Размножение
Различают вегетативное, бесполое и половое размножение.
Вегетативное размножение
К вегетативному можно отнести те процессы размножения, при которых части таллома отделяются без каких-либо заметных изменений в протопластах.
Вегетативное деление клеток десмидиальных, диатомовых, многих одноклеточных сине-зеленых водорослей, продольные деления монадных представителей, распад – диссоциация колоний на две или большее число частей (например, у синуры, диктиосфериума) – примеры вегетативного размножения. Распространенным примером вегетативного размножения нитчатых и более высокоорганизованных форм является фрагментация–разрыв талломов на отдельные участки. Этот способ наиболее обычен среди зеленых водорослей и достигает кульминации у таких родов, как клебсормидиум (нити нередко распадаются на отдельные клетки) и особенно у рода стихококкус, который встречается в виде отдельных клеток (нити даже из нескольких клеток редки). Диссоциация нитей на короткие отрезки или даже отдельные клетки легко происходит у зигнемальных.
Вегетативное размножение широко распространено и среди бурых водорослей. Гаметофиты рода ламинария, происходящие от одной зооспоры, могут распадаться на неограниченно большое число особей. Среди фукальных крупные скопления поддерживаются исключительно путем процесса регенерации участков таллома, то же относится к пелагическим видам рода саргассум.
Лежащая в основе вегетативного размножения способность регенерации целого таллома из отдельных частей или даже клеток экспериментально продемонстрирована и для красных водорослей (гриффитзия, родохортон). Этим объясняется их встречаемость в природе там, где внешние условия не благоприятствуют прорастанию спор.
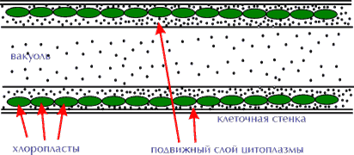
Более специализированные структуры, служащие для вегетативного размножения, – выводковые почки или пропагулы. Они могут быть одноклеточными, многоклеточными, сифонового строения, обычно содержат обильные запасные вещества и вырастают непосредственно в новый таллом. В качестве примера можно привести многоклеточные выводковые почки бурой водоросли сфацелярия с довольно сложной морфологией, различающейся у разных видов: У одних они двухраздельные, у других – трехраздельные. Пропагулы сифонового строения, по форме поразительно похожие на трехраздельные пропагулы сфацелярии, описаны для зеленой бриопсидальной водоросли дербезии. У другой бриопсидальной водоросли – кодиума – пропагулы в виде легко отделяющихся специализированных веточек. С многоклеточными пропагулами можно сблизить клубеньки харофициевых водорослей, которые образуются у некоторых видов на нижних, погруженных частях таллома и на ризоидах. У ряда сифоновых и сифонокладальных водорослей (бриопсис, галицистис, валония) голые участки цитоплазмы – протопласты, вытекающие из таллома при его повреждении, очень быстро (в течение нескольких минут) секретируют новые клеточные стенки и развиваются в соответствующие талломы.
Специализированным типом вегетативного размножения можно считать и образование структур, предназначенных для перенесения периодов, неблагоприятных для обычного вегетативного роста. В этих случаях в клетках утолщаются оболочки, в протопласте накапливаются запасные вещества – жир, крахмал. Такие клетки, называемые акинетами, встречаются у многих нитчатых форм (улотрикс, микроспора, кладофора).
Бесполое размножение
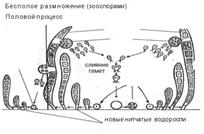
Бесполое размножение осуществляется посредством особых спор. Самый обычный способ бесполого размножения у многих водорослей – посредством зооспор, как правило, голых монадных клеток. Из содержимого каждой клетки может сформироваться только одна зооспора (например, у эдогониума) или чаще содержимое клетки делится на две, четыре, восемь и более частей и продуцируется соответствующее количество зооспор. У многих водорослей клетки, в которых формируются зооспоры, специально не дифференцированы, однако у трентеполиевых зеленых водорослей и у всех бурых есть спорангии, образующиеся только во время бесполого размножения и отличающиеся по форме и размерам от обычных вегетативных клеток.
В ряде случаев выяснены (экспериментально) условия внешней среды, стимулирующие переход к зооспорообразованию. Для многих нитчатых зеленых водорослей таким фактором служит повышенное содержание в воде свободной углекислоты. Это объясняет старые наблюдения над массовым зооспорообразованием при перенесении нитей из текучей воды в стоячую, со света в темноту, т.е. в условия, когда в воде накапливается углекислота.
