Регуляция биосинтеза белков на этапе трансляции
До недавнего времени считалось, что регуляция на уровне трансляции характерна почти исключительно для эукариот, где существует временное и пространственное разобщение транскрипции и трансляции в связи с наличием оболочки ядра и формированием долгоживущих, «защищенных» белками иРНК информосом, открытых А.С. Спириным.
Косвенным указанием на существование регуляции трансляции у прокариот является различие в скорости образования некоторых белков, кодируемых одним и тем же регулоном и транслируемых с общей полицистронной матрицы, как, например, в случае субъединиц РНКП и некоторых рибосомных белков.
Существует потенциальная возможность регуляции скорости трансляции на двух этапах: на этапе биосинтеза и сборки компонентов аппарата трансляциии и на этапе их функционирования.
Регуляция на этапе биосинтеза и сборки компонентов аппарата трансляции
Основными высокомолекулярными компонентами аппарата трансляции являются аминоацил-тРНК-синтетазы, транспортные РНК, рибосомные РНК, информационные РНК, рибосомные белки и белковые факторы трансляции.
Аминоацил-тРНК-синтетазы представляют собой довольно крупные белки, чаще всего мультимерные. Число их видов равно числу природных аминокислот. Уровень всех или большинства АРСаз регулируется координирование и пропорционален скорости роста. Избыток аминокислот не оказывает на синтез АРСаз прямого репрессирующего действия. Значительная часть АРСаз эукариот ассоциирована с полирибосомами и организована в полиферментные комплексы. Поэтому в отношении их действуют регуляторные механизмы, характерные для управления активностью ферментных ансамблей.
Транспортные РНК составляют около 10–15% всей клеточной РНК и кодируются 50–60 генетическими локусами. Биосинтез тРНК проходит промежуточное образование предшественников, которые затем укорачиваются и модифицируются – явление, называемое процессингом, или «созреванием» тРНК. Например, предшественник тирозиновой транспортной РНК у Escherichia coli содержит 129 нуклеотидов, это на 44 нуклеотида больше, чем «зрелая» форма. Такой предшественник можно выделить из температурочувствительного мутанта с дефектом РНКаз. Он подвергается процессингу с участием двух РНКаз: Р и PIII. Необходимо отметить, что РНКаза Р состоит из РНК и белка, причем РНК сама по себе может выполнять функции фермента, катализируя процессинг, тогда как белок только стимулирует ее активность. Таким образом, РНКаза Р является пока не очень многочисленным примером рибозима, т.е. фермента, представляющего собой не белок, а РНК.
Следующим этапом процессинга многих тРНК является модификация оснований, которая осуществляется при помощи большого количества ферментов, поскольку даже одна и та же модификация основания, находящегося в разных участках молекулы тРНК, может осуществляться разными ферментами.
Важно отметить, что в процессе эволюции количество модифицированных компонентов в тРНК возрастало. Это указывает на важную регуляторную роль модифицированных изоакцепторных тРНК. Далее рассмотрим гипотезы, касающиеся возможных механизмов регуляции трансляции, основанных на использовании изоакцепторных тРНК.
Рибосомные РНК у эукариот представлены четыремя типами.» 28S, 18S, 5,8S и 5S, у прокариот тремя: 23S, 16S и 5S. Кроме того, в митохондриях и хлоропластах эукариот содержатся рибосомы, близкие по составу к прокариотическим. Геном Escherichia coli содержит 6–7 оперонов, включающих локусы всех трех рРНК, а также разное число локусов тРНК. В процессе транскрипции синтезируется общий транскрипт, подвергающийся процессингу, который осуществляется в ходе транскрипции, так что общий транскрипт длиной 5600 нуклеотидов может быть выделен только в системе in vitro или у мутантов с дефектом РНКаз. РНКаза III «узнает» двунитевые участки, соответствующие границам локусов транскрибируемых РНК. После действия РНКазы III образуются не «зрелые» молекулы РНК, а их предшественники, которые подвергаются дальнейшему про-цессингу. В случае рРНК он может состоять, например, в метилировании, которое обычно осуществляется после того, как рРНК включена в состав субчастицы рибосомы.
У некоторых эукариот процессинг 28S РНК состоит в удалении «лишнего» фрагмента посредством аутосплайсинга, при котором РНК выполняет роль «псевдофермента», катализирующего собственный процессинг в отсутствие белков-ферментов.
