Работу выполнила студентка Самойлова Р.Н., Л-212 б группы II курса.
Башкирский государственный медицинский университет
Кафедра Биологической химии с курсом органической химии
Уфа 2004
Введение
Биосинтез белка
Одной из задач современной биологии и ее новейших разделов – молекулярной биологии, биоорганической химии, физико-химической биологии – является расшифровка механизмов синтеза молекулы белка, содержащей сотни, а иногда и тысячи остатков аминокислот. Механизм синтеза должен обладать точной кодирующей системой, которая автоматически программирует включение каждого аминокислотного остатка в определенное место полипептидной цепи Кодирующая система определяет первичную структуру, а вторичная и третичная структуры белковой молекулы определяются физико-химическими свойствами и химическим строением аминокислот.
Первоначальные представления, согласно которым синтез белка могут катализировать те же протеолитические ферменты, что и вызывающие его гидролиз, но путем обратимости химической реакции, не подтвердились. Оказалось, что синтетические и катаболические реакции протекают не только различными путями, но и в разных субклеточных фракциях. Не подтвердилась так же гипотеза о предварительном синтезе коротких пептидов с их последующим объединением в единую полипептидную цепь. Более правильным оказалось предположение, что для синтеза белка требуются источники энергии, наличие активированных свободных аминокислот и несколько видов нуклеиновых кислот.
В современные представления о механизме синтеза белка большой вклад внесли советские биохимики. Так, в лаборатории А. Е. Браунштейна было впервые указано на участие АТФ в синтезе квазипептидных связей. В. Н. Ореховичем еще 50-е годы было показано, что перенос аминоцильных или пептидильных группировок на NH2 группу аминокислот может осуществляться не только с амидной или пептидной, но и со сложноэфирной связи. Как будет показано ниже, именно этот механизм лежит в основе реакции транспептидирования в 50S рибосоме в стадии элонгации синтеза белка.
Значительно позже были получены доказательства, что в синтезе белка, протекающем в основном в цитоплазме, решающую роль играют нуклеиновые кислоты, в частности ДНК. После того как было установлено, что ДНК является носителем и хранителем наследственной информации, был поставлен вопрос о том, каким образом эта генетическая информация, записанная(зашифрованная) в химической структуре ДНК, трансформируется в фенотипические признаки и функциональные свойства живых организмов, передающиеся по наследству. В настоящее время можно дать однозначный ответ на этот вопрос: генетическая информация программирует синтез специфических белков, определяющих в свою очередь специфичность структуры и функции клеток, органов и целостного организма. В природе, как известно, существуют два типа биополимерных макромолекул, так называемые неинформативные биополимеры и информативные биополимеры, несущие первичную генетическую информацию и вторичную генетическую, точнее фенотипическую информацию. Эти общие представления могут быть выражены следующей последовательностью событий(поток информации):
ДНК®РНК®Белок®Клетка®Организм
Биосинтез белка, хотя непосредственно и регулируется рибонуклеиновыми кислотами, опосредованно связан с контролирующим влиянием ДНК ядра и что РНК сначала синтезируется в ядре, затем поступает в цитоплазму, где выполняет роль матрицы в синтезе белка. Полученные значительно позже экспериментальные данные подтвердили гипотезу о том, что основной функцией нуклеиновых кислот является не только хранение генетической информации, но и реализация этой информации путем программированного синтеза специфических белков.
Однако в этой последовательности ДНК®РНК®Белок недоставало сведений о том, каким образом происходят расшифровка наследственной информации и синтеза специфических белков, определяющие многообразие признаков живых существ. В настоящее время выяснены основные процессы, посредством которых осуществляется передача наследственной информации: они включают репликацию, т. е. Синтез ДНК на матрице ДНК, транскрипцию, т. е. Перевод языка и типа строения ДНК на молекулу РНК, и трансляцию – процесс, в котором генетическая информация, содержащаяся в молекуле мРНК, направляет синтез соответствующей аминокислотной последовательности в белке. Многие тонкие механизмы транскрипции окончательно не выяснены.
Получены экспериментальные доказательства наличия ДНК также в митохондриях. Она не гомологичная и не комплементарна ядерной ДНК. Предполагается, что митохондриальная ДНК кодирует синтез части структурных белков самих митохондрий.
Значительный вклад в современные представления о месте, факторах и механизме синтеза белка внесли исследования Т. Касперсона, П. Берга, П. Замечника, С. Очоа, А. А. Баева, А. С. Спирина и др.
Генетический код и его свойства
Необходимость кодирования структуры белков линейной последовательности нуклеотидов мРНК и ДНК продиктованы тем, что в ходе трансляции:
Нет соответствия между числом номеров в матрице мРНК и продукте – синтезируемом белке;
Отсутствует структурное сходство между мономерами РНК и белка.
Это исключает комплиментарное взаимодействие между матрицей и продуктом – принцип, по которому осуществляется построение новых молекул ДНК и РНК, в ходе репликации и транскрипции.
Отсюда становится ясно, что должен существовать «словарь», позволяющий выяснить, какая последовательность нуклеотидов мРНК обеспечивает включение в белок аминокислот в заданной последовательности. Этот «словарь» получил название генетического, биологического, нуклеотидного или аминокислотного кода. Он позволяет шифровать аминокислоты, входящие в состав белков, с помощью определенной последовательности нуклеотидов в ДНК и мРНК. Для него характерны определенные свойства.
Триплетность.Одним из основных вопросов при выяснении свойств кода был вопрос о числе нуклеотидов, которое должно определять включение в белок одной аминокислоты. Сразу было понятно, что это число не может быть равным 1 или 2, так как в этом случае количество кодирующих элементов будет недостаточно для шифрования 20 аминокислот в белках. Число кодирующих последовательностей из четырех нуклеотидов по три равно 43=64, что более чем в 3 раза превышает минимальное количество, которое необходимо для кодирования 20 аминокислот. В дальнейшем было установлено, что кодирующими элементами в шифровании аминокислотной последовательности действительно являются тройки нуклеотидов или триплеты, которые получили название «кодоны».
Смысл кодонов
Смысл кодонов стал понятен в 60-х г. XX столетия, когда, используя безклеточную систему синтеза белков и синтетические полирибонуклеотиды и заданной последовательностью нуклеотидов в качестве матрицы, М . Ниренберг и Г. Маттей синтезировали полипептиды определенного строения. Так, на матрице поли-У, состоящей только из остатков УМФ, был получен полифенилаланин, а на матрице поли-Ц –полипролин. Из этого следовало, что триплет – UUU кодирует Фен, а триплет -ССС – Про.
В последующих экспериментах использовали смешанные синтетические полирибонуклеотиды с известным составом. В результате этой работы удалось установить, что из 64 кодонов включения аминокислот в синтезирующуюся полипептидную цепь шифрует 61 триплет, а 3 остальных UAA, UAG, UGA не кодируют включение в белок аминокислот и первоначально были названы бессмысленными, или нонсенкодоном. Однако в дальнейшем было показано, что эти триплеты сигнализируют о завершении трансляции, и поэтому их стали называть терминируюшими, или стоп-кодонами.
Кодоны мРНК и триплеты нуклеотидов в кодирующей нити ДНК с направлением от 5¢ к 3¢ - концу имеют одинаковую последовательность азотистых оснований, за исключением того, что в ДНК вместо урацила (U), характерного для мРНК, стоит тимин (Т).
