Содержание
Введение
Половой диморфизм
Брачное поведение и образование пар
Территориальное поведение
Гнездостроение и гнезда птиц
Яйцо и его особенности у птиц
Вскармливание потомства
Список использованной литературы
Введение
Птицы - высокоорганизованные позвоночные. Их тело покрыто перьями, передние конечности видоизменены в крылья. Имеют постоянную температуру тела, которая обеспечивается интенсивным обменом веществ. Птицы обладают сложным инстинктивным поведением. Известно около 9000 видов птиц.
Птицы отлично приспособлены к полету: тело обтекаемой формы, облегченный скелет, воздушные мешки в легких и др. На голове птиц есть пара глаз, позади которых расположены органы слуха. Глаза защищены дополнительным третьим веком. Птицы обладают очень острым зрением. Ухо состоит из трех отделов: наружного, среднего и внутреннего. Челюсти вытянуты в клюв и одеты роговым покровом. Форма и размеры клюва зависят от характера употребляемой пищи. В клюве различают надклювье (верхняя часть) и подклювье (нижняя часть).
Перьевой покров птиц составляют контурные и пуховые перья. Контурные перья имеют прочный стержень, нижняя часть которого (очин) погружена в кожу. Широкая часть пера — опахало состоит из бородок 1-го и 2-го порядка, сцепленных между собой крючочками. Среди контурных перьев различают рулевые, участвующие в управлении полетом, и маховые, образующие поверхность крыльев. Под контурными перьями расположены пуховые, имеющие тонкий стержень и бородки первого порядка. Они способствуют сохранению постоянной температуры тела.
Кожа птиц сухая, лишена желез. У основания рулевых перьев расположена одна копчиковая железа, выделяющая маслянистую жидкость. Птица смазывает ею перья, предохраняя их от намокания.
Кости скелета тонкие и прочные. Полости трубчатых костей заполнены воздухом, что облегчает полет птиц. Шейный отдел позвоночника образован большим числом подвижных позвонков. Поясничные и крестцовые позвонки прочно срослись и служат опорой туловищу. К грудным позвонкам прикрепляются ребра, образуя с грудиной грудную клетку. Грудина имеет вырост - киль, к которому крепятся грудные мышцы, приводящие в движение крылья. Череп имеет достаточно крупную мозговую коробку с большими глазницами и вытянутыми костями челюстей, лишенными зубов.
Легкие птиц отличаются сложным губчатым строением. Входящие в легкие бронхи многократно ветвятся и заканчиваются в полостях, а некоторые, выйдя из легких, расширяются, образуя воздушные мешки. Благодаря воздушным мешкам у птиц осуществляется двойное дыхание, и внутренние органы охлаждаются во время полета. Дыхание осуществляется расширением и сужением грудной клетки при помощи мышц, приближающих грудину к позвоночнику. В полете дыхание осуществляется благодаря взмахам крыльев, при этом воздушные мешки растягиваются, и в них поступает воздух из легких. При опускании крыльев мешки сжимаются, и воздух из них снова через легкие выходит наружу. Таким образом, кислород дважды поглощается в легких - при вдохе и при выдохе.
У птиц четырехкамерное сердце, состоит из двух предсердий и двух желудочков. Левая половина сердца содержит артериальную кровь, а правая - венозную. Движение крови происходит по двум кругам кровообращения - малому и большому, нигде не смешиваясь.
Органы выделения — парные почки. От них отходят мочеточники, по которым моча поступает в клоаку. Мочевой пузырь отсутствует, что облегчает вес птиц в полете.
Птицы раздельнополы. У самцов парные семенники, а у самок - один яичник. От половых желез в клоаку тянутся семяпроводы или яйцевод. Яйца созревают и приобретают оболочки, постепенно продвигаясь по яйцеводу самки. Яйцо содержит желток с большим запасом питательных веществ, окруженный жидким белком и известковой скорлупой. Скорлупа имеет поры, через которые свободно проходит воздух. Снаружи скорлупа покрыта тоненькой пленочкой, защищающей от проникновения в яйцо микробов.
На ранних стадиях развития зародыш проявляет сходство с рептилиями и другими низшими позвоночными, что говорит об их родстве. На более поздних стадиях развития зародыш приобретает птичьи черты. В зависимости от уровня развития различают выводковых и гнездовых птенцов. Первые вылупляются опушенными, зрячими, способными самостоятельно питаться и следовать за матерью. Вторые появляются на свет слепыми, голыми, неспособными питаться самостоятельно.[1]
В отличие от рептилий, птицы для выведения птенцов насиживают яйца. Яйца насиживает один из родителей или оба попеременно. Все птицы заботятся о потомстве: находят для него корм, согревают, защищают от хищников и обучают летать птенцов.
Репродуктивный комплекс поведения включает в себя все то, что связано с размножением животных, и потому он имеет огромное значение для популяции вида, обеспечивает его существование во времени, связь поколений, микроэволюцию и соответственно гомеостаз популяции. В этот комплекс включено большое количество разных экологических актов и программ.
Перед тем, как приступить к изучению и описанию, нужно отметить, что в репродуктивном комплексе преобладают внутривидовые взаимоотношения. Наиболее эволюционно стабильные. Соответственно здесь преобладающими, но не единственными, есть видовые стереотипы, которые базируются, в основном, на врожденной основе.
В репродуктивном комплексе поведения каждого вида можно отметить несколько периодов, каждые из них имеет свою программу и следует один за один в виде цепочки, который подчиняется собственным сигналам как экзогенного, так и эндогенного характера.
Половой диморфизм
Внешние различия самцов и самок в той или иной форме выражены почти у всех птиц. Эти различия могут проявляться в размерах тела, в окраске и иных особенностях наряда. Наиболее отчетлив половой диморфизм у куриных, гусеобразных и воробьиных, у которых самцы заметно крупнее самок, более ярко окрашены, иногда имеют более громкий голос и сложную песню. У других видов птиц он выражен только в размерах: самцы несколько крупнее самок. Однако в тех немногих случаях, когда заботу о потомстве несут только самцы, самки крупнее самцов (у трехперсток, куликов-плавунчиков, некоторых видов кукушек). Правда, у дневных хищников и сов основная забота о потомстве лежит на самках и тем не менее они крупнее самцов. Связано это с тем, что в первое время жизни птенцов самка главным образом находится у гнезда, а мелкий корм птенцам в это время приносит самец.
Половые различия в окраске характерны в отдельных случаях не только для оперения, но и для других частей тела — клюва, радужины, голых частей кожи, языка. Иногда половые различия выражены в наличии у одного пола (обычно у самцов) выростов и придатков кожи на голове (у куриных), в присутствии шпор у самцов (у фазановых), в развитии характерных перьев — хохлов, длинных рулевых перьев (у фазанов и др.), в устройстве голосового аппарата (в основном у утиных и певчих воробьиных), в пропорциях и размерах отдельных частей тела (клюв часто крупнее у самца, таз относительно шире у самок) и т. д.