В связи с зооспорообразованием уместно остановиться на возникновении так называемых синзооспор. Синзооспоры – образования, соответствующие комплексу двух или многих сросшихся в различной степени зооспор, – наблюдались у разных водорослей, но особенно часто у разножгутиковых. Нередко при выходе из материнской клетки такие зооспоры остаются соединенными по две или по нескольку лишь посредством тонких цитоплазматических нитей и позднее разъединяются. Однако часто зооспоры оказываются связанными своими задними концами гораздо прочнее и на большем протяжении, так что свободными остаются только их передние концы со жгутиками или же, наконец, они образуют сплошную цитоплазматическую массу, из которой выступают пары жгутиков, указывая на число соединившихся зооспор. Такие стадии очень похожи на синзооспоры вошерии, которым они полностью соответствуют.
Синзооспоры часто встречаются у бочридиопсиса. трибонемы, ботридиума и др., однако, как правило, эти водоросли образуют двужгутиковые зооспоры. Эти случайные отклонения стабилизировались лишь у вошерии. Ее крупные многожгутиковые зооспоры уже давно рассматривали как сложную зооспору, образующуюся внутри спорангия, в котором деление спорапгиального протопласта на одноядерные двужгутиковые споры не наблюдается.
Синзооспоры имеются также у зеленых (сфероплея, кладофора, хламидомонас и др.) и бурых водорослей (Ectocarpus siliculosus).
У значительного числа водорослей вместо, зооспор образуются цпланрслоры – неподвижные, лишенные жгутиков споры. Примерами их могут служить моно- и тетраспоры ктэасных водорослей, тетраспоры диктиотовых из бурых водорослей. В тех случаях, когда апланоспоры, будучи заключенными в оболочку материнской клетки, принимают все отличительные черты этой клетки (характерные очертания, особенности оболочки), их называют автоспорами (например, у многих хлорококкальных из зеленых водорослей).
У ценобиальных (монадных и коккоидных) зеленых водорослей при бесполом размножении образуются дочерние колонии. В ряде групп водорослей бесполое размножение отсутствует: у конъюгатофициевых, харофициевых, многих зеленых водорослей сифонового строения, у всех диатомовых, фукальных из бурых водорослей.
На примере некоторых водорослей удалось глубже проникнуть в механизмы освобождения клеток бесполого размножения и их последующего прикрепления к субстрату. Вот некоторые примеры. Для многих (более 40) видов хламидомонады было показано, что молодые клетки выделяют литический фактор – автолизин спорангиальной стенки, вызывающий лизис клеточной стенки спорангия, что способствует освобождению зооспор. Фермент действует только на спорангиальную стенку, но не на стенки вегетативных клеток и зооспор и обладает групповой специфичностью, т.е.: влияет только на морфологически сходные виды, а в некоторых случаях – только на спорангии вида-продуцента.
У вольвокса клетки также образуют и выделяют в окружающую среду ферменты, лизирующие инволюкрум материнского ценобия. облегчая этим освобождение дочерних сфероидов. Из культуральной среды Chiorella fusca var. vacuolates был выделен фермент (отсутствующий в среде до начала сноруляции), разрушающий клеточную стенку материнских клеток автоспор и не действующий на клеточные стенки самих автоспор, У вошерии в. кончике синзооспоры были обнаружены многочисленные везикулы, по-видимому, дериваты диктиосом, содержащие предположительно целлюлазу и другие энзимы, разрушающие полисахариды. Высказано предположение, что они ответственны за разрушение стенки в апикальной области спорангия – необходимое условие выхода синзооспоры. В зооспорах зеленой водоросли Oedogonium cardiacum, бурой – Chorda tomentosa, тетраспорах красной водоросли Palmaria palmata обнаружены обильные везикулы, содержащие клеющий материал, который, возможно, способствует прикреплению спор к субстрату. То же, по-видимому, справедливо и для тетраспор и карпоспор Ceramium rubrum.
Половое размножение
Половое размножение (воспроизведение), сущность которого заключается в слиянии – копуляции двух различных в половом отношении клеток-гамет с образованием зиготы, известно у большинства эукариотических водорослей. Во многих случаях, особенно у водорослей с монадной организацией, обнаружению полового процесса способствовала разработка методов их культивирования в лабораторных условиях. Например, при определенных условиях культивирования в последние годы был описан половой процесс у ряда динофлагеллят, ранее считавшихся лишенными его.
Половое размножение – сложный многоступенчатый процесс, начинающийся с половой дифференцировки клеток–гаметогенеза, включающий слияние гамет (происходящее в несколько этапов и приводящее к образованию зиготы) и завершающийся мейозом.