Специфичность
Каждому кодону соответствует только одна определенная аминокислота. В этом смысле генетический код строго однозначен.
Выраженность
В мРНК и ДНК имеет смысл 61 триплет, каждый из которых кодирует включение в белок одну из 20 аминокислот. Из этого следует, что в информационных молекулах включения в белок одной и той же аминокислот определяет несколько кодонов. Это свойство биологического кода получило название вырожденности.
У человека одним кодоном зашифрованы только 2 аминокислоты – Мет и Три, тогда как Лей, Сер и Арг – шестью кодонами, а Ала, Вал, Гли, Про, Тре – четырьмя кодонами.
Избыточность кодирующих последовательностей – ценнейшее свойство когда, так как она повышает устойчивость информационного потока к неблагоприятным воздействиям внешней и внутренней среды. При определении природы аминокислоты, которая должна быть заключена в белок, третий нуклеотид в кодоне не имеет столь важного значения, как первые два. Для многих аминокислот замена нуклеотида третьей позиции кодона не сказывается на его смысле.
Линейность записи информации
В ходе трансляции кодоны мРНК «читаются» с фиксированной стартовой точки последовательно и не перекрываются. В записи информации отсутствуют сигналы, указывающие на конец одного кодона и начала следующего.
Кодон AUG является инициирующим и прочитывается только в начале, так и в других участках мРНК как Мет. Следующие за ним триплеты читаются последовательно без каких либо пропусков вплоть до стоп-кодона, на котором синтез полипептидной цепи завершается.
Универсальность
До недавнего времени считалось, что код абсолютно универсален, т. е. смысл кодовых слов одинаков для всех изученных организмов: вирусов, бактерий, растений, земноводных, млекопитающих, включая человека. Однако позднее стало известно одно исключение, казалось, что митохондриальная МРНК содержит 4 триплета, имеющих другое значение, чем в мРНК ядерного происхождения. Так, в мРНК митохондрий триплет UGA кодирует Три, AUA –Мет, а AGA и AGG причитываются как дополнительные стоп-кодоны.
Колинеарность гена и продукта
У прокариотов обнаружено линейное соответствие последовательности кодонов гена и последовательности аминокислот в белковом продукте, или, как говорят, существует колинеарность гена и продукта.
У эукариотов последовательности оснований в гене, колинеарные аминокислотной последовательности в белке, прерываются интронами. Поэтому в эукариотических клетках аминокислотная последовательность белка колинеарна последовательности экзонов в гене или зрелой МРНК после постранскрипционного удаления интронов.
Основные компоненты белоксинтезирующей системы
Для синтеза полипептидной цепи необходимо большое количество компонентов, совместное и согласованное взаимодействие приводит к образованию белка.
Аминокислоты
Все 20 аминокислот, входящих в структуру белков организма человека, должны присутствовать в достаточном количестве. Это требование прежде всего относится к незаменимым (т. е. не синтезирующимся в организме) аминокислотам, так как недостаточное снабжение клетки хотя бы одной незаменимой аминокислотой приводит к снижению, а иногда и полной остановке синтеза белка на кодоне, требующем включения этой аминокислоты в белок.
Транспортная РНК
В лаборатории Хогланда было выяснено, что при инкубации 14С –аминокислоты с растворимой фракцией цитоплазмы в присутствии АТФ и последующим добавлением трихлоруксусной кислоты в образовавшемся белковом осадке метка не открывается. Было сделано заключение, что меченая аминокислота не включается в белковую молекулу. Метка оказалась связанной ковалентно с РНК, содержащейся в белковом фильтрате. Показано, что РНК, к которой присоединяется меченая аминокислота, имеет небольшую молекулярную массу и сосредоточена в растворимой фракции, поэтому ее сначала назвали растворимой, а позже адаптерной или транспортной РНК. На долю тРНК приходится около 10-15% общего количества клеточной РНК. К настоящему времени открыто более 60 различных тРНК. Для каждой аминокислоты в клетке имеется по крайней мере одна специфическая РНК( для ряда аминокислот открыто более одной, в частности для серина – 5 разных тРНК, для лизина и глицина – по 4 разных тРНК, хотя и в этом случае каждая тРНК связана со специфической аминоацил–тРНК-синтетазой). Молекулярная масса большинства тРНК колеблется от 24000 до 29000 Да. Они содержат от 75 до 85 нуклеотидов. Аминокислоты присоединяются к свободной 3¢-ОН-группе концевого мононуклеотида, представленного во всех тРНК АМФ, путем образования эфирной связи. Интересно, что почти все тРНК обладают не только индивидуально сходными функциями, но и очень похожей трехмерной структурой.
Установлена первичная структура почти всех 60 открытых тРНК; знание последовательности нуклеотидов и, следовательно, состава тРНК дало в руки исследователей много ценных сведений о биологической роли отдельных компонентов тРНК. Общей для тРНК оказалась также нативная конформация, установленная методом рентгеноструктурного анализа и названная первоначально названная конформацией клеверного листа; на самом деле эта конформация имеет неправильную, Г-образную, форму.
Определение структуры тРНК позволило выявить ряд отличительных участков; так, 3¢-гидроксильном конце располагается одинаковая для всех тРНК последовательность триплета ЦЦА –ОН, к которой присоединяется посредством эфирной связи специфическая аминокислота. Связывание в основном происходит через 3¢-ОН-группу концевого аденилового нуклеотида, хотя получены доказательства возможности присоединения аминокислоты и через 2¢-ОН-группу. Тимидин-псевдоуридин-цитидиловая петля, по-видимому, обеспечивает связывание аминоацил-тРНК с поверхностью рибосомы. Имеется кроме того, добавочная петля, состав которой варьирует у разных типов молекул тРНК; ее назначение неизвестно. Дигидроуридиловая петля, с другой стороны, оказалась необходимой как сайт (место) для узнавания специфическим ферментом –аминоацил-тРНК-синтетазой. Имеется также антикодоновая петля, несущая триплет, названный антикодоном, и расположенная на противоположной стороне от того конца, куда присоединяется аминокислота. Антикодон является антипараллельными в своей комплементарности.
Тщательный анализ нуклеотидной последовательности разных тРНК показал, что все они содержат одинаковый 5¢-концевой нуклеотид – ГМФ со свободной 5¢-фосфатной -группой. Адапторная функция молекул тРНК заключается в связывании каждой молекулы тРНК со своей специфической функциональной аминокислотой. Но поскольку между нуклеиновой кислотой и специфической функциональной группой аминокислоты не существует соответствия и сродства, эту функцию узнавания должна выполнять белковая молекула, которая узнает как молекулу специфической тРНК, так и специфической аминокислоты.
Матричная РНК
Выше было указано на необходимость участия предобразованной молекулы РНК для правильной расстановки аминокислот в полипептидной цепи. Было высказано мнение, что предобразованная РНК, необходимая для изменения типа синтезируемого белка, должна обладать высокой скоростью обновления своего состава, т. е. молекула такой РНК должна синтезироваться и распадаться с такой скоростью, чтобы обеспечить быструю обновляемость нуклеотидного состава. Фактически же рРНК сказалась метаболически весьма стабильно, поэтому становилась очевидным, что она не может служить в качестве матрицы.