Половой диморфизм проявляется обычно у птиц, достигших половой зрелости, и лишь изредка — в первом весеннем наряде. Особенно ярко он выражен в брачный период.[2]
Брачный наряд — частный случай полового диморфизма. Характеризуется он, как правило, яркими красками и свойствен многим птицам. Проявляется обычно к началу размножения (в брачный период). У одних видов этот наряд возникает после частичной линьки в виде ярко окрашенного оперения (у уток), у вторых — бледные перья на некоторых частях тела к весне стираются и из-под них выступают яркие участки оперения (у вьюрков, чеканов, каменок), у третьих к весне появляются ярко окрашенные выросты: красные брови (у куриных), выросты у основания клюва (у поганок) и т. д. У многих видов, имеющих две линьки в году, половой диморфизм проявляется только в брачном наряде.
Половой диморфизм (в том числе и брачный наряд) играют важную роль в нормальном течении полового цикла. Прежде всего, внешние различия между полами облегчают опознание и нахождение брачного партнера. Эти различия оказывают влияние также и на поведение, а вместе с тем и на деятельность половых желез партнера. Эксперименты с полевым воробьем показали, что удлинение светового дня дает лишь небольшую стимуляцию яичников. Если самка сидит в гнезде без присутствия рядом самца, то эта стимуляция скоро совсем исчезает. Следовательно, половой цикл птиц начинается под влиянием удлинения светового дня, но чтобы овогенез закончился яйцекладкой, необходимо воздействие на самку новых внешних раздражителей — наличие самца и гнезда. Половой диморфизм играет важную роль в половом отборе и особенно в той его форме, которая выражается в активном выборе самцов самками.
Брачное поведение и образование пар
С наступлением весны в жизни птиц появляются большие изменения. Дальние мигранты покидают зимовки и возвращаются на родину; кочующие приближаются к местам гнездования. В этот период происходит рассеивание зимних стай, занятие гнездовых территорий, разбивка на пары и закрепление за каждой парой гнездового участка, на котором будет располагаться гнездо и на котором большинство гнездящихся птиц будет вести оседлый образ жизни в течение всего периода размножения.
Здесь, скорее всего, все начинается из той сигнализации, с помощью которой особи разного пола, но одного вида, обнаруживают, находят и узнают о физиологической готовности к размножению особей другого пола. Эта сигнализация имеет очень большое значение для развития и существования вида, популяции. Она базируется на наследственно фиксированных особенностях видоспецифического поведения, которые изучает этология. Значение этой сигнализации весьма многогранно. Прежде всего, она дает возможность определять видовую принадлежность партнера, в значительной степени исключает межвидовую гибридизацию и сохраняет, таким образом, вид как морфофункциональную стабильность. Иными словами, «если два сходных по своей морфологии вида будут иметь совпадающий ареал, то их скрещиванию и появлению бесплодных межвидовых гибридов обычно будут препятствовать «этологические преграды». Это своего рода коды, которые передаются типичными для вида звуками, запахами, окраской, телодвижениями и мимикой. При этом, хотя у животных, принадлежащих к самым разным классам можно видеть принципиально много сходного, но у каждого вида есть свой характерный набор видовых брачных сигналов, которые могут также несколько различаться и в различных участках видового ареала. Брачные сигналы дают возможность встреч и контактов, созревших и созревающих особей разного пола, выбора ими наиболее подходящих партнеров (половой отбор) и даже нередко ведут к половому дозреванию тех особей, которые в данный момент еще недостаточно подготовлены к размножению».[3]
Перечисленные три канала брачной сигнализации часто применяются животными комплексно и, хотя все они были отнесены к дистантной рецепции дальнего действия, каждый из них имеет свои особенности и преимущества: звуковой канал — дистантный и хорошо применяется в условиях плохой видимости (заросли леса, водоросли водоема, темнота ночи), химический также может быть дистантным и, кроме того, следовой, т. е. надолго остающийся после ухода животного и, наконец, оптический позволяют с большой степенью точности определить местонахождение партнера, его внешние особенности и характер движений и мимики. На низких уровнях эволюции, пожалуй, преобладает химическая сигнализация, как это показано для коловраток, выделяющих в воду химические специфичные для вида вещества.
На более высоких ступенях эволюции, в ветви как первичноротых, так и вторичноротых к химической сигнализации подстраивается, все больше оптическая и акустическая сигнализация. Таким образом, как у млекопитающих (высшее звено ветви вторичноротых), так и у насекомых (высшее звено первичноротых) можно наблюдать обычно брачную сигнализацию, идущую по всем трем каналам. При этом у разных видов могут преобладать то один, то другие каналы. Так, например, самцы бабочки сатурнии (Saturnia pyri Schiff.) по запаху находили зрелых самок, прилетая к ним и ночной время на расстоянии 2 и более километров.
Пожалуй, больше всего внимания исследователи уделили брачным ритуалам у птиц. Дело в том, что в связи с уменьшенной возможностью у представителей этого класса к использованию хеморецепции у них особенно сильно развиты звуковая и зрительная сигнализации. Поэтому в данном случае исследователи располагают наилучшими возможностями регистрации сигналов, как с помощью зарисовок и фотокиноаппаратуры, так и средствами звукозаписи. В литературе описано великое множество брачных сигналов и ритуалов у птиц, особенно у моногамов. Брачное ритуальное поведение у них имеет еще существенное значение в том отношении, что при этом будущие родители как бы демонстрируют друг другу свои способности к заботе о будущем потомстве. Как пример он приводит подражание самкой большой синицы и некоторых других птиц позе, движениям и голосу просящего пищу птенца, что вызывает реакцию самца на поиск насекомых и акт кормления. Поскольку в заботе о потомстве у птиц заключается важнейшая часть репродуктивного поведения, то именно по этой линии идет половой отбор при первоначальной встрече пар.
Песни воробьиных птиц имеют значение не только брачных церемоний ухаживания. Они обозначают также занятость данной территории самцом и его гнездом. При нарушении этой территории другими самцами они немедленно изгоняются. Вообще в поведении многих птиц в репродуктивный период у самцов обычно значительным элементом поведения становится агрессия, поскольку они должны охранять от других особей данного вида свое гнездо и прилежащий к нему кормовой участок, который необходим для обеспечения пищей будущего потомства. Это развито особенно у неколониальпых моногамов. В этом случае агрессия в период размножения имеет в первую очередь значение для занятия популяций вида более широкой площади и для обеспечения кормом развивающейся молодой генерации.[4]
Образование пар у молодых гусей и лебедей, у молодых и старых крякв происходит уже во время осеннего пролета и на зимовках; у большинства других уток — на зимовках и весеннем пролете. Поэтому на места гнездования они прилетают парами. У большинства же птиц формирование пар происходит на местах гнездования. У многих голенастых, воробьиных и других птиц самцы прилетают раньше самок, занимают гнездовые участки, охраняют их и издают брачные крики, совершают своеобразные полеты. Прилетающих позднее самок привлекает брачное поведение самцов, и они оседают на приглянувшемся им участке.
Поведение птиц в брачный период резко меняется. В это время широко распространяются «брачные игры» и токование. Самцы принимают особые, издали бросающиеся в глаза позы, издают своеобразные крики. У некоторых птиц наблюдается групповое токование.