Условия, вызывающие половой процесс. Переход к половому размножению и дифференцировке гамет зависит не только от внутренних причин, нередко определяемых возрастом водоросли, но и от факторов внешней среды. Среди этих факторов для многих зеленых водорослей ведущую роль играет содержание в среде азота. Исчерпание в среде азота и наступающее азотистое голодание служит сигналом для начала полового размножения многих видов Chlamydomonas (Ch. reinhardtii, Ch. moewusii var. rotunda, Ch. chlamydogama, Ch. eugametos), Pandorina morum, P. unicocca, Chlorococcum echinozygotum, Golenkinia minutissima, Scenedesmus obliquus, Oedogonium cardiacum и др. Хорошим примером зависимости половой дифференцировки от содержания в среде азота служит зеленая водоросль Sphaeroplea annulina. Эта азотолюбивая водоросль встречается у нас преимущественно ранней весной в мелких водоемах и уже в мае при понижении содержания в воде азота переходит к половому воспроизведению. В водоемах с более постоянным подтоком азотистых веществ, например на полях орошения Москвы, сфероплея вегетирует иногда почти все лето. В лаборатории также можно поддержать ее в вегетативном состоянии, культивируя в среде со значительным содержанием азота; перенесением же в среду, бедную солями азота, легко можно вызвать половой процесс.
У перечисленных и многих других видов источники азота (нитраты, аммоний, мочевина, глютамин и пр.), поддерживающие рост, препятствуют половой дифференцировке, которая наступает в условиях, неблагоприятных для вегетативного роста. Рост и размножение находятся как бы в антагонистических отношениях друг с другом. В отличие от большинства зеленых водорослей у вольвоксов половое размножение с последующим образованием покоящихся зигот происходит в то время, когда условия среды оптимальны для вегетативного роста. Это объясняется тем, что половой процесс у этого рода сопряжен с продукцией специальных половых ценобиев, в которых и формируются гаметы. В менее оптимальных для вегетативного роста условиях половые сфероиды уменьшались бы в размерах, понижалась бы их витальность. Половая дифференцировка у вольвоксов наступает под действием видоспецифичных половых гормонов – индукторов, выделяемых мужскими ценобиями, появляющимися в популяции спонтанно.
Дефицит в среде азота вызывает половой процесс и у многих динофлагеллят.
Для индуцирования полового процесса у десмидиальных водорослей наиболее важными факторами оказываются свет и температура, хотя в некоторых случаях формирование конъюгационных выростов и образование зигот наблюдалось при недостатке в среде азота. Свет и температура являются также решающими факторами среды, оказывающими воздействие на гаметогенез многих бурых водорослей. Так, для многих ламинариальных водорослей было показано, что переход их гаметофитов к гаметогенезу контролируется синим светом достаточно высокой интенсивности при низких (5–10°С) температурах. У организмов, приуроченных к водам с высоким содержанием солей, например у Dunaliella salina, понижение концентрации солей в среде вызывает половой процесс.
Формы полового процесса. Форма полового процесса может быть различной. У ряда лишенных клеточной оболочки монадных водорослей сливаются, (копулируют) клетки, не отличимые от вегетативных. Такая форма полового процесса называется хологамией (например, у зеленой водоросли дуналиелла). У многих водорослей гаметы, как и зооспоры, образуются внутри материнских клеток и, выйдя из них, копулируют попарно. Если обе сливающиеся монадные гаметы не различаются морфологически, половой процесс называют изогамным – изогамией, при копуляции монадных гамет, различающихся по размерам, имеет место гетерогамия или анизогамия. Более крупные макрогаметы трактуются как женские, более мелкие – микрогаметы – как мужские. При моогамном половом процессе – крупные женские гаметы лишены шулшов, неподвижны и называются яйцеклетками или яйцами, более мелкие мужские гаметы со жгутиками подвижны и носят название сперматозоидов или антерозоидов.
У красных водорослей яйцеклетки оплодотворяются мужскими клетками, лишенными жгутиков, – спермациями.
У зеленых водорослей за единичными исключениями наблюдается истинная оогамия, для которой характерно оплодотворение яйца внутри оогония – на месте образования (in situ) в отличие от примитивной оогамии, когда яйцеклетка покидает оогонии до оплодотворения, осуществляющегося в окружающей среде. Примитивная оогамия встречающаяся среди зеленых водорослей лишь в виде исключений, является правилом для оогампых бурых водорослей. У центрофициевых оогамных диатомовых наблюдается как истинная, так и примитивная оогамия. Если сливаются протопласты неподвижных клеток, не отличающихся от вегетативных, говорят о конъюгации.