Ступенчатый процессинг позволяет осуществить регуляцию на каждом из этапов биосинтеза и сборки, а указанная организация локусов обеспечивает образование всех рРНК в равных соотношениях.
Синтез рРНК и рибосомных белков регулируется координирование и определяется эффективностью роботы аппарата трансляции: например, при дефиците аминокислот транскрипция локусов, кодирующих рРНК и рибосомные белки, подавляется одновременно. Подробнее об этом будет рассказано в главе, посвященной регуляции скорости роста.
Информационная РНК у прокариот, как правило, полицистронна, а у эукариот – моноцистронна. Процессинг у эукариот состоит в вырезании интронов и в сплайсинге экзонов. Затем происходит метилирование оснований. На конце 5 достраивается специфическая группировка, так называемый «кэп», а на конце 3 последовательность из остатков адениловой кислоты, включающая до 100–400 нуклеотидов. Роль этой последовательности остается неясной. Поступившая в цитоплазму иРНК существует в виде нукле-опротеидного комплекса – информосом и в таком виде может сохраняться длительное время до наступления трансляции.
Для прокариот процессинг иРНК не характерен, хотя в некоторых иРНК архебактерий также обнаружены интроны, а у Escherichia coli полицистронный транскрипт локуса, включающего гены рибосомных белков LI, L7, L12 и гены, содержащие от 5 до 180 остатков адениловой кислоты. Показано, что они наиболее характерны для секретируемых белков.
Практически отсутствуют сведения о регуляции образования белковых факторов трансляции. У прокариот это факторы инициации, элонгации, терминации. У эукариот их больше, они сложнее по строению: только факторов инициации насчитывается свыше восьми, зато фактор терминации всего один.
По полученным за последнее время сведениям, некоторые из этих факторов способны модифицироваться; например, у эукариот может происходить фосфор ил ирование фактора инициации eIF‑2 и фактора элонгации EF‑1, а также ADP‑рибозил ирование фактора EF‑2, что, несомненно, влияет на скорость процесса трансляции. В случае прокариот обнаружено метилирование фактора элонгации EF-Tu Escherichia coii, однако регуляторное значение этого процесса неизвестно.
Регуляция на этапе функционирования аппарата трансляции
Регуляция может осуществляться как на этапе инициации, так и этапе элонгации.
1. Регуляция инициации. У прокариот регуляция инициации трансляции осуществляется путем специфического связывания регуляторных белков с инициаторным районом иРНК, в результате чего трансляция подавляется. Такая регуляция существует при развитии фага MS2, в процессе которого белок оболочки регулирует трансляцию матрицы РНК-репликазы. Аналогичный механизм действует при регуляции трансляции рибосомных белков Escherichia coii. Гены 52 рибосомных белков организованы в 16 оперонов, в ряде случаев они объединены с локусами компонентов РНКП и факторов трансляции, и для поддержания их независимого и координированного синтеза необходим механизм «обратной связи», в результате функционирования которого избыток рибосомных белков подавляет собственную трансляцию. Иногда рибосома обнаруживает разное сродство к инициаторным участкам разных иРНК, благодаря этому инициация может также регулироваться самой рибосомой. Эта особенность бывает постоянной или регулируется специальными факторами, в результате обеспечивается дифференциальная скорость трансляции разных локусов полицистронной матрицы иРНК.
В некоторых специализированных эукариотических клетках двухцепочечная РНК ингибирует трансляцию, вызывая активацию протеинкиназы, фосфорилирующей фактор инициации eIF‑1.
Описана также возможность блокирования трансляции у прокариот путем присоединения к инициаторному участку иРНК особой комплементарной регуляторной РНК.
Наконец, существует потенциальная возможность управления скоростью трансляции у прокариот на этапе инициации за счет изменения доступности инициаторной тРНК, которая является обязательной для инициации.
2. Регуляция элонгации. Создается впечатление, что в биосинтезе биополимеров определяющей стадией является инициация, а этап элонгации регулируется в меньшей степени или совсем не регулируется. Тем не менее существует несколько потенциальных возможностей регуляции этапа элонгации в процессе трансляции. Одна из таких возможностей состоит в избирательной трансляции матрицы иРНК за счет специфического набора изоакцепторных тРНК.