Групповое токование характерно для некоторых полигамных видов — глухарей, тетеревов, турухтанов и др. В определенном, специфическом для каждого вида месте обычно собирается группа самцов (от нескольких особей до нескольких десятков). Каждый самец совершает своеобразные телодвижения, принимает различные позы и вступает в драку с другими самцами. Некоторые при этом издают особые громкие, издалека слышимые звуки (глухарь, тетерев и др.), привлекающие самок, которые держатся на окраине токовища. Турнирные стычки и драки, на вид иногда ожесточенные, никогда не сопровождаются телесными повреждениями, они носят скорее демонстративный характер. Групповой ток делает самцов более заметными и поэтому облегчает встречу с самками; спаривание обычно происходит на периферии токовища. Одиночное токование свойственно моногамным видам. Формы токового поведения у этих птиц весьма разнообразны.
Воздушные игры и брачные полеты характерны для ряда видов. У дневных хищных птиц брачные полеты выражаются в том, что самец и самка взмывают высоко в воздух, падают вниз, преследуют друг друга, издавая при этом характерные крики. Брачным полетом является хорошо знакомая охотникам тяга вальдшнепа, весеннее «блеяние» и полет бекасов. Особенно часто токовые полеты наблюдаются у птиц, обитающих на опушках леса и открытых пространствах: у лесного конька, серой и ястребиной славок, варакушки, камышевки-барсучка, белой куропатки и др. В схеме эти полеты выглядят так: самец взлетает по крутой траектории вверх, а затем по отлогой опускается вниз. Полет обычно сопровождается своеобразными звуками.
Позы «ухаживания» свойственны всем птицам. Характер поз у разных видов весьма разнообразен: приседания, поклоны, вытягивание и поворот шеи, своеобразные движения крыльев и хвоста и т. д. Для каждого вида свойствен определенный набор телодвижений, свой «ритуал», совершаемый самцом перед самкой. При этом самец старается выставить напоказ наиболее ярко окрашенные участки оперения, которые по способу употребления напоминают наши флаги, светофоры, маяки. У некоторых беззащитных птиц с покровительственной окраской яркие пятна обычно скрыты и становятся видными лишь в тот момент, когда это необходимо. Например, зарянка окрашена весьма скромно, но когда самец настроен враждебно, он распускает коричнево-красное оперение на груди, адресуя этот угрожающий сигнал своему противнику.
Звуки и песни — широко распространенная форма брачного поведения. К этому типу сигналов относятся издаваемые некоторыми птицами механические звуки — барабанная трель дятлов, громкое щелканье клювом аистов и др. Пение особой силы и разнообразия достигает у воробьиных птиц. Регулярное пение самцов может продолжаться в течение всего гнездового периода и даже после вылета птенцов из гнезда. Это связано с растянутостью сроков размножения у разных пар и особенно у видов, имеющих два выводка в сезон.[5]
Для каждого вида птиц характерно определенное, отличающееся от других, даже близких видов, брачное поведение — голос, позы и т. д. Это дает основание считать, что оно служит сигналом для других особей того же вида: усиливает возбуждение токующей птицы и одновременно привлекает и возбуждает брачного партнера. Именно в половом отборе и синхронизации половой активности партнеров и заключается в первую очередь биологическое значение брачных церемоний у птиц. Дело в том, что половой цикл у представителей разных полов начинается не одновременно: у самцов раньше, чем у самок. В связи с этим необходимо воздействие различных сменяющихся раздражителей. Начало развития семенников у самцов вызывается условиями освещения, увеличением продолжительности световой части суток (по крайней мере, у большинства видов умеренной и северной полос). Эти условия, но в меньшей степени, влияют и на самок, у которых развитие яичника наступает позднее, чем семенников у самцов, и, кроме световых условий, стимулируется брачным поведением самца (брачные игры, пение и т. д.). В это время от организма еще не требуется больших затрат энергии.
Но позднее, в период спаривания, когда активность самцов достигает максимума, светового фактора для осуществления размножения уже недостаточно. Половой цикл становится возможным лишь при благоприятном состоянии энергетического баланса организма, т. е. при определенных температурных и кормовых условиях. Известны случаи, когда внезапное ухудшение погоды вызывает не только прекращение полового цикла самцов, но и возвращение их половой системы к состоянию покоя. У самок в период яйцекладки затраты организма очень велики и могут быть обеспечены лишь при благоприятных условиях. В опытах удается прервать половой цикл самок путем понижения температуры, ухудшения питания, т. д.
Для осуществления яйцекладки необходим сложный комплекс условий: определенное состояние организма, такие внешние раздражители, как поведение партнера, присутствие гнезда и т. п., а также благоприятная температура и высококачественное питание (в частности, белковое и минеральное — последнее для образования скорлупы).
Таким образом, брачное поведение птиц имеет важное и многообразное значение. Оно облегчает встречу самцов и самок, помогает формированию пары у моногамов, способствует синхронизации половой активности партнеров и популяции в целом, подготавливает их к спариванию и гнездостроению. Одновременно у всех моногамов брачное поведение служит сигналом занятости гнездового участка: элементы брачного поведения используются при его маркировке и защите.
Для образования пар существенное значение имеет соотношение полов в природе. У разных видов птиц перед началом размножения оно близко 1:1, т. е. число самцов и самок примерно одинаково. Лишь у немногих видов число самцов в природе превышает в несколько раз число самок. Для этих видов характерна палеандрия, когда самка откладывает несколько кладок, которые насиживают самцы: они же водят и выводки. Полиандрия установлена у якан, трехперсток, некоторых тинаму. Лишь у немногих видов, наоборот, наблюдается преобладание самок. В этих случаях возникает явление полигамии, при котором строят гнезда и насиживают только самки.
Взаимоотношение партнеров у преобладающего большинства остальных птиц можно свести к двум типам: моногамии и полигамии. У некоторых, преимущественно крупных птиц, пары сохраняются в течение длительного времени. При этом у гусей, лебедей, журавлей самец и самка держатся вместе в течение круглого года, а у дневных хищников, некоторых сов, аистов, крупных чаек, после размножения пара распадается, но перед следующим размножением вновь восстанавливается, так как, благодаря хорошо развитому гнездовому консерватизму, оба партнера возвращаются на старый гнездовий участок.
Большинство моногамов образуют пары только на сезон размножения. При повторном размножении в тот же сезон часть пар сохраняется, другие формируются заново (большая синица, воробьи и др.).
Взаимоотношения партнеров у моногамных видов весьма различны. Эти различия основаны на разной степени участия самца в постройке гнезда, насиживании яиц, выкармливании птенцов. Как правило, основная забота в осуществлении этих процессов лежит на самке. В редких случаях сооружает гнездо только самец (кулик-перевозчик, сорокопут-жулан). У некоторых видов самец участвует в выведении потомства наравне с самкой (голенастые, трубконосые, чайки, городские ласточки и др.). Но много и таких видов, у которых самец только частично и в разных размерах принимает участие в постройке гнезда, насиживании и выкармливании птенцов (пеликаны, журавли, многие воробьиные). Есть немало видов, у которых самец регулярно кормит насиживающую самку (стрижи, удоды, клесты, иволги).
У некоторых типичных моногамов изредка встречается бигамия: на участке самца поселяются две самки. В некоторых случаях самец помогает в насиживании и выкармливании птенцов обоим самкам, но чаще — одной.