Обычно предполагается, что форма полового процесса эволюционировала в следующем направлении: изогамия-» анизогамия-» оогамия. Эта прогрессивная эволюция началась с возникновения различающихся по размерам гамет у изогамных предковых форм. Образование мелких гамет повышает их нумерическую продуктивность – их число, но за счет редукции количества цитоплазмы. С другой стороны, большая масса цитоплазмы и резервных материалов у более крупных гамет (возникающих в материнских клетках в меньшем числе) необходимы для начального развития будущей зиготы. Таким образом, раз возникший диморфизм гамет дает основу для избирательного слияния между гаметами, различающимися по размерам: гаметы минимальных размеров получают компенсацию в количестве цитоплазмы посредством слияния с более крупными гаметами. Под эти чисто теоретические рассуждения в последнее время была подведена экспериментальная основа.
Л. Визе удалось экспериментально получить фенотипическую анизогамию у трех изогамных гетероталличных видов Chlamydomonas. У этих видов каждый тип спаривания («+» и «–») имеет два пути гаметогенеза, которые индуцируются подбором условий культивирования. Первый путь гаметогенеза заключается в том, что протопласт вегетативной клетки в результате внутриклеточной перестройки непосредственно превращается в одну крупную гамету. При втором способе гаметогенеза в материнской клетке происходят два или три митоза, и образуется 4–8 гамет меньшего размера. Комбинированием совместимых гамет, сформировавшихся разными способами, может быть получена фенотипически анизогамная копуляция. Высказано предположение, что любая мутация, которая зафиксировала бы тот или иной способ гаметогенеза (блокировала бы один путь гаметогенеза в пользу другого), дала бы непосредственно штаммы, производящие исключительно или микро-, или макрогаметы. Таким образом, могли возникнуть уже генетически анизогамные линии. В появившемся в результате мутации анизогамном таксоне при дальнейшей эволюции может быть утеряна подвижность у более крупных гамет. Сначала жгутики еще сохраняются, как у Ch. suboogama, полная же их потеря привела бы к настоящей оогамии как у Ch. coccifera. Ch. pseudogigantea.
Эволюция формы полового процесса происходила независимо от эволюции таллома. Это впервые продемонстрировал русский альголог И.Н. Горожанкин в конце прошлого века, обнаружив у разных видов хламидомонады (стоящих на одном уровне морфологической дифференцировки таллома) все основные типы полового процесса: наряду с изогамией, преобладающей у одноклеточных вольвокальных, гетерогамию у Ch. braunii и оогамию у Ch. coccifera.
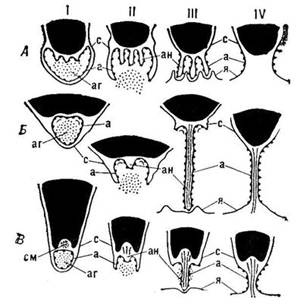
Гаметогенез. У форм с изогамным и гетерогамным половым процессом гаметогенез в большинстве случаев не связан с формированием каких-либо обособленных морфологических структур: изо- и гетерогаметы возникают в обычных вегетативных клетках, как и зооспоры, но обычно в большем числе и соответственно меньших размеров. Исключение составляют изогамные и гетерогамные бурые водоросли, где гаметы образуются в специальных органах: многогнездных или многокамерных спорангиях, в каждой камере (клетке) которых формируется по одной гамете. Оогамия у монадных и коккоидных форм обычно также не сопряжена с изменением клеток, производящих гаметы: они не отличаются от вегетативных. Наоборот, у водорослей с нитчатой и тканевой организацией таллома яйцеклетки и сперматозоиды развиваются в гаметангиях, как правило, резко отличающихся по форме от вегетативных клеток и носящих специальные названия оогонии и антеридии (сперматогонии).
Растения, производящие гаметы, могут быть обоеполыми (однодомными) и раздельнополыми (двудомными). В первом случае гаметы разного пола возникают на одном и том же талломе, во втором – на разных. Кроме понятий «однодомность» и «двудомность» существуют понятия «гомоталлизм» и «гетероталлизм». При гомоталлизме половой процесс имеет место в пределах клональной популяции (клон – культура, полученная из одной клетки). При гетероталлизме половой процесс возможен только при смешении клеток из разных клонов противоположного типа спаривания или знака; в случае гетероталличных изогамных форм гаметы разного типа спаривания обозначаются знаками «+» и «–». Понятия «гомоталличный» и «гетероталличный» не всегда совпадают с понятиями «однодомный» и «двудомный». Так, среди вольвоксов имеются гомоталличные и гетероталличные виды, включающие как однодомные, так и двудомные штаммы. Например, внутри клональной популяции могут развиваться особи как однодомные с яйцами и сперматозоидами в одном сфероиде, так и двудомные, у которых половые органы находятся в разных индивидах.