Для 20 природных аминокислот существует 61 значащий кодон. По правилу «неоднозначного соответствия» Крика, для их транслирования достаточно около 30 антикодонов. Однако в клетках значительно больше тРНК, некоторые из них специфичны только к одному из кодонов. Если данная аминокислота в данном белке кодируется именно таким кодоном, то наличие или отсутствие соответствующей специфической тРНК будет определять возможность трансляции данной матрицы. В свою очередь, в клетке набор изоакцепторных тРНК может очень тонко регулироваться. Показано, например, что на разных фазах роста Escherichia coli или при переходе от хемотрофного к фототрофному типу питания у Rhodobacter sphaeroides наблюдается резкое изменение набора тРНК. Таким образом, данный способ регуляции можно рассматривать как еще один пример системной регуляции.
Другой пример относится к развитию фагов, в процессе которого наряду с изменением процесса транскрипции блокируется трансляция иРНК организма-хозяина. В некоторых случаях это связано с модификацией рибосом, перестающих «узнавать» иРНК хозяина, однако у эукариот это обусловлено модификацией факторов элонгации, для чего имеются специальные ферменты, осуществляющие фосфор ил ирование или ADP‑рибозилирование, что резко уменьшает сродство данных белков к рибосомам.
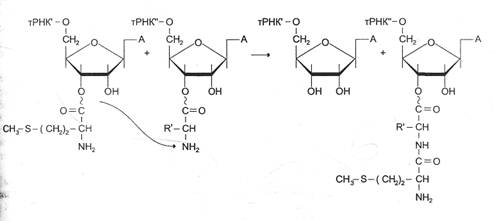
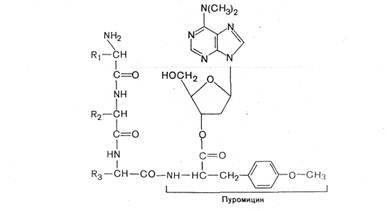
Что касается терминации трансляции, то специальные механизмы регуляции в этом случае не обнаружены. Однако необходимо отметить возможность «проскакивания» рибосомы через некоторые «слабые» терминирующие кодоны, что, с одной стороны, позволяет рибосоме транслировать полицистрон-ные матрицы без диссоциации от иРНК, а с другой – обеспечивает образование небольшого количества более длинных полипептидов, могущих выполнять важную функциональную роль.
Регуляция биосинтеза белков путем посттрансляционной модификации
Посттрансляционная модификация белков менее распространена, чем процессинг РНК. Тем не менее известны случаи, когда при развитии некоторых вирусов трансляция полицистронной матрицы приводила к образованию общей полипептидной цепи, разрезаемой в дальнейшем на индивидуальные белки специфическими протеиназами. Кроме того, широко известен процессинг ряда ферментов, превращающий их неактивные формы в активные.
У прокариот наиболее распространенным видом процессинга белков является удаление «сигнального» пептида из молекул секре-тируемых белков. Такие белки и ферменты содержат на NH2-Komje гидрофобный пептид из 15–30 аминокислот, который необходим для транслокации белка через цитоплазм атическую мембрану в процессе его синтеза. После завершения транслокации «сигнальный» пептид удаляется специальной «сигнальной» пептидазой.
К группе процессов посттрансляционной модификации можно отнести ферментативное присоединение коферментов к готовой молекуле апофермента, а также, с некоторой долей условности, и формирование мультимерных белков из нескольких полипептидных цепей с участием белков-шаперонов.
Регуляция круговорота белков путем избирательного протеолиза
Количественный и качественный состав белков в клетке может регулироваться не только на этапе их биосинтеза, но и на этапе деградации. В нормально растущих клетках бактерий за клеточный цикл распадается несколько процентов существующих белков. Более высокая скорость круговорота белка характерна для термофильных организмов. В условиях голодания скорость распада возрастает в несколько раз. Протеолиз клеточных белков выполняет две важные функции. Во-первых, расщепляются ненужные в данный момент ферменты, функционирование которых могло бы вызвать дисбаланс метаболизма, и восполняется ресурс аминокислот. Во-вторых, ликвидируются «ошибочные» белки, возникающие в результате сбоя в биосинтетических процессах или за счет мутаций.