В ряде лабораторий были получены данные о существовании в клетках в соединении с рибосомами короткоживущей РНК, названной информационной РНК; сейчас она обозначается как матричная РНК, потому что ее роль заключается в переносе информации от ДНК в ядре до цитоплазмы, где она соединяется с рибосомами и служит матрицей, на которой происходит синтез белка.
Эти опыты открыли прямую дорогу для экспериментальной расшифровки кода, при помощи которого информация от РНК передается на синтезируемый белок. Последовательность нуклеотидов РНК реализуется в специфической последовательности аминокислот синтезируемой полипептидной цепи. Опыты Ниренберга свидетельствуют также о том, что не рибосома и не рРНК являются матрицей, на которой синтезируются специфические белки, а эту роль выполняют поступающие извне матричные РНК. Итак, ДНК предает информацию на РНК, которая синтезируется в ядре и затем поступает в цитоплазму. Здесь РНК выполняет матричную функцию для синтеза специфической белковой молекулы. Матричная гипотеза синтеза белка, как и других полимерных молекул ДНК и РНК, получила в настоящее время полное подтверждение. Ее правильность была доказана в экспериментах, которые обеспечивали точное воспроизведение первичной структуры полимерных молекул; причем этот синтез в отличии от беспорядочного химического синтеза отличался не только высокой скоростью и специфичностью, но и направленностью самого процесса, в строгом соответствии с программой, записанной в линейной последовательности молекулы матрицы.
Аминоацил –тРНК синтетазы
В цитозоле клеток 20 различных аминокислот присоединяются a-карбоксильной группой к 3¢-гидрофильному акцепторному концу соответствующих тРНК с образованием сложноэфирной связи. Эти реакции катализирует семейство ферментов, носящее название аминоацил –тРНК синтетаз. Каждый член этого семейства узнает только одну определенную аминокислоту и те тРНК, которые способны связываться с этой аминокислотой. Из этого следует, что в группу тРНК синтетаз входит 20 различных ферментов. Они осуществляют активацию аминокислот в 2 стадии: на первой стадии аминокислота присоединяется к ферменту и реагирует с АТФ с образованием богатого энергией промежуточного соединения –аминоацил АМФ. На второй стадии аминоацильный остаток аминоациладенилата, оставаясь связанным с ферментом, взаимодействует с молекулой соответствующей тРНК с образованием аминоацил тРНК.
Для каждой аминокислоты существует свой фермент — своя аминоацил тРНК синтетаза: для глутамата — глутамил-тРНК синтетаза, гистидина — гистидил-тРНК синтетаза и т.д.
Аминокислоты присоединяются к 3'- или 2'-ОН группам рибозы на З'-конце тРНК, где все тРНК имеют общую нуклеотидную последовательность -ССА.
Энергия, заключённая в макроэргической сложноэфирной связи аминоацил-тPHK, впоследствии используется на образование пептидной связи в ходе синтеза белка.
Пирофосфат, выделяющийся в ходе этой реакции, гидролитически расщепляется с образованием двух молекул ортофосфата и выделением энергии, что делает реакцию активации аминокислот необратимой.
Чрезвычайно высокая специфичность аа-тРНК синтетаз в связывании аминокислоты с соответствующими тРНК лежит в основе точности трансляции генетической информации. В активном центре этих ферментов есть 4 специфических участка для узнавания: аминокислоты, тРНК, АТФ и четвёртый — для присоединения молекулы Н20, которая участвует в гидролизе неправильных аминоациладенилатов. За счёт существования в активном центре этих ферментов корректирующего механизма, обеспечивающего немедленное удаление ошибочно присоединённого аминокислотного остатка, достигается поразительно высокая точность работы: на 1300 связанных с тРНК аминокислот встречается только одна ошибка.
Аминокислота, присоединяясь к тРНК, в дальнейшем не определяет специфических свойств аа-тРНК, так как её структуру не узнаёт ни рибосома, ни мРНК. Участие в синтезе белка зависит только от структуры тРНК, а точнее, от комплиментарного взаимодействия антикодона аминоацил-тРНК с кодоном мРНК.
Антикодон расположен в центральной (антикодоновой) петле тРНК. Узнавание тРНК аа-тРНК синтетазами не всегда происходит по антикодоновой петле. Активный центр некоторых ферментов обнаруживает комплиментарное соответствие другим участкам пространственной структуры тРНК.
Рибосомы
Рибосомы представляют собой рибонуклеопротеиновые образования — своеобразные «фабрики», на которых идёт сборка аминокислот в белки. Эукариотические рибосомы имеют константу седиментации 80S и состоят из 40S (малой) и 60S (большой) субъединиц. Каждая субъединица включает рРНК и белки. В 40S субъединицу входит рРНК с константой седиментации 18S и около 30—40 белков. В 60S субъединице обнаружено 3 вида рРНК: 5S, 5,8S и 28S и около 50 различных белков.
Белки входят в состав субъединиц рибосомы в количестве одной копии и выполняют структурную функцию, обеспечивая взаимодействие между мРНК и тРНК, связанными с аминокислотой или пептидом.
В присутствии мРНК 40S и 60S субъединиц объединяются с образованием полной рибосомы, масса которой примерно в 650 раз больше массы молекулы гемоглобина.
В рибосоме есть 2 центра для присоединений молекул тРНК: аминоацильный (А) и пептидильный (Р) центры, в образовании которым участвуют обе субъединицы. Вместе центры А и Р включают участок мРНК, равный 2 кодонам. В ходе трансляции центр А связываем аа-тРНК, строение которой определяет кодон, находящийся в области этого центра. В струкЯ туре этого кодона зашифрована природа аминокислоты, которая будет включена в растущую полипептидную цепь. Центр Р занимает пептидил-тРНК, т.е. тРНК, связанная с пептидной цепочкой, которая уже синтезирована.
У эукариотов различают рибосомы 2 типом «свободные», обнаруживаемые в цитоплазма клеток, и связанные с эндоплазматическим ретикулумом (ЭР). Рибосомы, ассоциированнье с ЭР, ответственны за синтез белков «на экспорт», которые выходят в плазму крови и участвуют в обновлении белков ЭР; мембраны aаппарата Гольджи, митохондрий или лизосом.
Митохондрии содержат свой набор рибосом. Митохондриальные рибосомы мельче, чем рибосомы эукариотов, прокариотов и имеют константу седиментации 55S. Они также состояв из двух субъединиц, но отличаются от эукаририотических рибосом количеством и составом РНК и белков.
Белковые факторы
В каждой стадии белкового синтеза на рибосоме: инициации, элонгации и терминации участвует разный набор внерибосомных белковый факторов. Эти белки связываются с рибосомой или её субъединицами на определённых стадиях процесса и стабилизируют или облегчают функционирование белоксинтезирующей машины.
АТФ и ГТФ как источники энергии
На включение одной аминокислоты в растущую полипептидную цепь клетка затрачивает 4 макроэргические связи: 2 из АТФ в ходе реакции, катализируемой аа-тРНК синтетазой (в процессе активации аминокислот АТФ расщепляется на АМФ и пирофосфат), и 2 молекулы ГТФ: одна используется на связывание аа-тРНК в А-центре рибосомы, а вторая затрачивается на стадию транслокации. К этому |следует добавить использование ещё двух макроэргических связей молекул: АТФ и ГТФ на инициацию и терминацию синтеза полипептидной цепи.
