, а для разделения высокомолекулярных ДНК, более крупнопористый носитель — агарозу. С ней мы познакомимся чуть позже. А пока в порядке связи с предыдущими параграфами уместно сделать несколько замечаний об электрофорезе ДНК в ПААГ. В нижеследующей таблице представлены рекомендации по выбору пористости геля в зависимости от размеров относительно коротких фрагментов ДНК, охарактеризованных числом пар оснований (п.о.):
| Диапазон п.о. (штук) | 6-100 | 25-150 | 40-200 | 60-400 | 80-500 | 1-2 тыс. |
| % ПААГ | 20 | 15 | 12 | 8 | 5 | 3,5 |
В последнем диапазоне этой таблицы находятся предельные размеры ДНК, доступные автоматическому секвенированию последовательности нуклеотидов. Этот революционный метод анализа ДНК будет рассмотрен в следующей главе.
Сейчас же, прежде, чем перейти к электрофорезу в агарозе, я хочу познакомить учащихся и читателей с новыми идеями фракционирования крупных фрагментов ДНК в ПААГ с оригинальным использованием импульсов электрического напряжения, подаваемых на гель. Эти импульсы подаются поочередно в двух взаимно перпендикулярных направлениях. Идея здесь заключается вот в чем. Даже очень длинная и гибкая молекула ДНК может протиснуться через относительно малые поры геля, если она вытянута в направлении продвижения к «основному» аноду, скажем, расположенному внизу пластинки. Напряжение на этот анод подается не постоянно, а импульсами. Второй, «вспомогательный» анод создает напряженность электрического поля, перпендикулярную основному направлению движения ДНК к низу пластинки. Напряжение на него подается тоже достаточно мощными, но более короткими импульсами, чем на основной анод, чередуясь с ними.
Назначение «перпендикулярного» электрического поля состоит в том, чтобы поворачивать длинные молекулы больших ДНК так, чтобы в момент подачи «продольного» импульса они находились в положении, благоприятном для движения вдоль пластинки вниз, к основному аноду. Чем длиннее цепи ДНК, тем медленнее они будут менять свою ориентировку и потому медленнее подвигаться в нужном направлении. Авторы метода утверждают, что таким образом в ПААГ им удавалось разделять фрагменты ДНК длиной до пяти миллионов пар оснований.
Предложено несколько вариантов использования этой идеи. В одном из них чередующиеся импульсы одинаковой силы направлены каждый под углом 45° к продольной оси пластины, навязывая таким образом молекулам ДНК движение зигзагом, своего рода «рыскание». (Идея, по всей видимости, почерпнута из практики проталкивания через густую толпу людей.)
Гели агарозы
Агароза — это особо чистая фракция природного, линейного полисахарида, агара, выделяемого из морских водорослей. Мы с ним уже встречались.
Молекулярная масса одиночных нитей агарозы лежит в пределах 10-100 тысяч дальтон. Агароза для электрофореза поставляется в виде лиофилизированного (высушенного в вакууме) порошка. Гелеобразование происходит при остывании даже очень разбавленного горячего раствора агарозы в буфере. При температуре 84-96° С (а у некоторых типов агарозы уже при 70°) нити полимера плавятся и образуют с окружающей водной средой однородную прозрачную жидкость. Она обладает ярко выраженным температурным гистерезисом — застывает при температуре порядка 40°С. Остывая до этой температуры, даже 0,4% -ные растворы агарозы образуют прочные гели. У легкоплавких типов агарозы температура затвердевания снижается до 30°. Такая особенность агарозы облегчает все манипуляции с ее растворами, не опасаясь их затвердевания. Более того,, расплавленную на кипящей бане взвесь агарозы в воде охлаждают до 50-55° и только при этой температуре добавляют концентрат буфера и все другие добавки, а затем заливают в форму для электрофореза. Это удобно и не связано с возникновением тепловых деформаций.
Остывая, хаотически ориентированные нити затвердевшей агарозы благодаря множественным водородным связям между нитями собираются в жгуты. Эти жгуты свободно перекрещиваясь, создают в окружающей их жидкости очень крупнопористую и, вместе с тем, жесткую пространственную сетку.
Размер пор геля агарозы, естественно, связан с концентрацией ее исходного раствора. Можно привести некоторые рекомендации по выбору этой концентрации, почерпнутые из опыта электрофореза нуклеиновых кислот:
Для нуклеиновых кислот вирусов и крупных плазмид 0,4-0,5%.
Для рестриктов ДНК, содержащих 5—20 тысяч пар оснований 0,7-0,8%.
Для более коротких рестриктов ДНК и двунитевых РНК рео-вируса 1,5%.
Для рибосомальных РНК — 1,75%.
Для иРНК и рестриктов ДНК до 1000 п.о. — 2% .