Протеолиз играет особенно важную роль в процессах клеточной дифференцировки, о чем, например, свидетельствует утрата способности к спорообразованию при дефекте синтеза протеиназ. Возможны два основных типа протеолиза: АТР-независимый иАТР-зависимый. Первый активируется в условиях голодания и не требует затраты энергии; второй действует постоянно и весьма избирательно. В эти системы, вероятно, включаются разные ферменты, так как некоторые ингибиторы протеиназ подавляют первый процесс и не влияют на второй. АТР-зависимый протеолиз, по-видимому, включает стадию «узнавания» аномального белка и введение в него метки, которой является специальный белковый агент – убихитин, после чего меченый белок подвергается деградации протенназами.
Механизм «узнавания» аномальных или нефункциональных белков неизвестен, скорее всего, важную роль в нем играют особенности третичной структуры белков, и замена даже одной аминокислоты сильно снижает устойчивость белка к внутриклеточному протеолизу. Последнее обстоятельство может существенно мешать получению микроорганизмов-сверхпродуцентов, у которых повышенное образование целевого продукта обусловлено мутациями по соответствующим ферментам. Такие ферменты будут восприниматься системой узнавания как аномальные и подвергаться протеолизу, что тормозит биосинтетические процессы, а иногда и рост микроорганизма. Специфический протеолиз может дополнять регуляцию по механизму катаболитной репрессии. Например, у некоторых дрожжей глюкоза не только репрессирует синтез определенных ферментов, но и стимулирует их протеолитическую деградацию, по-видимому, за счет индукции или активации соответствующей протеиназы. Решающую роль играет протеолиз в так называемой SOS‑регуляции, т.е. активации SOS‑регулона, включающего около 20 генов, которые индуцируются в ответ на некоторые повреждения ДНК и образуют продукты, участвующие в ее репарации. Среди этих продуктов присутствует белок RecA, участвующий в ряде клеточных процессов является более «быстродействующим» механизмом и раньше откликается на изменение внешних условий, чем регуляция биосинтеза этих посредников. Однако, как мы уже отмечали, оба уровня регуляции необходимы для координированного управления биохимическими процессами в клетке. В свою очередь, процессы регуляции активности белковых посредников можно разделить на две большие группы: регуляция активности путем обратимой ковалентной модификации посредника и регуляция активности без ковалентной модификации посредника.
Регуляция активности белковых посредников путем их ковалентной модификации
Отличие этого механизма регуляции от посттрансляционной модификации состоит в обратимости процесса и отсутствии изменения длины полипептидной цепи. Кроме того, и это особенно важно отметить, обе формы – модифицированная и немодифицированная – активны, хотя и различаются по величине активности н/или по регуляторным свойствам.
У эукариот самым распространенным способом модификации является фосфорилироеание. Один из наиболее изученных примеров – процесс синтеза и распада гликогена.
Фосфор ил ирование ферментов, участвующих в синтезе и распаде гликогена, осуществляется киназами, которые сами активируются сАМР. Как уже отмечалось, концентрация сАМР в клетке обратно пропорциональна концентрации АТР. Следовательно, потребность в энергии приводит к фосфори-лированию указанных ферментов, т.е. к стимуляции гидролиза гликогена и торможению его синтеза. Дополнительная стимуляция гидролиза достигается за счет активирующего эффекта AMP, который накапливается при снижении энергетического заряда клетки. Напротив, при накоплении глюкозо‑6‑фосфата, что свидетельствует об активном протекании энергетических процессов, гидролиз гликогена тормозится.
У прокариот, как показано в последнее время, модификация белков путем их фосфорилирования также распространена достаточно широко. Так, в процессе инициации спорообразования у бацилл активируется транскрипция ряда генов, кодирующих белки, часть которых является протеинкиназами, а часть – акцепторами фосфата. Один из последних белков способен связываться с ДНК и, по-видимому, является регулятором транскрипции. Фосфорилирова7 ние влияет на его регуляторные свойства. Этот же белок необходим для развития у клеток Bacillus subtilis состояния компетентности. Путем фосфорилирования регулируется также активность некоторых белков у Khizobium, участвующих в фиксации азота, а также в транспорте ди‑и трикарбоновых кислот. Регуляция транспорта Сахаров путем фосфорилирования компонентов фосфотрансферазной системы обнаружена у Escherichia coli. Вообще же у этой бактерии найдено около 170 белков, способных фосфорилироваться.
Однако наиболее изученным примером регуляции путем ковалентной модификации является аденилирование и уридилирование ферментов в системе регуляции активности глутаминсинтетазы.