Этапы синтеза полипептидной цепи
Синтез белка представляет собой циклический многоступенчатый энергозависимый процесс, в котором свободные аминокислоты полимеризуется в генетически детерминированную последовательность с образованием полипептидов. Система белкового синтеза, точнее система трансляции, которая использует генетическую информацию, транскрибированную в мРНК, для синтеза полипептидной цепи с определенной первичной структурой, включает около 200 типов макромолекул – белков и нуклеиновых кислот. Среди них около 100 макромолекул, участвующих в активировании аминокислот и их переносе на рибосомы, более 60 макромолекул, входящих в состав 70S или 80S рибосом, и около 10S макромолекул, принимающих непосредственное участие в системе трансляции. Не разбирая природу других важных для синтеза факторов, рассмотрим подробно механизм индивидуальных путей синтеза белковой молекулы в искусственной синтезирующей системе. Прежде всего при помощи изотопного метода было выяснено, что синтез белка начинается с N-конца и завершается C-концом, т.е. процесс протекает в направлении : NH2®COOH.
Белковый синтез, или процесс трансляции, может быть условно разделен на 2 этапа: активирование аминокислот и собственно процесс трансляции.
Второй этап матричного синтеза белка, собственно трансляцию, протекающей в рибосоме, условно делят на три стадии: инициации, элонгации и терминации.
Активирование аминокислот
Необходимым условием синтеза белка, который в конечном счете сводится к полимеризации аминокислот, является наличием в системе не свободных, а так называемых активированных аминокислот, располагающих своим внутренним запасом энергии. Активация свободных аминоксилот осуществляется при помощи специфических ферментов аминоацил –тРНК-синтетаз в присутствии АТФ. Этот процесс протекает в 2 стадии:
Обе стадии катализируются одним и тем же ферментом. На первой стадии аминокислота реагирует с АТФ и образуется пирофосфат и промежуточный продукт, который на второй стадии реагирует с соответствующей 3¢-ОН-тРНК, в результате чего образуется аминоацил –тРНК и освобождается АМФ. Аминоацил-тРНК располагает необходимым запасом энергии.
Аминокислота присоединяется к концевому 3¢-ОН-гидроксилу АМФ, который вместе с двумя остатками ЦМФ образует концевой триплет ЦЦА, являющийся одинаковым для всех транспортных РНК.
Процессы трансляции
Инициация
Инициация трансляции представляет собой событие, в ходе которого происходит образованиe комплекса, включающего Мет-тРНКiмет, мРНК и рибосому, где—тРНКiмет инициирующая метиониновая тРНКВ этом процессе участвуют не менее 10 факторов инициации, которые обозначают как elF (от англ. eukaryotic initiation factors) с указанием номера и буквы. Первоначально 40S субъединица рибосомы соединяется с фактором инициации, который препятствует её связыванию с 60S субъединицей, но стимулирует объединение с тройным комплексом, включающим Мет-тРНКiмет, eIF-2 и ГТФ. Затем этот теперь уже более сложный комплекс связывается с 5'-концом мРНК при участии нескольких elF. Один из факторов инициации (elF-4F) узнаёт и присоединяется к участку «кэп» на молекуле мРНК, поэтому он получил название кэпсвязывающего белка. Прикрепившись к мРНК, 40S субъединица начинает скользить по некодирующей части мРНК до тех пор, пока не достигнет инициирующего кодона AUG кодирующей нуклеотидной последовательности. Скольжение 40S субъединицы по мРНК сопровождается гидролизом АТФ, энергия которого затрачивается на преодоление участков спирализации в нетранслируемой части мРНК. В эукариотических клеках некодирующие участки мРНК имеют разную длину, но обычно от 40 до 80 нуклеотидов, хотя встречаются области с протяжённостью более 700 нуклеотидов.
Достигнув начала кодирующей последовательности мРНК, 40S субъединица останавливается и связывается с другими факторами инициации, ускоряющими присоединение 60S субъединицы и образование 80S рибосомы за счёт гидролиза ГТФ до ГДФ и неорганического фосфата. При этом формируются А- и Р-центры рибосомы, причём в Р-центре оказывается AUG-кодон мРНК с присоединённым к нему Мет-тРНКiмет.
В клетках есть 2 различающиеся по структуре тРНК, узнающие кодон AUG. Инициирующий кодон узнаёт тРНКiмет, а триплеты мРНК, кодирующие включение метионина во внутренние участки белка, прочитываются другой тРНКiмет
Элонгация
По завершении инициации рибосома располагается на мРНК таким образом, что в Р-центре находится инициирующий кодон AUG с присоединённой к нему Мет-тРНКiмет., а в А-центре — триплет, кодирующий включение первой аминокислоты синтезируемого белка. Далее начинается самый продолжительный этап белкового синтеза — элонгация, в ходе которого рибосома с помощью аа-тРНК последовательно «читает» мРНК в виде триплетов нуклеотидов, следующих за инициирующим кодоном в направлении от 5' к З'-концу, наращивая полипептидную цепочку за счёт последовательного присоединения аминокислот.
Включение каждой аминокислоты в белок происходит в 3 стадии, в ходе которых:
• аа-тРНК каждой входящей в белок аминокислоты связывается с А-центром рибосомы;
• пептид от пептидил-тРНК, находящейся в Р-центре, присоединяется к a-NH2-группe аминоацильного остатка аа-тРНК А-центра с образованием новой пептидной связи;
• удлинённая на один аминокислотный остаток пептидил-тРНК перемещается из А-центра в Р-центр в результате транслокации рибосомы.
Связывание аминоацил-тРНК в А-центре. Кодон мРНК, располагающийся в А-центре рядом с инициирующим кодоном, определяем природу аа¢-тРНКаа¢, которая будет включена в А-центр. аа¢-тРНКаа¢ взаимодействует с рибосомой в виде тройного комплекса, состоящего из фактора элонгации EF-1, аа¢-тРНКаа¢, и ГТФ Комплекс эффективно взаимодействует с рибосомой лишь в том случае, если антикодон аа¢-тРНКаа¢, комплиментарен и антипараллелен кодону мРНК в А-центре. Включение аа¢-тРНКаа¢, в рибосому происходит за счёт энергии гидролиза ГТФ до ГДФ и неорганического фосфата
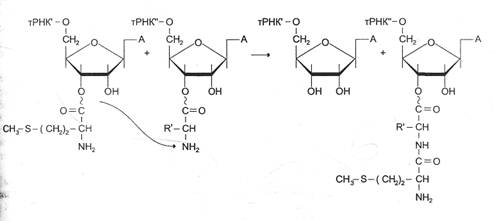
Образование пептидной связи происходит сразу же после отщепления комплекса EF-1 и ГДФ от рибосомы. Эта стадия процесса получила название реакции транспептидации.
В ходе этой реакции остаток метионина аа¢-тРНКаа¢, связывается с a-аминогруппой первой аминокислоты, присоединённой к тРНКаа¢, и pacположенной в А-центре, образуется первая пептидная связь. Установлено, что пептидилтранс|феразная активность большой субъединици рибосомы принадлежит 28S рРНК. К настоящему времени обнаружена целая группа РНК, обладающая свойствами ферментов. Эти каталитически активные РНК получили название рибозимов Полагают, что рибозимы можно считать «реликтами» раннего периода революции, когда белки ещё не приобрели такого значения, как в последующие периоды.