У некоторых видов пары сохраняются только на период спаривания и яйцекладки, после чего они распадаются, а дальнейшая забота о потомстве падает или только на самку (многие утки, беседковые птицы и др.), или только на самца (некоторые тинаму, трехперстки, яканы, кулики-плавунчики).
Полигамы — птицы, не образующие пар даже на короткий срок: самка может спариваться с разными самцами. Постройка гнезда, насиживание и забота о потомстве ложатся только на самку. К полигамам относятся большинство курообразных, дрофы, некоторые кулики (турухтан и др.), почти все колибри, большинство райских птиц и др.[6]
Территориальное поведение
Практически у всех птиц в период размножения хорошо выражена территориальность: привязанность к гнездовой территории и ее защита от вторжения особей своего, а иногда и других видов.
Самцы целого ряда видов птиц включают в ритуал привлечения самок обнаруженное ими место для гнезда. В качестве примера можно привести наблюдения за характерным поведением мухоловки-пеструшки [Muscicapa hypoleuca]. Обнаружив в середине мая пустую дуплянку, самец обследовал ее и начал усиленно петь и часто лазить в леток. Это продолжалось около полутора суток, причем самец пытался петь и около другой дуплянки, занятой большими синицами (Parus major), которые его активно гоняли. Через двое суток появилась самка мухоловка-пеструшка, которая заинтересовалась и самцом и его дуплянкой. Самец долго гонял самку (агрессивное поведение) и не давал ей залетать в дуплянку. Через 4—5 час он перестал ее гонять (в чем немалое значение имели ее умиротворяющие ритуальные движения) и допустил влезть в дуплянку, но только следом за собой. Еще через 2—3 часа самка начала носить в дуплянку материал для гнезда. При этом еще сутки самец пускал самку с материалом в дуплянку только после того, как он предварительно влетал туда и вылетал обратно. Лишь через сутки самка самостоятельно смогла беспрепятственно влетать в дуплянку. Можно было думать, что у самца угасло агрессивное поведение, но появившуюся еще через сутки вторую самку, которая тоже хотела осмотреть дуплянку, самец долго гонял и прогнал со своего участка. Таким образом, у наблюдателя создалось законное впечатление, что самец примерно через сутки научился узнавать именно свою самку и отличать ее от других. Агрессивность же его при защите своей гнездовой территории не угасла до момента вылета птенцов: любая залетевшая сюда «чужая» птица того же вида немедленно подвергалась нападению. Аналогичные наблюдения достаточно хорошо описаны в ряде статей и книг. Опыты подтверждают, что в защите гнездового участка нет права сильного, а есть право первого; хозяин, территории всегда победитель. Это же отметил А.Н. Промптов, указывая, что у воробьиных птиц при столкновениях из-за гнездового участка «победа, как правило, оказывается не на стороне сильного, а на стороне «законного владельца»; «агрессор», даже более сильный дерется слабее хозяина, отстаивающего свои права.[7]
При неколониальном (одиночном) гнездовании птицы гораздо меньше защищены от врагов и поэтому тщательно прячут и маскируют свои гнезда, не отлетая далеко от них за пищей. Последнее заставляет их тщательно охранять от конкурентов пригнездовой кормовой участок. При этом, конечно, можно наблюдать все переходы от колониальных птиц (чаек, чистиковых и др.) к птицам слабоколониальным и одиночным. Например, дрозды-рябинники (Turdus pilaris) могут гнездиться и одиночно, но предпочитают делать гнезда небольшими колониями. В них гнезда располагаются довольно далеко одно от другого, но птицы защищают их от врагов все вместе. По тревожному крику одной птицы немедленно взлетают в воздух и собираются на крик все члены этой колонии. Поэтому возможные враги — вороны, сороки, сойки и белки — быстро научаются избегать территории, занятой группой гнезд дроздов-рябинников.
У птиц-полигамов, как правило, защита гнездовой территории отсутствует. У многих из них самцы весной собираются в определенные места — токовища, где поют брачные песни и устраивают турниры, основное значение которых — половой отбор. Самки, собираясь к токовищам, спариваются с наиболее крепкими и сильными самцами. Места токовищ обычно довольно постоянны, и как самцы, так и самки собираются сюда с довольно больших расстояний. У некоторых видов брачные песни на токовищах слышны лишь на очень небольшом расстоянии, как, например, у глухаря (Tetrao urogallus), или кавказского тетерева (Lyrurus mlokosiewiczi), в то время как у других, как, например, у тетерева-косача (Lyrurus tetrix), токовища слышны на несколько километров. Последнее может служить для ориентации при нахождении токовищ для самок и самцов. Это может иметь особое значение при меняющемся месте токовища, что нередко можно наблюдать именно у тетерева. В то же время такое дальнее звучание токовища может представлять собой и определенную опасность для популяции, поскольку на него могут ориентироваться и такие хищники, как ястреб-тетеревятник (Accipiter gentilis), а в утренних сумерках — u филин (Bubo bubo). Можно думать, что именно поэтому тетеревиные токовища обычно располагаются на безлесных пространствах с большой площадью обзора.
Строгая оседлость птиц в период размножения возможна только при условии выбора такой гнездовой территории, на которой есть все необходимое для размножения: гнездилище, достаточное количество пищи (для взрослых и птенцов), надежные защитные условия, благоприятный микроклимат (температурный режим, влажность, освещенность) и др. Комплекс всех этих условий каждый вид птиц находит в том специфичном для себя гнездовом биотопе, в пределах которого выбирается гнездовый участок. Помимо всего прочего, эти условия оказывают влияние на нормальный ход физиологических процессов в организме в период размножения. Будучи источником внутренних (пища), а также внешних раздражителей (свет, температура, вид гнездилищ и привычной обстановки), комплекс условий стимулирует физиологическую подготовку организма и обеспечивает осуществление процесса размножения. Этот процесс протекает нормально только в том случае, если на гнездовой территории имеются все необходимые условия. При отсутствии хотя бы части из них (гнездилища, защитных условий и т. д.) гнездовая территория теряет стимулирующее значение и такую территорию птицы не заселяют. Подтверждением сказанному являются довольно частые случаи, когда не нашедшие благоприятных гнездовых участков половозрелые особи остаются холостыми.
По характеру распределения пар на гнездовой территории и по удаленности гнезд друг от друга всех птиц обычно разделяют на две группы: колониальные и одиночно гнездящиеся.
Из всех гнездящихся на территории бывшего СССР птиц примерно 1/6 часть гнездится колониально и 5/6 — одиночно. Каждая группа характеризуется своими особенностями взаимоотношений между соседними парами.
Колониальными называют птиц, гнезда которых располагаются близко одно к другому (чаще от нескольких сантиметров до нескольких метров), образуя гнездовые колонии. Из всех живущих в бывшем СССР колониальных видов 2/3 относится к водным и водно-болотным и только 1/3 — к сухопутным. В состав первой группы входят представители чистиковых, чайковых, веслоногих, аистообразных, поганок и др., в состав второй — ласточковых, ткачиковых, врановых, длиннокрылых и др.[8]
Величина колоний у разных видов разнообразна. У морских птиц она достигает десятков и сотен тысяч гнезд, у обитателей пресных водоемов — тысяч, редко десятков тысяч, у сухопутных — обычно десятков и сотен гнезд. Эти различия связаны с неодинаковым количеством пищи и с разной обеспеченностью удобными гнездилищами.