Указанная регуляция активности ГС дополняется регуляцией на уровне биосинтеза фермента: немодифицированный белок РП через посредство других специальных белков подавляет транскрипцию локусов ГС. В свою очередь, эти белковые регуляторы могут фосфорилироваться с участием специфических протеинкиназ и изменять свою регуляторную активность.
Все эти события – яркий пример каскадной регуляции – наиболее эффективного механизма регуляции сложных метаболических путей, каким и является, в частности, азотный метаболизм.
Регуляция активности белковых посредников путем нековаленткого взаимодействия с эффекторами
1. Взаимодействие с субстратами. Ферменты, активность которых регулируется субстратом, должны иметь несколько активных центров, сходных по природе и взаимодействующих между собой. Здесь возможны два случая:
а) присоединение первой молекулы субстрата облегчает присоединение последующих молекул, и скорость реакции растет по экспоненциальному закону. График зависимости начальной скорости реакции от концентрации субстрата имеет S‑образную форму;
б) присоединение первой молекулы субстрата затрудняет присоединение последующих молекул.
Аналогичные механизмы регуляции действуют при трансмембранном транспорте некоторых субстратов.
2. Взаимодействие с продуктами и другими эффекторами, отличными от субстратов. Ферменты, активность которых регулируется по этому механизму, должны иметь различающиеся по природе активные центры: каталитический и регуляторный. Эти центры обычно размещены на разных субъединицах фермента, причем связывание эффектора с регуляторным центром влияет на конформацию каталитического центра и изменяет сродство к субстрату, которое, как правило, снижается. При этом возможны разные обстоятельства:
а) в анаболических процессах конечный продукт метаболического пути, накапливаясь выше определенного уровня, подавляет свой биосинтез, ингибируя активность первого фермента данного пути;
б) в катаболических путях метаболизма отрицательными эффекторами часто служат соединения, являющиеся аккумуляторами энергии, а другие компоненты аденилатной системы могут выступать в качестве положительных эффекторов. Таким образом, активность данных ферментов зависит от «энергетического заряда» клетки;
в) амфиболические ферменты могут регулироваться с помощью обоих механизмов;
г) в разветвленных биосинтетических путях подавление одним из конечных продуктов активности фермента, катализирующего начальные этапы процесса, приводило бы к дефициту других продуктов данного пути. Поэтому необходима особая организация регуляторных процессов. Существуют две основные возможности.
Во-первых, образование изоферментов, катализирующих начальную стадию пути, активность каждого из которых избирательно подавляется только одним из конечных продуктов. Примером может служить биосинтез ароматических аминокислот у Escherichia coli, в котором конечные продукты – тирозин, триптофан и фенилаланин – подавляют каждый активность одной из альдолаз, катализирующих первую реакцию пути. Во-вторых, использование ферментов, имеющих несколько взаимодействующих регуляторных центров, каждый из которых специфичен только для одного из эффекторов. По отдельности они не оказывают существенного влияния на активность фермента, а при их совместном действии активность подавляется. Это так называемое согласованное, или мультивалентное ингибирование. Например, активность аспартаткиназы у Escherichia coli подавляется только сочетанием лизина, метионина и лейцина. Для глутаминсинтетазы обнаружено восемь кумулятивных эффекторов: аланин, глицин, гистидин, триптофан, ЦТР, AMP, карбамоилфосфат, глюкозамин‑6‑фосфат.
Регуляция активности белковых посредников путем пространственного разобщения и взаимодействия с мембранами
Механизмы первого типа более распространены у эукариот в связи с локализацией ферментов в субклеточных органеллах: митохондриях, лизосомах и т.д. Однако и в клетках прокариот возможны определенные виды компартментации:
а) часть ферментного аппарата прокариот локализована в периплазматическом пространстве. Таким образом, создается возможность регуляции активности ферментов путем управления скоростью проникновения в «отсек» субстратов или выхода из него продуктов;
б) ферменты, катализирующие серию последовательных реакций, могут формировать «ансамбль», локализованный либо в цитоплазме, либо в цитоплазматической мембране. Продукты, образуемые на предыдущей стадии, «подхватываются» последующим ферментом без освобождения в среду.