Транслокация — третья стадия элонгации. К рибосоме присоединяется фактор элонгации EF-2 и за счёт энергии ГТФ продвигает рибосому по мРНК на один кодон к З'-концу. В результате дипептидил-тРНК, которая не меняет своего положения относительно мРНК, из А-центра перемещается в Р-центр. Свободная от метионина тРНКаа¢, покидает рибосому, а в область А-центра попадает следующий кодон.
По завершении третьей стадии элонгации рибосома в Р-центре имеет дипептидил-тРНК, а в А-центр попадает триплет, кодирующий включение в полипептидную цепь второй аминокислоты. Начинается следующий цикл стадии элонгации, в ходе которого на рибосоме снова проходят вышеописанные события. Повторение таких циклов по числу смысловых кодонов мРНК завершает весь этап элонгации.
Терминация
Терминация трансляции наступает в том случае, когда в А-центр рибосомы попадает один из стоп-кодонов: UAG, UAA или UGA. Для стоп-кодонов нет соответствующих тРНК. Вместо этого к рибосоме присоединяются 2 белковых высвобождающих фактора RF (от англ, releasing/actor) или фактора терминации. Один из них с помощью пептидилтрансферазного центра катализирует гидролитическое отщепление синтезированного пептида от тРНК. Другой за счёт энергии гидролиза ГТФ вызывает диссоциацию рибосомы на субъединицы.
Интересно отметить, что факторы трансляции, реализующие эффекты за счёт гидролиза ГТФ, являются членами суперсемейства G-белков, в которое входят G-белки, участвующие в трансдукции сигналов гормонов и других биологически активных веществ, и Ras-белки, функционирующие как факторы роста. Все G-белки связывают и гидролизуют ГТФ. Когда они связаны с ГТФ, то активны и участвуют в соответствующих метаболических процессах, а когда в активном центре в результате гидролиза ГТФ превращается в ГДФ, эти белки приобретают неактивную конформацию.
Таким образом, матричная природа процесса трансляции проявляется в том, что последовательность поступления аминоацил-тРНК рибосому для синтеза белка строго детерминирована мРНК, т.е. порядок расположения кодонов вдоль цепи мРНК однозначно задает структуру синтезируемого белка. Рибосома сканирует цепь мРНК в виде триплетов и последовательно отбирает из окружающей среды «нужные» аа-тРНК, освобождая в ходе элонгаци деацилированные тРНК.
Малая и большая субъединицы рибосомы: процессе трансляции выполняют разные функции: малая субъединица присоединяет мРНК декодирует информацию с помощью тРНК механизма транслокации, а большая субъеданица ответственна за образование пептидных связей.
Полирибосомы
В процессе синтеза белка рибосома присоединяется к 5'-концу мРНК и перемещается в направлении З'-конца. При этом 5'-конец мРНК освобождается, и к нему может присоединиться новая рибосома, на которой начинается poст ещё одной полипептидной цепи. Как правило много рибосом одновременно участвует в синтезе белка на одной и той же мРНК, образуя комплекс, который называют полирибосомой, или полисомой.
Каждая рибосома занимает на мРНК участок длиной около 80 нуклеотидов, поэтому рибрсомы располагаются на мРНК с интервалом примерно в 100 нуклеотидов. Чем длиннее полипептидная цепочка синтезируемого белка, тем больше рибосом может одновременно осуществлять синтез этого белка, значительно увеличиваая таким образом эффективность использования матрицы.
Каждая рибосома способна катализировать образование около 100 пептидных связей в минуту. Полирибосомы могут существовать в виде частиц, плавающих в цитоплазме клеток, или могут быть связаны с ЭР. Свободные цитоплазматические полирибосомные частицы ответственны за синтез белков, выполняющих внутриклеточные функции. Полирибосомы, ассоциированные с ЭР, под электронным микроскопом имеют вид «шероховатой» поверхности. Белки, синтезируемые «шероховатым» ЭР, должны транспортироваться через мембрану для того, чтобы они достигли места окончательной локализации. Для них характерно присутствие на N-конце лидерной, или сигнальной, последовательности длиной от 15 до 30 аминокислотных остатков, которая содержит много аминокислот с гидрофобными радикалами и обеспечивает прохождение белка через липидный бислой мембран. Некоторые из этих белков для дальнейшего транспорта упаковываются аппаратом Гольджи в секреторные гранулы.
Транспорт синтезированных белков через мембраны.
Помимо использования белков для нужд самой клетки, многие так называемые экспортируемые белки, которые функционируют вне клетки, подвергнутся переносу через клеточную мембрану при помощи особых низкомолекулярных пептидов (от 15 до 30 аминокислот), получивших название лидирующих, или сигнальных, пептидов. Особенностью их состава является преимущественное содержание гидрофобных радикалов, что позволяет им легко проникать через бислойную липидную мембрану или встраиваться в мембрану. Эти сигнальные последовательности в рибосомах образуются первыми с N-конца при синтезе белка по программе сигнальных кодонов, расположенных сразу после инициаторного кодона, и легко узнаются рецепторными участками мембраны эндоплазматической сети. При этом образуется комплекс между мРНК, рибосомой и мембранными рецепторными белками, формируя своеобразный канал в мембране, через который сигнальный пептид проникает внутрь цистерны эндоплазматической сети, увлекая и протаскивая за собой синтезируемую и растущую молекулу секреторного белка. В процессе прохождения или после проникновения полипептида в цистерны N-концевая сигнальная последовательность отщепляется под действием особой лидирующей (сигнальной) пептидазы, а зрелый белок через пластинчатый комплекс (аппарат Гольджи) может покидать клетку в форме секреторного пузырька. Следует указать на возможность активного участия в транспорте белков и других полимерных молекул через мембраны, помимо сигнальных пептидов, также особых белков, получивших наименование поринов; химическая природа и механизм их действия выяснены пока недостаточно.
Синтез митохондриальных белков
В митохондриях клеток высших организмов содержится до 2% клеточной ДНК, отличающейся от ДНК ядра. Митохондрии содержат весь аппарат, включая рибосомы, тРНК и мРН К, необходимый для синтеза определенных белков. Синтезируемые в митохондриях белки в основном относятся к нерастворимым белкам, участвующим в организации структуры этих органелл, в то время как источником синтеза растворимых митохондриальных белков являются рибосомы цитоплазмы, откуда они затем транспортируются в митохондрии. Рибосомы в митохондриях имеют меньший размер чем 80S рибосомы в цитоплазме. Интересно отметить, что в качестве инициирующей аминокислоты при синтезе белка в митохондриях эукариот может участвовать N-формилметионин, а не свободный метионин, как в цитоплазме. Это обстоятельство свидетельствует о том, что митохондриальный синтез белка по своему механизму, очевидно, близок к синтезу белка у прокариот.
Посттрансляционные модификации полипептидной цепи
Полипептидные цепи могут подвергаться структурным модификациям, либо будучи ещё связанными с рибосомами, либо после завершения синтеза. Эти конформационные и структурные изменения полипептидных цепей получили название посттрансляционных изменений. Они включают удаление части полипептидной цепи, ковалентное присоединение одного или нескольких низкомолекулярных лигандов, приобретение белком нативной конформации.