Приготовление пластины для электрофореза в агарозе очень просто. Нужного размера стекло оклеивают со всех сторон по ребру сплошной липкой лентой так, чтобы ее верхний край на несколько миллиметров выступал над поверхностью стекла. Последнее кладут на строго горизонтальный столик. Прямо на стекло выливают рассчитанный объем еще жидкого раствора агарозы. Затем в него близ одного края стекла устанавливают гребенку, подобную той, которую ставят в форму ПААГ перед его полимеризацией. Только на этот раз гребенку погружают в агарозу перпендикулярно стеклу (зубцы ее, однако, не должны касаться стекла). После застывания агарозы гребенку вынимают, образуя таким образом «колодцы» для препаратов. Электрофорез ведут в горизонтальном положении (в треках), накладывая фитили, идущие от резервуаров с буферами. Рабочее значение напряженности поля 2-5 В/см.
ДНК, особенно двунитевые, а также и РНК хорошо окрашиваются желтым флюоресцентным красителем — бромистым эти-дием. Этот краситель несет положительный заряд, что позволяет ему взаимодействовать с фосфатными группами нуклеиновых кислот. Кроме того он способен интеркалировать (встраиваться) между оснований двунитевой ДНК, что приводит к резкому усилению его флюоресценции при освещении ультрафиолетовым светом. Окраску можно производить и после электрофореза вымачиванием геля в течение 0,5—1 часа в водном растворе красителя (1 мкг/мл) или же вводить его непосредственно в гель. В последнем случае можно следить за движением полос в геле. Стеклянную пластину в этом случае освещают УФ-лампой снизу. Чувствительность окраски высока. Можно наблюдать и фотографировать полосы, содержащие 0,01 мкгДНК.
В конце электрофореза ДНК можно зафиксировать в геле осаждением из раствора, вымачивая гель в 70% -ном этаноле.
Пипетки
Конечно, современные микропипетки совсем не похожи на те, которые употребляют в школьном химическом кабинете. Их устройство таково. Довольно объемистую, пластмассовую (снаружи) рукоятку пипетки удобно держать четырьмя пальцами руки, оставив большой палец свободным для нажатия кнопки, торчащей из верхнего конца рукоятки. Книзу из нее выходит длинный, слегка конический металлический стержень, на конец которого плотно надевается конический полый наконечник из специальной пластмассы.
В рукоятке спрятан механизм, главную часть которого составляют очень точно подогнанные друг к другу тонкий цилиндр и поршень, отжимаемый кверху пружиной. Нажатием кнопки поршень перемещается вниз. Опустив наконечник в жидкость и освободив предварительно нажатую кнопку набирают в наконечник объем в 1, 2, 3 и более микролитров — в соответствии с калибровкой пипетки. Вторичным нажатием кнопки жидкость из наконечника полностью выталкивается. Наконечники поставляются стерилизованными и используются однократно.
Есть пипетки с возможностью предварительной регулировки объема жидкости, например, от 2-х до 20-ти микролитров. Регулировку осуществляют поворотом головки винта, который устанавливает длину хода поршня. Выбранный объем указывает связанный с винтом градуированный барабанчик.
Влияние вторичной структуры ДНК
В электрофоретическом поведении однонитевых (денатурированная ДНК, РНК) и двунитевых молекул нуклеиновых кислот многое определяется их размерами. В случае коротких полинуклеотидных цепей их нативная двунитевая молекула имеет более жесткую структуру, чем таких же размеров однонитевая молекула. Она труднее изгибается, проходя через пространственную сетку геля. В силу этого относительно короткие двунитевые фрагменты ДНК будут отставать при электрофорезе в ПААГ от денатурированных ДНК такой же длины. Такая ситуация будет иметь место даже для ДНК бактериофага ФХ-174 с молекулярной массой в 3,5 миллиона дальтон. Однако для более крупных молекул ситуация может измениться на противоположную. Длинная двунитевая цепочка оказывается уже достаточно гибкой: она продвигается через поры геля как бы «извиваясь ужом». Между тем однонитевая цепь той же длины сворачивается в рыхлый «хаотический клубок» такого размера, что его продвижение в геле оказывается более затрудненным. Денатурированная ДНК в этом случае при электрофорезе отстает от нативной. Естественно, что граница обращения описанного эффекта зависит от размеров пор геля.
Вирусные и митохондриальные двунитевые ДНК, а также плазмиды бактерий имеют структуру замкнутого двунитевого кольца. Нативное состояние такого кольца — «сверхскрученное» (ему отвечает минимум внутренних напряжений). Кольцо в целом сворачивается в «жгут», что сильно увеличивает его компактность (форма I). Если же хотя бы в одной из двух скрученных нитей кольца появляется единичный разрыв сахаро-фосфатной цепи, то жгут разворачивается и под действием сил электростатистического отталкивания фосфатных групп кольцо расправляется. Компактность молекулы уменьшается, ее наружные размеры увеличиваются (форма II). Что касается линейной двухните-вой молекулы ДНК (форма III), то в зависимости от среднего размера пор геля она может мигрировать быстрее или медленнее, чем сверхскрученное кольцо одинаковой с ней массы. Для крупнопористого геля решающим фактором может оказаться компактность формы I; для более мелких пор на первый план выступает большая гибкость линейной молекулы ДНК (форма III).