Освобождение репродуктивных клеток. Некоторые морские водоросли, обитающие в верхней части литоральной зоны, обнаруживают периодичность в формировании и освобождении репродуктивных клеток как бесполого (спор), так и полового (гамет) размножения. Так, у берегов Англии у водоросли Dictyota dichotoma формирование половых органов (антеридиев и оогониев) приурочено к летним месяцам и происходит периодически с двухнедельными интервалами, что соответствует таковым между двумя сигизийными приливами, а освобождение гамет – яиц и сперматозоидов – наступает через несколько приливов после наивысшего из последующих квадратурных приливов и продолжается 7–9 дней. Таким образом, периодичность находится в строгом соответствии с лунным ритмом. Периодичность сохраняется и при культивировании талломов в лаборатории. У одного и того же вида ритм освобождения гамет зависит от географического положения местообитания. Так, у D. dichotoma в Северной Каролине периодичность наблюдалась, но иная, чем у берегов Англии: гаметы освобождались не через двухнедельные интервалы, а через месячные, а у берегов Ямайки половые органы формировались периодически, но период созревания гамет растягивался настолько, что выход их происходил почти непрерывно.
Наряду с полулунным ритмом в освобождении яйцеклеток у D. dichotoma была описана и дневная периодичность. При культивировании в условиях смены света (14 ч) и темноты (10 ч) вскоре после начала светового периода наблюдался активный выход яйцеклеток, который продолжался в течение часа, после чего освобождение яиц прекращалось до начала следующего светового периода, когда вновь начиналось освобождение яиц. Дневной выход яиц может быть индуцирован очень слабым кратковременным освещением. Так, 50% яиц зрелого оогония освобождается после 20-секундного освещения белым светом в 1 Лк. Выход яиц зависел также от температуры: если при 20С максимум освобождения яиц достигается через 25–30 мин после начала дневного светового периода, то при 10С этот максимум наступал только через 100 мин. Эффект света данной интенсивности также зависит от температуры. Кроме того, вскрытие полностью созревшего оогония может быть ускорено или замедлено путем изменения осмотического давления среды.
Специальное исследование по выяснению влияния некоторых внешних условий на периодичность освобождения гаме было проведено в Японии над популяциями обычной в прибрежных водах водоросли – Monostroma nitidum, занимающей второе (после видов порфиры) место в качестве объекта промышленного культивирования. Это культивирование осуществляется обычно двумя методами: на хибисетках, расположенных горизонтально на такой глубине, при которой талломы обнажаются во время отлива приблизительно на 4 ч, и на бета-сетках с растениями, постоянно погруженными в морскую воду вблизи ее поверхности. Образование гамет легко распознается невооруженным глазом благодаря желтоватой окраске плодущей части таллома. Были проведены сравнительные наблюдения над талломами, произрастающими поблизости друг от друга, но в различающихся условиях: на прибрежных скалах – в естественных местах обитания, на хиби-сетках и бета-сетках. Период плодоношения (с февраля по июнь) во всех грех случаях был сходным, а периодичность в освобождении гамет оказалась разной. Более четко (с двухнедельными интервалами, во время каждого квадратурного прилива) периодичность была выражена у растений, растущих на скалах. Менее четко она обнаруживалась у талломов на хиби-сетках, а у постоянно погруженных талломов на бета-сетках никакой периодичности не наблюдалось. Продемонстрировано также, что периодичность в образовании гамет зависит от освещения (интенсивность света и продолжительность фотопериода) и температуры.
Периодичность наблюдалась и в спороношении Viva pertusa, в освобождении яйцеклеток у Sargassum muticum.