Многие модификации осуществляются в ЭР. Здесь происходят фолдинг полипептидных цепей и формирование уникальной третичной или четвертичной структуры белков. Причём для поддержания нативной конформации молекул огромное значение имеет правильное формирование дисульфидных связей.
Частичный протеолиз
Многие белки, секретируемые из клеток, первоначально синтезируются в виде молекул-предшественников, функционально неактивных. Удаление части полипептидной цепи специфическими эндопротеазами приводит к образованию активных молекул. Некоторые белки-предшественники расщепляются в ЭР или аппарате ; Гольджи. другие — после секреции. Так, неактивные предшественники секретируемых ферментов — зимогены — образуют активный фермент после расщепления по определённым участкам молекулы: зимоген панкреатической железы трипсиноген превращается в активный трипсин после секреции в тонкий кишечник.
Наглядным примером последовательного двухстадийного протеолиза служит образование активных форм пептидных гормонов (например, инсулина или глюкагона) из препрогормонов. Первоначально N-концевой сигнальный пептид молекулы-предшественника удаляется в ЭР процессе синтеза белка и образуется неактивный прогормон. Затем прогормон в секреторных гранулах, формирующихся в аппарате Гольджи подвергается действию эндо- и/или экзопротеаз и превращается в активный гормон.
Ковалентные модификации
Структурные белки и ферменты могут акгивироваться или инактивироваться в результате присоединения различных химических групп фосфатных, ацильных. метальных, олигосахаридных и некоторых других.
Фосфорилирование белков осуществляется по гидроксильным группам серина, треонина и, реже, тирозина ферментами из группы протеинкиназ, тогда как дефосфорилирование катализируют гидролитические ферменты фосфопротеинфосфатазы
Гликозилирование. Белки, входящие в состав плазматических мембран или секретирующиеся из клеток, подвергаются гликозидированию. Углеводные цепи присоединяются по гидроксильным группам серина или треонина (О-гликозилирование) либо аспарагина (N-гликозилирование). Последовательное наращивание углеводного фрагмента происходит в ЭР и аппарате Гольджи.
Многочисленным модификациям подвергаются боковые радикалы некоторых аминокислот: в тиреоглобулине йодируются остатки тирозина; в факторах свёртывания крови карбоксилируются остатки глутамата; в ЭР фибробластов гидроксилируился остатки пролина и лизина в цепях тропоколлагена.
Регуляция синтеза белка
Основным условием существования любых живых организмов является наличие тонкой, гибкой, согласованно действующей системы регулирования, в которой все элементы тесно связаны друг с другом. В белковом синтезе не только количественный и качественный состав белков, но и время синтеза имеет прямое отношение ко многим проявлениям жизни. В частности, от этого зависит приспособление микроорганизмов к условиям окружающей питательной среды как биологической необходимости или приспособление сложного многоклеточного организма к физиологическим потребностям при изменении внутренних и внешних условий.
Клетки живых организмов обладают способностью синтезировать огромное количество разнообразных белков. Однако они никогда не синтезируют все белки. Количество и разнообразие белков, в частности ферментов, определяются степенью их участия в метаболизме. Более того, интенсивность обмена регулируется скоростью синтеза белка и параллельно контролируется аллостерическим путем. Таким образом, синтез белка регулируется внешними и внутренними условиями, которые диктуют клетке синтез такого количества белка и таких белков, которые необходимы для выполнения физиологических функций. Все это свидетельствует о весьма сложном, тонком и целесообразном механизме регуляции синтеза белка в клетке.
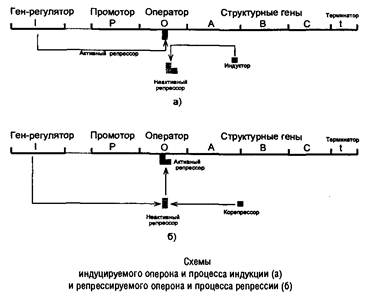
Общую теорию регуляции синтеза белка разработали Ф. Жакоб и Ж. Моно. Сущность этой теории сводится к «выключению» или «включению» генов как функционирующих единиц, к возможности или невозможности проявления их способности передавать закодированную в структурных генах ДНК генетическую информацию для синтеза специфических белков. Эта теория, доказанная в опытах на бактериях, получила широкое признание, хотя в эукариотических клетках механизм регуляции синтеза белка вероятно более сложный. У бактерий доказана индукция ферментов (т. е. синтез ферментов de novo) при добавлении в питательную среду субстратов этих ферментов. Добавление конечных продуктов реакции, образование которых катализируется этими же ферментами, напротив, вызывает уменьшение количества синтезируемых ферментов. Это последнее явление получило название репрессии синтеза ферментов. Оба явления — индукция и репрессия — взаимосвязаны.
Согласно теории Жакоба и Моно в биосинтезе белка у бактерий участвуют по крайней мере три типа генов: структурные гены, ген-регулятор и ген-оператор. Структурные гены определяют первичную структуру синтезируемого белка. Именно эти гены в цепи ДНК являются основой для биосинтеза мРНК, которая затем поступает в рибосому и, как было указано выше, служит матрицей для биосинтеза белка.
Синтез мРНК на структурных генах молекулы ДНК непосредственно контролируется определенным участком, называемым геном-оператором. Он служит как бы пусковым механизмом для функционирования структурных генов. Ген-оператор локализован на крайнем отрезке структурного гена или структурных генов, регулируемых им. «Считывание» генетического кода, т. е. формирование мРНК, начинается спромотора— участка ДНК, являющегося точкой инициации для синтеза мРНК, и далее распространяется последовательно вдоль оператора и структурных генов. Координированный одним оператором одиночный ген или группа структурных генов образует оперон.
В свою очередь деятельность оперона находится под контролирующим влиянием другого участка цепи ДНК, получившего название гена-регулятора. Поскольку структурные гены и ген-регулятор находятся в разных участках цепи ДНК, связь между ними, как предполагают Ф. Жакоб и Ж. Моно, осуществляется при помощи вещества-посредника, оказавшегося белком и названного репрессором. Образование репрессора происходит в рибосомах ядра на матрице специфической мРНК, синтезированной на гене-регуляторе. Репрессор имеет сродство к гену-оператору и обратимо соединяется с ним в комплекс. Образование такого комплекса приводит к блокированию синтеза мРНК и, следовательно, синтеза белка, т.е. функция гена-регулятора состоит, в том, чтобы через белок-репрессор прекращать деятельность структурных генов, синтезирующих мРНК. Репрессор, кроме того, обладает способностью строго специфически связываться с определенными низкомолекулярными веществами, называемыми индукторами, или эффекторами. Когда такой индуктор соединяется с репрессором, последний теряет способность связываться с геном-оператором, который таким образом выходит из-под контроля гена-регулятора, и начинается синтез мРНК.
Это типичный пример отрицательной формы контроля, когда индуктор, соединяясь с белком-репрессором, вызывает изменения его третичной структуры настолько, что репрессор теряет способность связываться с геном-оператором. Этот процесс аналогичен взаимоотношениям аллостерического центра фермента с эффектором, под влиянием которого изменяется третичная структура фермента и он теряет способность связываться со своим субстратом.