Колониальное гнездование возникло у тех видов птиц, у которых имеет место спорадичное размещение в местах с богатой кормовой базой. Например, птичьи базары кайр существуют там, где благодаря оптимальному гидрологическому режиму формируется богатая биогенами зона, способствующая развитию планктона и нектона, и в первую очередь рыбы.
Самая крупная в мире колония бакланов и олушей, насчитывающая 10 млн. птиц, расположена на побережье Перу, где к самому берегу подходит холодное Перуанское течение, изобилующее анчоусами, которыми питаются птицы. Плотность в колонии достигает 4 гнезд на 1 кв. м. Благодаря богатой фосфором и азотом пищи помет птиц — гуано — лучшее из естественных удобрений в мире. Птицы на этом побережье откладывают за 100 лет слой гуано примерно в 15 см толщиной. Его залежи там достигают 50 м. Это ценное удобрение издавна используется в сельском хозяйстве, но особенно интенсивные разработки ведутся в настоящее время.
Существенную роль в возникновении колониального гнездования у птиц играет также самозащита. Хищнику трудно приблизиться к гнездам незамеченным, так как в колонии всегда находятся птицы. Кроме того, даже крупные хищники не могут противостоять организованному массовому нападению со стороны обитателей гнездовой колонии и быстро удаляются за ее пределы. Коллективный отпор врагу обитателей гнездовых колоний можно наблюдать довольно часто даже у мелких воробьиных птиц. Скворцы, например, по первому сигналу тревоги быстро собираются целой группой и с отчаянными криками прогоняют прилетевшую к скворечнику ворону. Деревенские ласточки с пронзительными воинственными криками успешно преследуют появившуюся у сарая, под крышей которого расположены их гнезда, кошку, не выдерживающую нападения пикирующих на нее птиц и моментально убегающую.
К защите колониальных видов прибегают и некоторые одиноко гнездящиеся птицы. Так, на островах Балтийского моря около колоний полярной крачки нередко устраивают свои гнезда хохлатая чернеть, черношейная поганка, камнешарка. У колоний серебристых чаек встречаются гнезда обыкновенной гаги. О защитной роли гнездовых колоний можно судить по следующему примеру. В одной местности хищники разорили почти половину (43%) гнезд обыкновенной гаги, расположенных вне колонии, а в колонии — лишь 13% У ряда видов птиц колониальность могла возникнуть и вследствие недостатка подходящих мест для постройки гнезд. Примером таких видов может служить береговая ласточка, размещающая свои гнездовые норы на дефицитных береговых обрывах, карьерах и т. д.
Из-за концентрации большого числа особей в одном месте птицам нередко приходится совершать полеты за кормом на значительное расстояние от гнездовой колонии. Дальность таких полетов у колониальных птиц гораздо большая, чем у большинства одиночно гнездящихся. Розовые скворцы в период выкармливания птенцов улетают за кормом на расстояние 5-10 км от гнезда, чайки — на 20-30 км, альбатросы — даже на сотни километров, в то время как одиночно гнездящиеся птицы обычно собирают корм поблизости от гнезда. Большинство колониальных птиц являются хорошими летунами, например чайки, трубконосые, длиннокрылые, из воробьиных — ласточки и др. Исключение составляют виды, имеющие вблизи от колоний действительно неистощимые запасы корма (чистиковые или же гнездящиеся мелкими группами воробьи, коноплянки и др.).
Одной из особенностей колониальных птиц, обеспечивающей успешное выкармливание птенцов в условиях некоторой отдаленности гнезд от кормовой базы, является способность приносить птенцам корм к гнезду сразу большими порциями. Птицы приспособились при сборе корма сначала его заглатывать, а прилетев к гнезду, отрыгивать и давать птенцам. Так поступают чайки, бакланы и др. Птицы, не проглатывающие корм, набирают в клюв обычно гораздо большую его порцию, чем птицы, отыскивающие пищу поблизости от гнезда. Благодаря этой особенности сокращается число полетов колониальных птиц за кормом.
К одиночно гнездящимся относятся птицы, располагающие свои гнезда на значительном расстоянии одно от другого. При этом расстояние между соседними гнездами у разных видов весьма различно и может колебаться от нескольких десятков метров (кулики, мелкие воробьиные) до нескольких километров (крупные хищники). В отличие от колониальных птиц соседние пары здесь не связаны между собой и ведут более или менее изолированный образ жизни. К этой группе принадлежит абсолютное большинство птиц.
Главное преимущество одиночно гнездящихся птиц состоит в том, что хищникам труднее обнаружить редкие и обычно хорошо замаскированные гнезда, чем колониальные. Кроме того, для некоторых птиц одиночное гнездование связано с преимуществом поиска корма в одиночку.
У некоторых видов наблюдается промежуточный тип гнездования, так называемый групповой, при котором гнезда располагаются дальше друг от друга, чем в колонии (от нескольких метров до десятков метров), и ближе, чем у одиночно гнездящихся. К таким видам относятся дрозды-рябинники, скворцы и др. Корм эти птицы собирают за пределами охраняемой территории. При приближении хищника к такому гнездовью на него нападают все члены группы.
Привязанность типа к местам гнездования (гнездовый консерватизм и дисперсия). Вопрос о гнездовом консерватизме птиц до последнего времени остается в известной мере дискуссионным. Накопленные орнитологами данные о территориальном поведении птиц достаточно противоречивы. С одной стороны, имеется много фактов, свидетельствующих о большом постоянстве мест гнездования птиц, об их способности ежегодно возвращаться на свое место с зимовок. На основании этого многие орнитологи стали считать, что гнездовый консерватизм — универсальное явление, свойственное, за редким исключением, всем птицам. С другой стороны, накопилось немало данных, показывающих обратную картину: смену значительной частью птиц мест гнездования вплоть до выселения в другие географические области. Учитывая это, ряд орнитологов выступают против универсальности гнездового консерватизма.
Вопрос о степени привязанности птиц к своим постоянным местам гнездования имеет в современной орнитологии важное значение. От правильного его решения зависит объективность наших представлений о природе популяций птиц, а в конечном итоге — о ходе у них эволюционного процесса.
На основании изучения экологии отдельных видов, и особенно кольцевания, сторонники гнездового консерватизма пришли к заключению, что не только старые птицы из года в год сохраняют за собой раз уже занятый гнездовой участок, но и молодые птицы (первогодки) гнездятся преимущественно в непосредственной близости от того места, где они вывелись. По их убеждению, правило регулярного возврата на родину почти в одинаковой мере применимо как к старым, так и к молодым птицам. При этом «родину» понимали достаточно узко — как территорию лесного массива, лесопарка, долины и т. п. Все случаи нахождения окольцованных птиц гнездящимися вдали от мест их рождения стали расценивать как исключение, а особей, не возвратившихся на родину, считать в основном погибшими. В итоге возникло представление о чрезвычайно высоком проценте гибели птиц на первом году их жизни (90-95%).