В последние годы в ряде работ прослежена дифференцировка и механизм освобождения спермациев у красных водорослей на электронно-микроскопическом уровне. Так, у Ptilota plumosa спер-матогонии, в которых образуются мужские гаметы – спермании, обладают ясно выраженной полярностью, заключающейся в том, что клеточное ядро занимает апикальное положение, а в базальной части молодого сперматогоиия находится одна или несколько вакуолей. В зрелых сперматогониях вакуоли замещаются крупными фиброзными исчерченными вакуолями, занимающими приблизительно половину объема клетки и возникающими, возможно, за счет слияния пузырьков, происходящих от диктиосом. Фиброзные вакуоли содержат мукополисахарид, Вокруг клеточного ядра и окружающей цитоплазмы, по-видимому, образуется новая плазмалемма, не включающая базальных фиброзных исчерченных вакуолей. Таким образом формируется спермаций, который покидает оболочку сперматогония. Одновременно с разрывом клеточной стенки сперматогония опорожняются базальные крупные фиброзные вакуоли, что способствует освобождению спермация. Спермаций без клеточной оболочки одет тонкой слизистой обверткой, возможно происходящей от матрикса фиброзной вакуоли. Эта обвертка играет важную роль во время прилипания спермация к трихогине. Сходные процессы наблюдаются и при дифференцировке и освобождении спермация у Polysiphonia hendryi. У ламинарии при образовании сперматозоидов аппарат Гольджи выделяет большое количество слизи, способствующей разрыву антеридия и освобождению сперматозоидов.
У вошерии везикулы, возможно, содержащие целлюлазы, наблюдались в апикальной части оогониального клювика, у места, где сперматозоиды входят в оогоний вскоре после того, как клювик образует пору. Такие же везикулы наблюдались вблизи кончика антеридия, где выходят сперматозоиды.
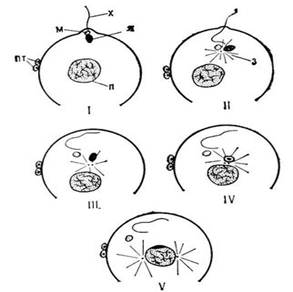
Копуляция. Слияние – спаривание гамет, приводящее к возникновению зигот, происходит в несколько этапов. Первый из этих этапов – образование групп, в которых гаметы разного пола вступают в первоначальный контакт. Если в капле воды или культуральной жидкости наблюдать поведение снабженных жгутиками гамет одного пола (в случае изогамет – одного типа спаривания или одного полового знака «+» или «–»), можно убедиться, что гаметы равномерно рассеяны по всему полю зрения микроскопа. Стоит к капле с такими гаметами добавить гаметы противоположного пола (знака), как сейчас же образуются скопления – группы. Это впервые наблюдалось в конце прошлого века у изогамной бурой водоросли Ectocarpux siliculosus, где много гамет одного пола окружают одну гамету противоположного пола. Это было продет моистрировано с применением витального окрашивания гамет противоположных полов в разные цвета. Сходным образом образуются группы гамет у Chlamydomonas раирега.
У других водорослей каждая группа содержит по многу гамет обоих полов, что показано у Dunalielld saltna путем смешения окрашенных в зеленый цвет гамет одного пола (образовавшихся в культуре с избытком азота и фосфора) с окрашенными в оранжевый цвет гаметами противоположного пола (такая окраска возникает в культурах с недостатком в среде азота и фосфора). Среди анизогамных водорослей, например у Ulva lobata, микроскопические наблюдения показывают, что в одной группе имеется много гамет обоих размеров.
Образование групп уже давно связывалось с выделением каких-то половых веществ (аттрактантов), хемотаксически привлекающих гаметы противоположного пола. На такой вывод наталкивали и самые ранние наблюдения за половым процессом у водорослей (фукуса, эдогониума).
В ряде случаев присутствие привлекающих половых веществ было продемонстрировано экспериментально, например, у зеленой водоросли Sphueroplea annulina. У нее наблюдается оогамный половой процесс: в одних клетках (оогониальных) образуется много яйцеклеток, в других (антеридиальных) – колоссальное количество двужгутиковых сперматозоидов золотистого цвета. При созревании половых клеток в стенках оогониев и антеридиев возникают отверстия. Вышедшие сперматозоиды скапливаются у отверстий, ведущих в оогоний, врываются в оогоний и своими движениями приводят в движение яйцеклетки. Если среди нитей сфероплеи со зрелыми оогониями поместить хлопчатобумажные ниточки, они абсорбируют предполагаемый агтрактант и, перенесенные в равномерную суспензию сперматозоидов, привлекают их: сперматозоиды скапливаются вокруг таких ниточек. Выделение аттрактантов гаметами Tetraspora lubrica было продемонстрировано иным способом: при добавлении в жидкость из-под гамет одного знака, удаленных центрифугированием, гамет противоположного знака, последние образовывали группы. Такая изоаглютинация наблюдается и у других водорослей.