Для регуляторного действия конечного продукта доступен только последний фермент, но он может передавать эффект на предыдущие ферменты за счет кооперативных взаимодействий. Существует представление о «метаболоне», т.е. комплексе ферментов, закрепленных на «подложке». Каталитические свойства в этом случае проявляются внутри ансамбля, а подложка реагирует на регуляторное действие эффекторов. Например, ни один из ферментов может не реагировать на данный эффектор, но в ансамбле, за счет конформационных изменений подложки, они становятся к нему чувствительными.
Важную роль в регуляции активности ферментов может играть их взаимодействие с мембранами. В мембранно-связанном состоянии физико-химические свойства ферментов изменяются. Это явление называется аллотопия. Гидрофобные взаимодействия мембранных липидов и белков могут переводить последние в неактивное состояние, а электростатические взаимодействия, напротив, вызывать активацию белков. В свою очередь, сила электростатического взаимодействия липидов и белков зависит от внутриклеточной концентрации электролитов, а следовательно, от состава среды, окружающей клетку, и от физиологического статуса последней. Таким образом, для регуляции ферментов по этому механизму существуют широкие возможности, хотя конкретные механизмы в силу труднопреодолимых методических препятствий пока изучены мало.
Похожие работы

... процессы репликации ДНК и деления клетки. Реальные механизмы регуляции репликации еще не расшифрованы. У эукариот определенную роль в регуляции репликации, по-видимому, играют гистоны. 4. Транскрипция генетической информации Этот этап биосинтеза белков состоит в «переписывании» информации, закодированной в ДНК, на олигонуклеотидную последовательность информационной РНК и осуществляется ...
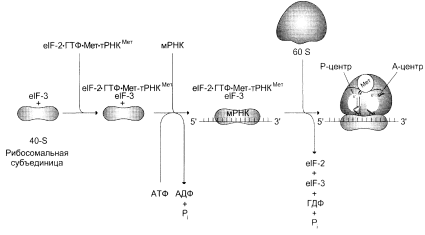
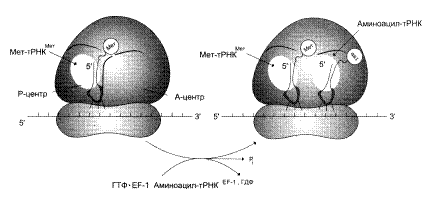
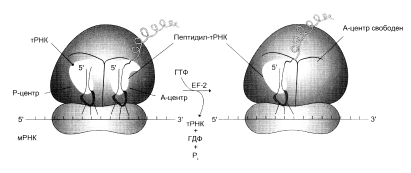
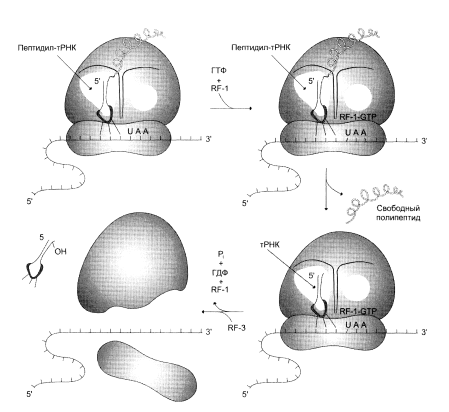
... и их переносе на рибосомы, более 60 макромолекул, входящих в состав 70S или 80S рибосом, и около 10S макромолекул, принимающих непосредственное участие в системе трансляции. Белковый синтез, или процесс трансляции, может быть условно разделен на 2 этапа: активирование аминокислот и собственно процесс трансляции. Второй этап матричного синтеза белка, собственно трансляцию, протекающей в рибосоме, ...
... , вызывает уменьшение количества синтезируемых ферментов. Это последнее явление получило название репрессии синтеза ферментов. Оба явления — индукция и репрессия — взаимосвязаны. Согласно теории Жакоба и Моно в биосинтезе белка у бактерий участвуют по крайней мере три типа генов: структурные гены, ген-регулятор и ген-оператор. Структурные гены определяют первичную структуру синтезируемого белка. ...
... организма, в том числе состав и структура синтезируемых белков. Первичная структура ДНК представляет собой определенную последовательность мононуклеотидов, каждые три из которых носят названия триплет и кодируют вполне определенную аминокислоту. Синтез белка можно условно разделить на три этапа. 1 этап – синтез информационной РНК - транскрипция. Синтез информационной РНК происходит в ядре ...
0 комментариев