Биологический смысл строгой привязанности к постоянным местам гнездования заключается в том, что местные «микропопуляции» лучше приспособлены к конкретным местным условиям жизни, ибо рефлексы и навыки, облегчающие им существование в этих условиях, легче всего создаются на первом году жизни.
Местная популяция из года в год населяет определенную территорию и пополняется главным образом за счет своих потомков. Таким образом, в понятие «элементарной» или «микропопуляции» вкладывается определенный генетический смысл, предполагающий большую изоляцию от других популяций.
Наряду с этим за последнее время накоплено немало прямых и косвенных доказательств того, что помимо консервативной стороны поведения птицам свойственна и значительная подвижность территориальных связей в пределах гнездовой области (дисперсия). Стало очевидным, что у птиц существует явление расселения и постоянного довольно широкого генетического обмена между группами особей, населяющих соседние районы. Наблюдения показывают, что расселение, пожалуй, не менее широко распространенное в природе явление, чем гнездовой консерватизм. Весной и в начале лета расселение птиц можно наблюдать ежегодно. Оно становится особенно заметным при изучении процесса заселения птицами биотопов, вновь возникающих в результате хозяйственной деятельности человека (полезащитные лесные полосы, искусственные водохранилища и т. д.), а также при анализе большинства достаточно многочисленных случаев освоения птицами новых территорий за пределами гнездового ареала.
К этой же категории явлений следует отнести хорошо известные факты так называемой «абмиграции» уток,[9] у которых формирующиеся на зимовках пары нередко складываются из партнеров, прилетевших из разных, весьма отдаленных друг от друга географических областей; в результате один из партнеров улетает гнездиться в новую для себя область. Происходящий в природе процесс перемещения особей в гнездовой области выявляется также при многолетних стационарных исследованиях с применением метода массового кольцевания птенцов. Этим не заканчивается перечень примеров достаточно высокой у птиц территориальной подвижности, хотя явление расселения — наименее изученная сторона их биологии.
В работах последних десятилетий показано, что гнездовый консерватизм характерен преимущественно для оседлых, удачно размножавшихся и некоторой части молодых птиц. Возвращаясь на старые места гнездования, они оказываются в более выгодном положении, так как могут использовать здесь преимущества опыта в привычных для них условиях. Стремление к поиску новых мест, расселение свойственно преимущественно молодым особям. В первую очередь за счет них расширяется ареал, заселяются новые биотопы, происходит перемещение населения соседних «микропопуляций».
Степень выраженности у разных видов и количественные отношения явлений территориального консерватизма и дисперсии в природе остаются еще недостаточно изученными. Тем не менее накопившиеся к настоящему времени фактические данные дают основание признать «права и гражданство» не только за гнездовым консерватизмом, но и за дисперсией.
Таким образом, расселение особей в пределах гнездовой области не менее характерная черта биологии птиц, чем тяготение к постоянным местам гнездования. По биологическому смыслу эти оба явления противоположны. Консерватизм, свойственный в основном старым особям, сохраняет эволюционные завоевания вида. Подвижность (дисперсия), выражающаяся главным образом в расселении молодых птиц, обусловливает возможность новых завоеваний. Препятствуя изоляции и осуществляя панмиксию, дисперсия способствует повышению общего уровня экологической пластичности особей и всей организации вида на широких пространствах. Равновесие двух сторон обеспечивает стабильное существование вида.
Гнездостроение и гнезда птиц
На определенной стадии развития полового цикла птицы приступают к постройке гнезда. Гнездостроение — особенность, характерная для птиц, появилась впервые среди позвоночных животных. Так называемые «гнезда» у некоторых представителей низших классов позвоночных, служащие главным образом для охраны яиц, — явление, принципиально отличное от гнезд птиц. Значение птичьих гнезд более разнообразно и касается не только охраны яиц, но и развития зародышей (эмбрионального и постэмбрионального).
Понятие «гнездо» для некоторых птиц в известной мере условно, так как не все они строят себе настоящие гнезда. Прямо на голый карниз кладет свое единственное яйцо обитатель птичьих базаров кайра.
Не делает какого-либо подобия гнезда козодой, помещая два яйца на естественную лесную подстилку. Не строят гнезда и совы — они несут яйца прямо на землю или в готовое дупло. На такие гнезда птицы совершенно не затрачивают ни времени, ни усилий. Но подобных птиц мало. Преобладающее большинство птиц имеют гнезда, на сооружение которых затрачиваются специальные усилия и время.
Настоящими следует считать также гнезда, постройка которых сводится к расчистке птицами лишь небольшого углубления в почве (глухарь, рябчик и др.). Однако было бы неправильно считать, что у кайры, козодоя и других птиц, откладывающих яйца на естественный грунт, нет гнезда. Гнездом обычно принято считать то место, на которое отложены яйца и на котором происходит их насиживание и выкармливание птенцов. Будет ли это место естественным субстратом или же специальным сооружением птицы, значение его для процесса размножения остается тем же.
Есть, кроме того, птицы, которые сами гнезда не строят, а используют гнезда других птиц. Так, соколы и некоторые другие дневные хищники занимают гнезда вороны, грача, канюков, сороки и др.
Так или иначе, но гнезда имеют все птицы. Исключения редки и относятся к видам, обладающим гнездовым паразитизмом. Эти виды своих гнезд не имеют и яйца подкладывают в гнезда других птиц.
Значение птичьих гнезд
Гнезда у птиц выполняют разнообразные и очень важные функции:
- гнездо предохраняет яйца от раскатывания и удерживает их в компактной кучке, благодаря чему вся кладка умещается под телом насиживающей птицы и прогревается;
- у птенцовых птиц гнездо служит местом нахождения птенцов в период их выкармливания до их вылета;
- гнездо способствует созданию оптимального для развития яиц и птенцов температурного режима, предохраняя их от ветра, дождя, сырости, холода и других неблагоприятных условий;
- гнездо играет большую роль в защите яиц, птенцов и насиживающей птицы от врагов, что достигается маскировкой гнезда и расположением его в недоступных для врагов местах.
Рассмотрим каждую из этих функций в отдельности.
1. Удержание яиц в компактной кучке особенно важно для видов с обильной кладкой, что обеспечивает возможность удерживать ее под наседкой и осуществлять нормальную инкубацию.
