Основные группы ферментов генетической инженерии
Механизм действия рестриктаз
Обратная транскриптаза
Репортерные гены, кодирующие нейтральные для клеток белки, наличие которых в тканях может быть легко тестировано
Регуляция экспрессии гена у прокариот
Наличие интронов и экзонов в молекуле ДНК
Способы прямого введения гена в клетку
Введение генов в клетки млекопитающих
Генотерапия
Получение трансгенных животных
Навигация
Механизм действия рестриктаз
рДНК-биотехнология. Способы биотрансформации клеток
81258
знаков
0
таблиц
5
изображений
1.1.1 Механизм действия рестриктаз
В качестве мишеней (мест узнавания) часто выступают палиндромы из 4-6 пар оснований - сайты рестрикции. Точки узнавания рестриктазами симметричны относительно поворота на 180оС, то есть последовательность нуклеотидов слева направо в одной нити такая же, как справа налево в другой. Симметрия подразумевает, что те из них, которые должны быть метилированы, встречаются на обеих цепях ДНК. В результате сайт-мишень может быть полностью метилирован (обе цепи модифицированы), полуметилирован (только одна цепь метилирована) или не метилирован.
Полностью метилированный сайт не подвержен ни рестрикции, ни модификации. Полуметилированный сайт не узнается ферментом рестрикции, но может быть превращен с помощью метилазы в полностью метилированный. У бактерий метилирование, как правило, связано с сохранением имеющегося состояния модификации. Репликация полностью метилированной ДНК ведет к образованию полуметилированной ДНК. Вероятно узнавание полуметилированных сайтов представляет собой обычный этап функционирования метилазы in vivo.
Неметилированный сайт-мишень пред¬ставляет собой субстрат либо для рестрикции, либо для модификации in vitro. В клетке немодифицированная ДНК с большей вероятностью рестрицируется. Реакция разрезания осуществляется в две ступени. Сначала разрезается одна цепь ДНК, а затем рядом разрезается другая. В областях, прилегающих с каждой стооны к сайту разрезания, может иметь место экзонуклеотическая деградация. Происходит эффективный гидролиз АТФ, роль которого еще не выяснена.
Каким образом фермент узнает один сайт, а разрезает другой, достаточно удаленный? Важно отметить, что белок никогда не отделяется от молекулы ДНК, с которой он первоначально связался. Если фермент инкубировать со смесью модифицированной и немодифицированной ДНК, он предпочтительно разрезает немодифицированную ДНК. Следовательно, узнавая сайт связывания, белок не отделяется от неметилированной ДНК для того, чтобы найти сайт разрезания.
Существуют две альтернативные модели, объясняющие взаимосвязь между сайтами узнавания и разрезания: в соответствии с одной из них движется фермент, согласно другой модели, перемещается ДНК. Если движется фермент, то его перемещение вдоль ДНК будет продолжаться до тех пор, пока он не сделает выбор сайта разрезания. Если же движется ДНК, то фермент остается прикрепленным в сайте узнавания, а ДНК протаскивается через второй сайт связывания на ферменте, и это продолжается до тех пор, пока фермент не достигает области разрезания (пока не охарактеризованной). Получены электронно-микроскопические данные, свидетельствующие, что фермент вызывает образование петли в ДНК и остается, по-видимому, связанным с сайтом узнавания после разрезания; эти данные подтверждают вторую модель.
1.1.2 Построение рестрикционных карт
Ферменты рестрикции стали эффективным инструментом исследования. Они позволяют превращать молекулы ДНК очень большого размера в набор фрагментов длиной от нескольких сотен до нескольких тысяч оснований. С помощью метода электрофореза в агарозном геле (см. раздел 1) фрагменты ДНК, различающиеся по размеру, можно легко разделить, а затем исследовать каждый фрагмент отдельно.