На рис. 1, на правом треке показан результат фракционирования ДНК E.coli: основной ее массы (2), кольцевой плазмидной ДНК (1) и сверхскрученной плазмиды (3) в 0,8% геле агарозы. (Telford, Science 195, 391, 1977). На левом треке того же рисунка указаны положения «маркеров» — плазмид известной молекулярной массы (соответствующие цифры указаны в миллионах дальтон).

Рис.1
Скорость миграции линейных двухнитевых молекул ДНК уменьшается с увеличением их массы, но лишь до определенного предела. При молекулярной массе более 5 миллионов в 1,6%-ном геле агарозы и при массе более 12 миллионов дальтон в 0,8% -ной агарозе молекулы мигрируют практически с одинаковой скоростью, независимо от их массы. В этих случаях решающую роль играет гибкость — способность очень длинных молекул, извиваясь, проходить через гель одинаково легко (или одинаково трудно) при любой длине.
Рибосомальные РНК могут иметь существенно более невыгодную для миграции в геле вторичную структуру, чем ДНК. Дело в том, что у крупных молекул РНК эта структура представлена многочисленными, торчащими во все стороны «шпильками». Это — места локального спаривания в жесткие двунитевые структуры отдельных, зачастую далеко отстоящих друг от друга по основной последовательности, комплементарных участков РНК. Такая молекула уже не может продвигаться через поры геля Собирая основной «сэндвич» (гель, фильтр и облегающие их листки фильтровальной бумаги) следует проверять, что между ними не остаются пузырьки воздуха.
Перенос ДНК на нитроцеллюлозный фильтр занимает 2—3 часа. Следует заметить, что короткие фрагменты ДНК на нитроцел-люлозном фильтре задерживаются плохо. Если это существенно, то лучше использовать фильтры из так называемой, «диазобума-ги» или «ДБМ-бумаги». Фирма Ватман выпускает ее под наименованием «Whatman 540».
Литература
1 Курашвили Л.В. Нарушения, липидного обмена при состояниях напряжения. II Захарьинские чтения. Тезисы докладов. - 1995. -С.127.
2 Курашвили Л.В. Активность липазы и ЛХАТ при длительном обезвоживании. В кн.: Актуальные вопросы диагностики, лечения и реабилитации больных. Тезисы докладов. - Пенза, 1995. -С.71-72.
3 Курашвили Л.В. Фосфолипидный статус при нарушении водно-электролитного обмена. В кн.: Актуальные вопросы диагностики, лечения и реабилитации больных. Тезисы докладов. - Пенза, 1995.-С.69-70.
Похожие работы
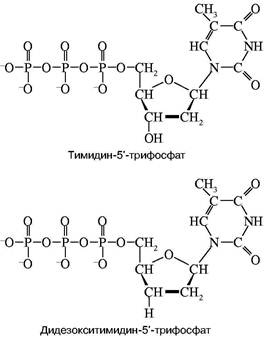
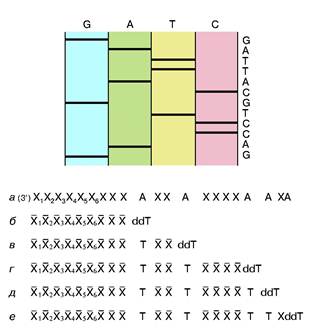
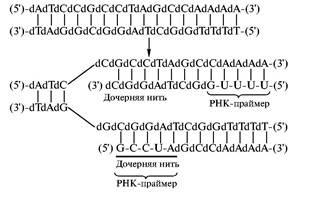
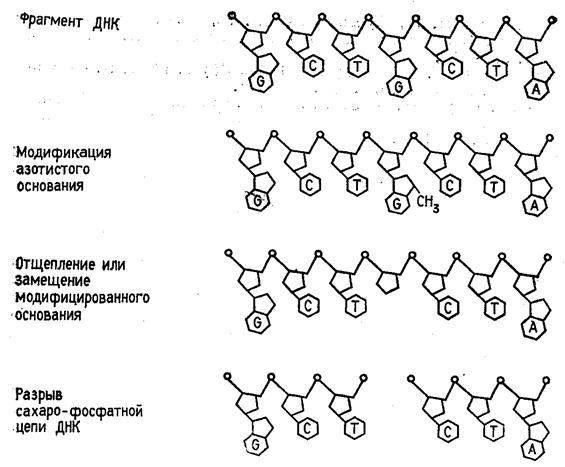
... то есть ген, способствующий неограниченному размножению клеток — клетки становятся злокачественными и возникает раковая опухоль. Что современная химия и биохимия умеют делать с нуклеиновыми кислотами Химия и биохимия нуклеиновых кислот не только углубили наши представления об огромной группе важнейших биологических процессов, связанных с сохранением, размножением и использованием наследственной ...
... (например, семейства генов) необходимо провести клонирование ПЦР-продуктов в векторах типа М13, в результате каждый фаг будет содержать только одну вставку. При прямом секвенировании смеси генов наблюдается несколько одинаково расположенных полос в разных дорожках геля. При амплификации же одного гена можно проводить прямое секвенирование, не прибегая к промежуточному субклонированию. Выбор ...
0 комментариев