Глубина гнезд у разных видов весьма различна, но достаточна для того, чтобы яйца полностью помещались в гнезде. Интересное и парадоксальное исключение относится к упоминавшейся выше кайре. Свое единственное яйцо кайры кладут на уступы голых скал без какой-либо подстилки и углубления. Яйцо легко умещается под телом птицы, и проблема предохранения яиц от раскатывания здесь как-будто отпадает. Но отсутствие углубления вызывает другую опасность: при малейшем неосторожном движении птицы, например при поспешном взлете с гнезда, яйцо легко может скатиться с узкого карниза скалы и разбиться. И действительно, яйца кайр иногда разбиваются, падая от толчков птиц и порывов ветра. Но число таких случаев не так велико, как этого можно было бы ожидать, благодаря своеобразной грушевидной форме яйца — тупого с одного конца и острого — с другого. При толчке яйцо не катится, а вращается вокруг острого конца на одном месте, предотвращая таким образом падение с гнездового карниза. Падение яиц можно наблюдать чаще в начале периода размножения, в конце же такая гибель яиц почти полностью прекращается. Объясняется это теми изменениями, которые происходят в яйце в процессе насиживания. В начале насиживания центр тяжести яйца, вследствие незначительных размеров воздушной камеры, находится ближе к тупому концу. По мере развития зародыша воздушная камера в яйце постепенно увеличивается, и центр тяжести перемещается в сторону острого конца. Благодаря этому по мере насиживания радиус окружности, описываемой яйцом при толчке, а следовательно и вероятность его падения с карниза, уменьшается. Повышение устойчивости яйца в конце насиживания имеет большой биологический смысл, так как по мере насиживания возможность возобновления птицами пропавших кладок резко снижается.
Птицы, имеющие средние и крупные кладки, как, например, утки, тетерева, куропатки и многие другие, яйца размещают в гнезде не в один, а в два ряда. В таких кладках при насиживании не все яйца прогреваются одинаково: верхние лучше, нижние — хуже. Казалось бы, вследствие этого и развитие зародышей в яйцах должно проходить неравномерно, а птенцы выклевываться неодновременно. Однако у птиц, приступающих к непрерывному насиживанию по окончании всей кладки, этого не наблюдается. Даже в гнездах с кладкой 15-16 яиц, например у тетерева и белой куропатки, появление птенцов происходит, как правило, в течение одних лишь суток.
Обусловлено это тем, что птицы в период насиживания систематически перемещают яйца в гнезде из нижнего ряда в верхний, с краев кладки в центр, обеспечивая тем самым равномерное прогревание всей кладки. Таким образом, у птиц с крупной кладкой в гнезде, где яйца находятся в компактной кучке, создаются известные трудности для равномерного прогревания яиц. Эти трудности устраняются путем систематического перемещения яиц, производимого насиживающей птицей.
2. Использование гнезда как места нахождения птенцов в период выкармливания родителями свойственно только птенцовым птицам. В связи с этим относительные размеры гнезд у птенцовых, как правило, крупнее, чем у выводковых. В связи с относительно небольшой величиной гнезд обильные кладки выводковых птиц (рябчиков, тетерева, куропаток и др.) обычно полностью заполняют гнездо. Вышедшие из этих яиц птенцы через несколько часов (как только обсохнут) покидают гнездо. Выводковые птицы используют гнездо только для насиживания яиц.
У птенцовых птиц гнездо глубокое, вместимость его больше требуемой для размещения яиц. Вылупившиеся птенцы находятся в гнезде до тех пор, пока не приобретут известную самостоятельность. Гнездо, таким образом, служит не только для размещения яиц, но и для выращивания птенцов.
Однако у разных биологических групп птенцовых птиц относительная величина гнезд (точнее, вместимость) неодинаковая. Так, у птиц, гнездящихся на земле, они более мелкие, чем у видов, располагающих гнезда на деревьях. Эта разница связана с тем, что подросшие птенцы гнездящихся на земле птиц обычно покидают гнездо, еще не умея летать, чего не могут сделать птенцы из гнезд, расположенных на деревьях.
3. В гнезде поддерживается оптимальный для развития яиц и птенцов температурный режим. Это свойственно большинству птиц и имеет большое значение для успешного воспроизведения потомства. Свои гнезда птицы почти всегда устраивают на сухой почве и из сухого строительного материала. Правда, деревенская и городская ласточки лепят гнезда из грязи и глины, смешивая их с травинками, фламинго — из ила. Но эти сооружения заселяются только после их просыхания. К тому же внутри они обычно имеют подстилку из сухого материала.
В зависимости от влажности среды толщина подстилки, дна и стенок самого гнезда бывает различной. В этом легко убедиться, просматривая серию гнезд лесных коньков, дроздов-белобровиков и других гнездящихся на земле птиц. Чем сырее почва, тем толще стенки и особенно дно гнезда, и наоборот. Особенно наглядны в этом отношении гнезда озерных чаек, размещаемые на плавнях заболачиваемых озер. В тех случаях, когда гнездо чайки сделано на сухой почве, толщина его дна составляет 2-4 см. Но чем сырее место, тем основание гнезда плотнее и дно толще: на ровной сырой почве плавней толщина его достигает 12-15 см. Сухой материал, толстое дно и стенки гнезда предохраняют яйца и птенцов от сырости, создают более устойчивую температуру в гнезде, что очень важно для нормального насиживания, а также для развития птенцов.
4. Велико значение гнезд в охране яиц и птенцов от врагов. Даже у взрослых птиц много врагов, а их хрупкие яйца и слабые беспомощные птенцы служат лакомой добычей для многих животных. Не случайно, что большое число погибает в первые дни, недели и месяцы их жизни.
Защита гнезд осуществляется птицами разнообразными способами. Одни птицы вьют гнезда в недоступных для многих врагов местах: на высоких деревьях (дневные хищники, цапли, зяблики, дрозды, голуби, грачи), на скалах (кайры, чистики, некоторые чайки), на высоких зданиях (стрижи, ласточки), в дуплах (дятлы, поползни, некоторые синицы), в земляных норах (береговая ласточка, зимородок) и пр.
Другие птицы хотя и устраивают гнезда на земле, на низких кустарниках и в иных доступных для своих врагов местах, но они настолько хорошо маскируются, что практически обнаружить врагам их трудно. Особенно большую роль в маскировке гнезда играет его наружная отделка. Для нее птицы инстинктивно выбирают такой материал, который наиболее хорошо гармонирует с окружающей обстановкой и делает гнездо незаметным. Важна для маскировки также покровительственная окраска яиц и самих насиживающих.
У дуплогнездников и других закрыто гнездящихся птиц, у которых кладка яиц спрятана от посторонних взоров в дупла, земляные норы и иные закрытые места, окраска яиц имеет более яркие тона, без пятен и крапинок. Исключение представляют синицы, которые хотя и гнездятся в дуплах, но яйца их имеют пеструю окраску. Сравнительно яркая окраска яиц наблюдается и у некоторых открыто гнездящихся видов, но только в тех случаях, когда насиживающая птица при уходе с гнезда на кормежку прикрывает яйца тем или иным маскирующим материалом. Так, например, утки маскируют яйца защитно окрашенным пухом, выщипанным из груди; некоторые куриные — листьями; поганки — водяными растениями, выдернутыми из воды по соседству с гнездом.
Многие птицы в тропиках строят свои гнезда рядом с гнездами ос или устраивают их на деревьях, населенных муравьями. Такой способ надежно защищает их гнезда от нападения мелких хищников. Мелкие птицы нередко гнездятся вблизи гнезд крупных и сильных птиц, которые, защищая свои гнезда, обеспечивают сохранность гнезд и мелких птиц. К этому, например, прибегают воробьи, устраивающие свои гнезда в стенах гнезд орлов, орланов и скоп, а ушастые поганки гнездятся под защитой колоний чаек или крачек.