Короткие фрагменты мигрируют намного быстрее, чем длинные. При сравнительно высокой концентрации агарозы большие фрагменты вообще не могут проникнуть в гель. В процессе миграции рестрикционные фрагменты не деградируют, их можно элюировать (вымывать) в виде биологически активных двуцепочечных молекул. При окрашивании гелей красителями, связывающимися с ДНК, выявляется набор полос, каждая из которых отвечает рестрикционному фрагменту, молекулярную массу которого можно определить, проведя калибровку с помощью ДНК с известными молекулярными массами.
Сравнение размеров фрагментов ДНК, полученных после обработки определенного участка генома набором рестрицирующих нуклеаз позволяет построить рестрикционную карту, на которой указано положение каждого сайта рестрикции относительно других участков.
Молекулу ДНК длиной 5000 пар нуклеотидов (п. н.). обрабатывают отдельно рестриктазами А и В. Фрагменты разделяют электрофорезом. Фермент А разрезал ДНК на 4 фрагмента размером 2100, 1400, 1000 и 500 п. н. Обработка рестриктазой В дала 3 фрагмента: 2500, 1300 и 1200 п. н. (рис. 37). Для определения расположения сайтов рестрикции этих ферментов на следующем этапе применяют процедуру двойного расщепления – обрабатывают ДНК двумя эндонуклеазами. Обработка изучаемого фрагмента одновременно двумя рестриктазами дала 6 фрагментов: 1900, 1000, 800, 600, 500, 200 п. н.
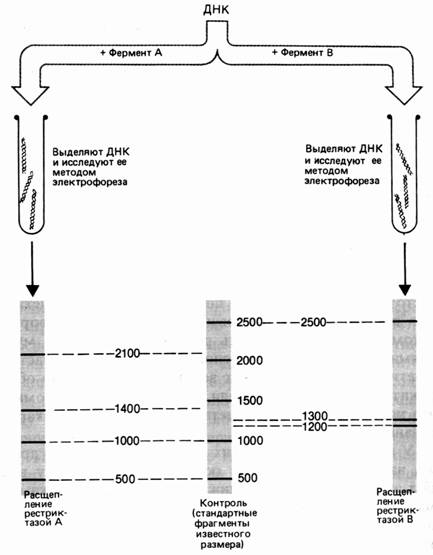
Рис. 1. Результаты электрофореза после обработки фрагмента ДНК разными рестриктазами
Наиболее полный вариант – элюировать каждый фрагмент, образующийся в результате расщепления одной рестриктазой, а затем обработать его второй. Смесь фрагментов, полученных после такой обработки, также анализируют с помощью электрофореза. В нашем примере были получены следующие результаты:
Обработка каждого из 4-х А-фрагментов рестриктазой В
2100 - 1900 и 200,
1400 - 800 и 600,
1000 - 1000 (изменений нет)
500 - 500 (изменений нет)
Обработка каждого из 3-х В-фрагментов рестриктазой А
2500 - 1900 и 600
1300 - 800 и 500
1200 - 1000 и 200
Анализ полученных результатов показывает, что каждый из ферментов, полученный при расщеплении А-фрагментов рестриктазой В можно обнаружить в образцах, полученных при расщеплении В-фрагментов рестриктазой А. Ключом к рестрикционному картированию являются перекрывающиеся фрагменты. Такими в рассматриваемом примере являются В-фрагмент 2100 и А-фрагмент 2500. При обработке другой рестриктазой они дают фрагмент 1900.
Из данных о расщеплении этих фрагментов мы предполагаем, что с одной стороны на расстоянии 200 п. н. от фрагмента 1900 находится следующий А-сайт, а с другого конца, на расстоянии 600 п. н. – следующий В-сайт (рис. 38). При обработке двумя эндонуклеазами фрагмент 200 п. н. образуется 1 раз, при обработке рестриктазой А из В-фрагмента 1200, т. е. фрагмент 1200 лежит слева. Остается определить, как продолжается карта вправо. Очевидно, это А-фрагмент 1400, так как он рассечен рестриктазой В на фрагменты 600 и 800. Вправо от фрагмента 2500 следует отложить, очевидно, фрагмент 1300. Тогда логично наличие А-фрагмента 500 и деления В-фрагмента 1300 рестриктазой А на 800 и 500.