Делались также попытки установить химическую природу аттрактантов (иначе гамонов). Однако все более ранние попытки оказались тщетными, что в значительной мере объясняется ограниченными в то время возможностями аналитической химии. Лить в 70-е годы XX в. для нескольких морских бурых водорослей удалось установить точную химическую природу аттрактантов. Это стало возможным только благодаря развитию методов аналитической химии, а также в связи с возможностью культивирования водорослей в условиях лаборатории, позволяющей накопить большую биомассу, необходимую для биохимических исследований.
У некоторых водорослей аттрактанты, по-видимому, отсутствуют. Например, у Volvox rousseletii глобоиды сперматозоидов, освободившиеся из мужских ценобиев. плавают в воде быстро, и в присутствии женских ценобиев могут прикрепиться к ним. Однако специфическое привлечение отсутствует, скорее, имеет место случайное столкновение. Нередко наблюдаются глобоиды сперматозоидов, прикрепленные к вегетативным и мужским сфероидам.
В группах осуществляется первичный контакт гамет и их попарное слияние, начинающееся со слияния их цитоплазм – плазмогамии.
Использование альгологически чистых и аксеничных культур водорослей и развитие методов, позволяющих индуцировать гаметогенез, сделали возможным более тщательное и детальное исследование процессов оплодотворения у водорослей как в световом, так и в электронном микроскопе, а иногда даже с применением микрокипематографии (например, у Prasiola scipitata, AscophyUum nodosum, Halidrys siliquosa, Oedogonium cardiacum). В ряде случаев процесс действительного слияния половых клеток удалось наблюдать при использовании штаммов, специально полученных в условиях культуры и не встречающихся в природе. Так, слияние сперматозоида с яйцеклеткой у вольвокса (Volvox carteri f. nagariensis) удалось наблюдать у женского мутанта, характеризующегося большим числом яиц и небольшим (по сравнению с диким штаммом) количеством соматических клеток – обилие их затрудняло наблюдение этого процесса. У полученного в культуре женского диплоидного штамма Oedogonium cardiacum более крупные, чем у гаплоидного (обычного в природе), оогонии, в которых, как правило, обнаруживается большее пространство между поверхностью яйца и окружающей оогониальной стенкой, что дает явное преимущество для наблюдения начальных стадий слияния гамет в живом материале. Конкретные примеры плазмогамии и следующего этапа полового слияния – кариогамии будут даны при описании родов, у которых этот процесс подробно исследовался.
В результате полового процесса образуется зигота, содержащая одно купуляционное диплоидное ядро – продукт слияния двух гаплоидных ядер, происходящих из той и другой гаметы. Обычно зигота окружается толстой оболочкой, переполняется запасными продуктами и растворимым в жирах пигментом кирпично-красного цвета – астаксантином. Зигота прорастает или непосредственно после образования, или после более или менее продолжительного периода покоя.
Жизненный цикл. Соотношение диплоидной и гаплоидной фаз в жизненном цикле разных водорослей неодинаково. В одних случаях прорастание зиготы сопровождается редукционным делением копуляционного ядра (зиготическая редукция) и развивающиеся при этом растения оказываются гаплоидными. Так, у многих зеленых водорослей (вольвокальные, большинство хлорококкальных, конъюгатофициевые, харофициевые) зигота – единственная диплоидная стадия в цикле развития, вся вегетативная жизнь проходит у них в гаплоидном состоянии, они являются гаплонтами.
У других водорослей, наоборот, вся вегетативная жизнь осуществляется в диплоидном состоянии, а гаплоидная фаза представлена лишь гаметами, перед образованием которых и происходит редукционное деление ядра (гаметическаяредукция). Зигота без редукционного деления ядра прорастает в диплоидный таллом. Эти водоросли – джлонты. Таковы многие зеленые водоросли, имеющие сифоновое строение, все диатомовые, из бурых – представители порядка фукальные.