Механизм описанной регуляции синтеза белка и взаимоотношения репрессора со структурными генами были доказаны в опытах на Е. coli, на примере синтеза Р-галактозидазы (лактазы) — фермента, гидролизующего молочый сахар на глюкозу и галактозу. Дикий штамм Е. coli, обычно растущий на глюкозе, не может расти, если вместо глюкозы в питательную среду добавить лактозу (новый источник энергии и углерода) до тех пор, пока не будут синтезированы соответствующие ферменты (адаптивный синтез). При поступлении в клетку лактозы (индуктора) молекулы ее связываются с белком-репрессором и блокируют связь между репрессором и геном-оператором. При этом ген-оператор и структурные гены начинают снова функционировать и синтезировать необходимую мРНК, которая «дает команду» рибосомам синтезировать р-галактозидазу. Одновременно ген-регулятор продолжает вырабатывать репрессор, но он блокируется новыми молекулами лактозы, поэтому синтез фермента продолжается. Как только молекулы лактозы будут полностью расщеплены, репрессор освобождается и, поступив в ДНК, связывает ген-оператор и блокирует синтез мРНК, а следовательно, синтез Р-галактозидазы в рибосомах.
Таким образом, биосинтез мРНК, контролирующий синтез белка в рибосомах, зависит от функционального состояния репрессора. Если репрессор, который представляет собой белок, построенный из 4 субъединиц с общей молекулярной массой около 150000 Да, находится в активном состоянии, не связан с индуктором, то он блокирует ген-оператор и синтез мРНК не происходит. При поступлении метаболита-индуктора в клетку его молекулы связывают репрессор, превращая его в неактивную форму (или, возможно, снижая его сродство к гену-оператору). Структурные гены выходят из-под запрещающего контроля и начинают синтезировать нужную мРНК.
Выше было указано, что концентрация ряда ферментов в клетках резко снижается при увеличении концентрации отдаленных конечных продуктов, образующихся в цепи последовательных ферментативных реакций. Такой эффект, получивший название репрессии ферментов, часто наблюдается при реакциях биосинтеза. В этих случаях оказалось, что молекулы репрессора, также образующиеся в рибосомах ядра по «команде» гена-регулятора, являются неактивными и сами по себе не обладают способностью подавлять деятельность гена-оператора и, следовательно, всего оперона, но приобретают такую способность после образования комплекса с конечным или одним из конечных продуктов биосинтетического процесса.
Конечный продукт выступает, таким образом, в качестве корепрессора. Имеются данные, показывающие, что в качестве корепрессоров в синтезе ферментов обмена аминокислот выступает не свободная аминокислота как конечный продукт биосинтетической реакции, а комплекс ее с тРНК — аа-тРНК.
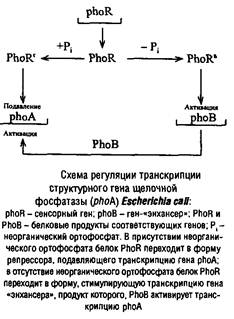
В регуляции экспрессии структурных генов специфическое участие принимает особый белок, получивший название катаболитный ген-активирующий белок (от англ, catabolite gene activation protein, сокращенно обозначаемый САР); этот белок взаимодействует с цАМФ, образуя комплекс, способствующий прикреплению РНК-полимеразы к промоторному участку генома. В присутствии комплекса САР-цАМФ фермент может начать транскрипцию оперона, включая структурные гены, т. е. в клетках имеется еще один, дополнительный САР-цАМФ регулятор, действующий скорее всего в качестве положительного регулятора, поскольку его присутствие необходимо для начала экспрессии гена. Таким образом, концепции Жакоба и Моно о механизме проявления активности генов признана одним из блестящих достижений молекулярной биологии. Она явилась логическим развитием многочисленных исследований, проведенных генетиками и биохимиками в предшествующие десятилетия.
В заключение следует вкратце рассмотреть вопрос о регуляции процессов дифференцировки клеток высших организмов. ДНК, присутствующая во всех соматических клетках, вероятнее всего, имеет одинаковую первичную структуру у данного организма и соответственно располагает информацией для синтеза любых или всех белков тела. Тем не менее клетки печени, например, синтезируют сывороточные белки, а клетки молочной железы — белки молока. Нет сомнения в том, что в дифференцированных клетках, очевидно, существует тонкий механизм контроля деятельности ДНК в разных тканях, обеспечивающий синтез многообразия белков.
Механизмы, лежащие в основе этой регуляции, пока неизвестны. Для объяснения их имеется ряд гипотез. Предполагается, что контроль осуществляется на уровне транскрипции по аналогии с индукцией ферментов у бактерий и что в этом случае в клетках животных должны функционировать аналогичные репрессоры.. Поскольку с молекулой ДНК у зукариот связаны гистоны, считается, что именно они выполняют роль репрессоров. Однако прямые доказательства их роли в качестве репрессоров отсутствуют, как и точные данные о существовании и природе каких-либо репрессоров в клетках эукариот. Высказано предположение, что в ядре синтезируется гигантская молекула мРНК, содержащая информацию для синтеза широкого разнообразия белков, но в цитоплазму, как было показано выше, попадает только небольшая часть зрелой мРНК, а основная часть распадается. Неясны, однако, биологический смысл и назначение этого механизма избирательного распада и, соответственно, траты огромной части молекулы мРНК.
Существует еще одно предположение, что на ДНК клетки синтезируются все возможные мРНК, которые поступают в цитоплазму, и процесс трансляции регулируется путем специфического и избирательного взаимодействия с определенными молекулами мРНК.
Ингибиторы синтеза белка
Одним из путей выяснения механизмов синтеза нуклеиновых кислот и белков в клетках является использование таких лекарственных препаратов, которые могли бы избирательно тормозить эти процессы у бактерий, не оказывая влияния на организм человека. Некоторые препараты действительно обладают таким действием, однако многие из них оказываются токсичными и для человека. В настоящее время в медицинской практике применяются многие антибиотики, часть из которых будет рассмотрена ниже с целью выяснения механизма их действия на ключевые химические реакции синтеза белка и нуклеиновых кислот.
Одним из мощных ингибиторов белкового синтеза является пуромицин. В результате структурного сходства с концевым остатком АМФ в аминоацил-тРНК' он легко взаимодействует с А-участком пептидил-тРНК с образованием пептидил-пуро-мицина.
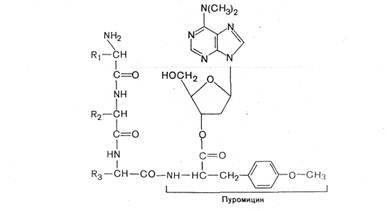
Поскольку пептидил-пуромицин не несет на себе триплета антикодона, он тем самым тормозит элонгацию пептидной цепи, вызывая обрыв реакции. При помощи пуромицина было доказано, например, что гормональный эффект в ряде случаев зависит от синтеза белка de novo. Укажем также, что пуромицин тормозит синтез белка как у прокариот, так и у эукариот.
Белковый синтез тормозится актиномицином D, обладающим противоопухолевым эффектом, который вследствие высокой токсичности применяется редко. Он оказывает тормозящее влияние на синтез всех типов клеточной РНК, в особенности мРНК. Это свойство вызвано тормозящим влиянием актиномицина D на ДНК-зависимую РНК-полимеразу, поскольку он связывается с остатками дезоксигуанозина цепи ДНК, выключая матричную функцию последней. Можно считать, что актиномицин D ингибирует транскрипцию ДНК.