При построении рестрикционных карт обычно используют несколько рестриктаз, поэтому приходится анализировать сложные соотношения между фрагментами, полученными при действии разных ферментов. Для упрощения процедуры картирования можно применять неполное расщепление. В определенных условиях рестриктаза узнает и расщепляет не все сайты в молекуле ДНК. Например, при частичном расщеплении ДНК ферментом А могут образовываться фрагменты 3100 п. н., 1400 п. н. и 500 п. н. Сопоставив их с данными полного расщепления (2100, 1400, 1000 и 500), можно сразу поставить рядом 2100 и 1000 (фрагмент 3100). А получив фрагмент 3500 – расположить рядом 2100 п. н. и 1400 п. н.
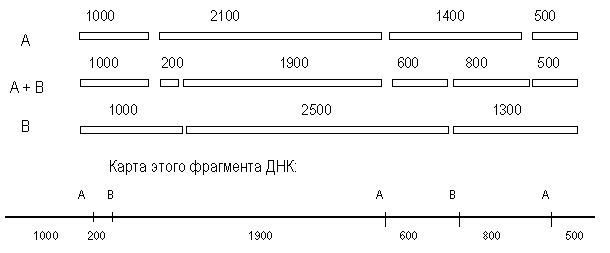
Рис. 2. Анализ фрагментов рестрикции и карта фрагмента ДНК
Другой прием – введение радиоактивной концевой метки. Концевые фрагменты определяются в этом случае по включению метки. Можно также сопоставить фрагменты путем гибридизации нуклеиновых кислот. Перекрывающиеся фрагменты (в данном случае 2100 и 2500) будут гибридизоваться.
Первая карта была получена для вируса SV40 (обезьяний вирус, вызывающий злокачественную трансформацию), содержащего 5423 пары оснований. Использовали рестриктазу Hind-II, расщепляющую кольцевую ДНК вируса на 11 фрагментов. Порядок их расположения в ДНК был установлен путем исследования наборов фрагментов, образующихся по мере того, как расщепление доходит до конца. Первый разрыв превращал кольцевую молекулу в линейную, которая затем расщеплялась на все меньшие и меньшие фрагменты. Исследовали вначале наборы перекрывающихся фрагментов, а затем продукты полного расщепления. Таким образом была получена рестрикционная карта кольцевой вирусной ДНК, на которую были нанесены сайты расщепления рестриктазой. Повторив подобные эксперименты с другой рестриктазой можно получить более подробную карту, где отмечено много сайтов рестрикции.
Располагая такой информацией, можно идентифицировать на ДНК биологически важные участки. Поскольку рестрикционная карта отражает расположение определенной последовательности нуклеотидов в данном участке, сравнение таких карт для двух или более родственных генов позволяет оценить гомологию между ними. Анализируя рестрикционные карты, можно сравнивать определенные участки ДНК разных видов животных без определения их нуклеотидной последовательности. Таким образом, например, было установлено, что хромосомные участки, кодирующие цепи гемоглобина у человека, орангутанга и шимпанзе сохранились в практически неизменном виде в течение последних 5 - 10 млн. лет (с тех пор как виды дивергировали).
Метод рестрикционного картирования позволяет увидеть крупные генетические изменения, такие как делеции или инсерции. При этом происходит уменьшение или увеличение рестрикционных фрагментов, а также исчезновение или возникновение сайтов рестрикции.
Один из приемов картирования – фингерпринт («метод отпечатков пальцев» или DNA-fingerprint). Он подразумевает использование неупорядоченных и неполных наборов фрагментов, которые являются характеристикой генома, хотя описывает его не полностью.
0 комментариев