У третьих редукционное деление ядра предшествует образованию зооспор или апланоспор, развивающихся, как правило, на диплоидных талломах (спорическая редукция). Эти клетки бесполого размножения вырастают в гаплоидные растения, размножающиеся только половым путем. После слияния гамет зигота развивается в диплоидное растение, несущее только органы бесполого размножения. Таким образом, у этих водорослей имеет место чередование форм развития (генераций): диплоидного бесполого спорофита и гаплоидного полового гаметофита. Оба поколения могут быть одинаковы морфологически (изоморфная смена генераций) или же резко различны по внешнему виду (гетероморфная смена генераций). Изоморфная смена генераций характерна для морских видов ульвы, энтероморфы, кладофоры, хетоморфы из зеленых водорослей, для ряда порядков бурых и большинства красных водорослей (несколько усложненная), Гетероморфная смена генераций особенно распространена среди бурых водорослей, но встречается у зеленых и красных. Следует отметить, что у ряда этих водорослей, имеющих смену генераций, помимо двух кардинальных моментов в их жизненном цикле – копуляции (сиигамии), сопровождающейся удвоением числа хромосом в копуляционном ядре, и мейоза, приводящего к редукции числа хромосом, – могут происходить и иные процессы: спонтанное умножение числа хромосом, партеногенетическое развитие гамет, явление гетеробластии. В результате способ размножения (бесполое или половое) далеко не всегда коррелирует с плоидностью таллома, т.е. спорофиты могут быть не только диплоидными, но и гаплоидными и тетраплоидными, а гаметофиты обнаруживают не только гаплоидный, но и диплоидный наборы хромосом и т.п. Такие сложные циклы развития были установлены, например, у эктокарпуса, ульвы. Кроме того, при определенных внешних условиях из жизненного цикла одной и той же водоросли могут выпадать какие-либо стадии, например, у эктокарпуса происходит репликация спорофита посредством нейтральных зооспор, и в некоторых местообитаниях известны только диплоидные талломы этой водоросли.
У одного и того же вида наблюдаются различные жизненные циклы. Так, у зеленой водоросли Bryopsis hypnoides встречается как однофазный цикл, так и двухфазное развитие, у красных водорослей Gigartina steliata, G. agardhtii отмечается как половой цикл, так и апомиктическое (без слияния половых клеток) развитие. Даже у таких высокоорганизованных форм, как ламинария, где, как правило, регулярная смена спорофита и гаметофита носит облигатный характер, возможны отклонения в жизненном цикле: партеногенетическое (из неоплодотворенной яйцеклетки) развитие спорофитов и апоспорическое (из вегетативных клеток спорофита) развитие гаметофитов.
Жизненные циклы водорослей очень сложны и разнообразны. Особенно это относится к красным водорослям, исследование развития которых позволило обнаружить гораздо больше типов жизненных циклов, чем это было известно ранее. Культуральные исследования позволили установить связи между водорослями, долгое время считавшимися самостоятельными видами и оказавшимися лишь стадиями в жизненном цикле одной и той же водоросли, т.е. значительно умножились известные нам случаи гегероморфной смены генераций. Это относится не только к красным водорослям, но и к зеленым – улотрихальным, бриопсидальным.
Наконец, у немногих водорослей, например у водоросли празиолы (из зеленых) – леманеи, батрахоспермума (красные), мейоз происходит в некоторых вегетативных клетках диплоидного таллома (соматическая редукция), из которых затем развиваются гаплоидные талломы. Подробнее жизненные циклы будут разбираться при описании соответствующих групп.
Похожие работы
... , что свидетельствует об известном параллелизме эволюционного развития в пределах этих отделов. В настоящее время различают 9 основных типов морфологической структуры тела водорослей. Из них 4 относятся к одноклеточным формам, 1 – к неклеточным, остальные 4 – к многоклеточным. 1. Амёбоидная структура представлена одноклеточными организмами; 2. Монадная структура свойственна одноклеточным ...
ерхового брожения типа Hefeweizen, Wei bier и др. Благодаря высокой экстрактивности доля пшеничного солода при производстве пшеничного пива находится в пределах 50-60%. [4] В качестве пивоваренных используются только некоторые сорта пшеницы, причем большим спросом пользуются озимые сорта из-за более низкого содержания белка и повышенного содержания экстракта, а также как дающие более светлое ...
... ), коренных зубов всего четыре (по два на верхней и нижней челюстях), имеют хобот, который образуется в результате сращения носа и верхней губы. Источники 1) Голубева Е., ЗАНИМАТЕЛЬНОЕ ЕСТЕСТВОЗНАНИЕ, Нескучный учебник, Тригон, С-П., 2002 г.,365с. 2) Рохлов В., Теремов А., Петросова Р., ЗАНИМАТЕЛЬНАЯ БОТАНИКА, "АСТ-ПРЕСС", М., 2003 г., 430 с. 3) ПРИРОДА РОССИИ: ...

... — слияние мужской и женской половых клеток — гамет, в результате чего образуется зигота, способная развиваться в новый организм. Оплодотворение лежит в основе полового размножения и обеспечивает передачу наследственных признаков от родителей потомкам. Оплодотворение у растений. Оплодотворение свойственно большинству растений; ему обычно предшествует образование гаметангиев — половых органов, в ...
0 комментариев