Другим антибиотиком, также тормозящим синтез клеточной РНК, является используемый при лечении туберкулеза рифамицин. Этот препарат тормозит ДНК-зависимую РНК-полимеразу путем связывания с ферментом. Наиболее чувствительна к нему бактериальная РНК-полимераза. На организм животных этот антибиотик оказывает незначительное влияние. По механизму действия он резко отличается от актиномицина t). Следует указать на недавно открытое противовирусное действие рифамицина, в частности, он успешно используется при лечении трахомы, которая вызывается ДНК-содержащим вирусом. По-видимому, этот антибиотик найдет применение в лечении опухолей, вызываемых вирусами.
Выяснены механизмы действия ряда других антибиотиков, применяемых при лечении тифозных инфекций. Так, хлорамфеникол оказывает ингибирующее влияние на пептидилтрансферазную реакцию (на стадии элонгации) синтеза белка в 70S рибосоме бактерий. На этот процесс в 80S рибосоме он не действует. Противоположное тормозящее действие на синтез белка в 80S (без поражения процесса в 70S рибосоме) оказывает циклогексимид, являющийся ингибитором транслоказы.
Весьма интересен молекулярный механизм действия дифтерийного токсина. Он оказался наделен способностью катализировать реакцию АДФ-рибозилирования фактора элонгации (трансляционный фактор-2, TF-2). выключая тем самым его из участия в синтезе белка. Резистентность многих животных к дифтерийному токсину обусловлена трудностью проникновения токсина через мембрану клеток.
Противотуберкулезные и антибактериальные антибиотики, в частности стрептомицин и неомицин, действуют на белоксинтезирующий аппарат чувствительных к ним -штаммов бактерий. Высказано предположение, что эти антибиотики вызывают ошибки в трансляции мРНК, приводящие к нарушению соответствия между кодонами и включаемыми аминокислотами; например, кодон УУУ вместо фенилаланина начинает кодировать лейцин — в результате образуется аномальный белок, что приводит к гибели бактерий.
Широко применяемые в клинике тетрациклины также оказались ингибиторами синтеза белка в 70S рибосоме (меньше тормозится синтез в 80S рибосоме). Они легко проникают через клеточную мембрану. Считается, что тетрациклины тормозят связывание аминоацил-тРНК с аминоацильным центром в 50S субчастице рибосомы. Возможно, что тетрациклины химически связываются с этим центром, выключая тем самым одну из ведущих стадий процесса трансляции.
Пенициллины не являются истинными ингибиторами синтеза белка, однако их антибактериальный эффект связан с торможением синтеза гексапептидов, входящих в состав клеточной стенки. Механизм их синтеза отличается от рибосомального механизма синтеза белка. Эритромицин и олеандомицин тормозят активность транслоказы в процессе трансляции, подобно циклогексимиду, исключительно в 80S рибосомах, т. е. тормозят синтез белка в клетках животных.
Полученные к настоящему времени данные по механизму действия антибиотиков на синтез белка с учетом стадии и топографии процесса трансляции суммированы в табл. 13.2 (по Харперу).
Следует еще раз подчеркнуть, что нарушение или выпадение любого звена, участвующего в синтезе белка, почти всегда приводит к развитию патологии, причем клинические проявления болезни будут определяться природой и функцией белка, синтез которого оказывается нарушенным (структурный или функциональный белок). Иногда синтезируются так называемые аномальные белки как результат действия мутагенных факторов и, соответственно, изменения генетического кода (например, гемоглобин при серповидно-клеточной анемии). Последствия этих нарушений могут выражаться в развитии самых разнообразных синдромов или заканчиваться летально. Следует отметить, что организм располагает мощными механизмами защиты: подобные изменения генетического аппарата быстро распознаются специфическими ферментами — рестриктазами, измененные последовательности вырезаются и вновь замещаются соответствующими нуклеотидами при участии полимераз и лигаз.
Список литературы
Березов Т. Т., Коровкин Б. Ф. Биологическая химия//Учебная литература для студентов медицинского института, 1990.
А. С. Спирин Вестник Российской академии наук, том 71,2001.
Кнорре Д.Г., Мызина С.Д Биологическая химия: Учебник/.-3-е, испр. изд.-М.: Высш.шк., 2000.
В.И.Агол; Ред. А.С.Спирин-М Молекулярная биология: Структура и биосинтез нуклеиновых кислот: Учеб. для биол. спец. вузов/ Высш.шк., 1990.
Похожие работы
... от иРНК, а с другой – обеспечивает образование небольшого количества более длинных полипептидов, могущих выполнять важную функциональную роль. Регуляция биосинтеза белков путем посттрансляционной модификации Посттрансляционная модификация белков менее распространена, чем процессинг РНК. Тем не менее известны случаи, когда при развитии некоторых вирусов трансляция полицистронной матрицы ...

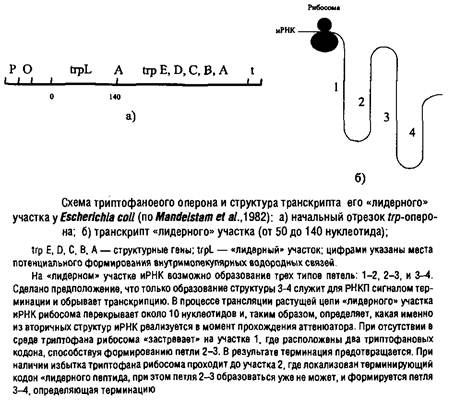
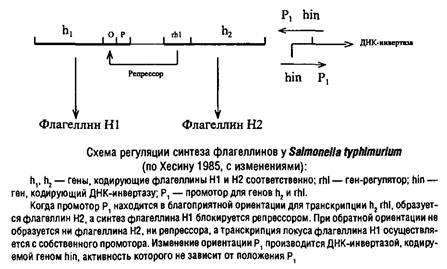
... процессы репликации ДНК и деления клетки. Реальные механизмы регуляции репликации еще не расшифрованы. У эукариот определенную роль в регуляции репликации, по-видимому, играют гистоны. 4. Транскрипция генетической информации Этот этап биосинтеза белков состоит в «переписывании» информации, закодированной в ДНК, на олигонуклеотидную последовательность информационной РНК и осуществляется ...
... . ЛИТЕРАТУРА: 1. Курс низших растений //Под ред. М. В. Горленко – М.: Высш. школа, 1981. 2. Жизнь растений, т. 1–3, - М.: Просвещение, 1974-77. 3. Н. Грин, У. Стаут, Д. Тейлор. Биология, т. 1-3. – М.: Мир, 1990. 4. Н.П.Горбунова , Е.С.Клюшникова, Н.А.Комарницкий. и др. Малый практикум по низшим растениям. – М.: Высш. школа, 1976. 5. Русанов В. А. Низшие растения. Методические указания ...
... которая обеспечивается печенью. Таким образом здоровый организм находится в равновесии с окружающей средой. Транспортные системы в организме человека. Метаболические процессы, протекающие во всех клетках тела, требуют непрерывного притока питательных веществ и кислорода и непрерывного удаления продуктов обмена. У некоторых видов животных транспортная система, кроме того, служит для переноса ...
0 комментариев