К числу приспособлений следует отнести и ту особенность, что птенцы открыто гнездящихся птиц не подают голоса, в то время как находящиеся в безопасности птенцы дуплогнездников дружно пищат при каждом прилете родителей с кормом.
Яйцо и его особенности у птиц
Отложенное яйцо имеет плотную наружную оболочку — известковую скорлупу, прикрытую тонкой кутикулярной надскорлуповой оболочкой, предохраняющей яйцо от проникновения в него через поры в скорлупе различных микроорганизмов. Под скорлупой находятся две тонкие пергаментообразные подскорлуповые оболочки, обволакивающие белок. На тупом конце яйца подскорлуповые оболочки расслаиваются, образуя воздушную камеру. Далее располагается толстая белковая оболочка, прикрывающая шаровидный желток. На анимальном полюсе желтка находится зародышевый диск. От внутренней подскорлуповой оболочки к желтку идут жгуты плотного белка — халазы. Свободно поворачивающийся на халазах желток при любом положении яйца обеспечивает зародышевому диску всегда местоположение наверху. Все оболочки формируются при прохождении яйца по яйцеводу.
Желток содержит основной запас питательных веществ, идущих на формирование тканей зародыша, на обеспечение его основных энергетических затрат и частично — потребности в воде. Белковая оболочка — основной источник необходимой для зародыша воды и лишь отчасти добавочный резерв энергетических веществ.
Количество желтка по отношению к белку у птиц очень различно: у вертишейки и баклана оно составляет 15%, у некоторых уток — 50%, у птенцовых птиц — (воробьиных, голубей) — 20%, у выводковых (кур, уток) — 35%. У птенцовых (например, бакланов и дятлов) весь желток к моменту выхода птенцов из яйца расходуется полностью, а у выводковых (например, лебедей и поганок) почти треть желтка остается в полости брюха вылупившихся птенцов как запас пищи для первых дней жизни, когда они еще не могут обеспечить себя кормом. Жир желтка необходим им также и для терморегуляции при похолодании и других неблагоприятных условиях.
Период времени между откладыванием отдельных яиц неодинаков: у большинства птиц он составляет 24 часа, у крупных птиц — 72 часа, у наиболее крупных среди них (у бородача) — 120 часов.
Размеры яиц у разных видов сильно колеблются не только абсолютно, но и относительно величины самой птицы. Как правило, мелкие птицы откладывают относительно крупные яйца. У птенцовых птиц яйца мельче, чем у выводковых. У последних птенцы появляются на свет более развитыми. В крупных кладках яйца относительно мельче, чем в мелких. Минимальные размеры яиц характерны для кукушек. Так, наша обыкновенная кукушка имеет массу примерно такую же, как и бекас (т. е. около 100 г), но масса яйца у бекаса в среднем составляет 17 г, а у кукушки — всего лишь 3 г. Мелкий размер яиц у кукушки обусловлен приспособлением к гнездовому паразитизму: свои яйца она откладывает в гнезда мелких воробьиных птиц.
Наибольшие размеры имело яйцо не столь давно вымершего крупного мадагаскарского эпиорниса. Его масса составляла 10 кг 460 г, длина 340 мм, ширина 245 мм, масса скорлупы достигала 3 кг 846 г.
У разных видов варьирует соотношение массы всей кладки яиц к массе тела. У большой поганки (с массой тела около 1 кг) вся кладка из 4 яиц составляет 12-14% массы тела; у полярной гагары (с массой 4 кг) кладка из 2 яиц равна 12-13% общей массы самки; у кулика-турухтана (масса 100 г) кладка из 4 яиц составляет 88% массы птицы. У некоторых птиц масса кладки даже превосходит массу самой самки. Так, у обыкновенного погоныша масса кладки из 12 яиц равна 125% массы самки, у кулика-перевозчика (в кладке 4 яйца) — 117%, у королька (в кладке 11 яиц) — 120%, у утки-гоголя — 110%; у утки рыжей савки, имеющей массу примерно 0,5 кг, масса ее средней кладки из 9 яиц равна почти 1 кг, а крупной — из 14-15 яиц достигает 1,5 кг.[10]
Приведенные цифры — показатель того, как много органического вещества и энергии в некоторых случаях расходуется организмом птицы при откладывании яиц.
Форма яиц — чаще овальная, но имеется ряд модификаций.
Выделяют пять типов форм яиц: равномерно эллиптические по обоим концам — у козодоев, колибри, рябков; равномерно эллиптические с двумя заостренными концами — у казуаров, нанду; почти круглые — у сов (кроме сипухи), щурок, зимородков; эллиптические с одним тупым концом и заостренным другим — у большинства птиц отряда воробьиных, куриных, дневных хищников и др.; конические, грушевидные — у куликов, чистиков.
Форма яиц и их размеры обусловлены объемом и формой яйцепроводящих путей. Птицы с узким тазом имеют только узкие яйца.
Окраска яиц соответствует отложению пигментов в различных слоях скорлупы при прохождении яйца через особый отдел яйцевода.
По окраске яйца можно разбить на три группы: неокрашенные, одноцветно окрашенные, пестрые.
Неокрашенные яйца свойственны большинству голубей и непаразитирующим кукушкам, а также видам, гнездящимся в дуплах и других закрытых местах. У ряда этих видов на время отсутствия насиживающей птицы на гнезде яйца прикрываются пухом или растительной ветошью.
Одноцветные яйца бывают окрашены в желтый (у голубей), черный (у некоторых воробьиных), но чаще — в голубоватый и зеленоватый цвета с разными оттенками (у уток и др.).
Пятнистая окраска имеет наибольшее распространение; у открыто гнездящихся птиц она хорошо гармонирует с окружающими предметами и выполняет функцию маскировки.
Окраска яиц в пределах вида в общем однотипна, но подвержена индивидуальным вариациям. Одна и та же самка кладет весьма однообразные яйца, хотя в природе нет двух совершенно одинаковых яиц.
В окраске яиц довольно четко выражена географическая изменчивость, особенно у птиц с яйцами пестрой окраски. Изменчивость эта ярче проявляется у птиц со слабыми индивидуальными колебаниями в окраске и размерах яиц.
Вскармливание потомства
У птиц при вылуплении птенцов происходят новые резкие изменения поведения и активности птиц-родителей, происходит смена значений раздражителей в связи с переключением доминант периода насиживания и выкармливания. В период выкармливания птенцов происходят существенные перемены в общей активности взрослых птиц, так как воспитание потомства требует от родителей большой затраты сил.
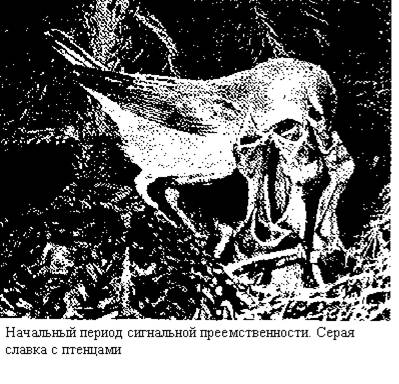
После вылупления мощнейшим сигнальным фактором становятся именно птенцы. Гнездо теряет значение «ключевого раздражителя».[11] При этом выводковые птицы (зрелорождающиеся) покидают гнездо в короткие сроки после вылупления, а птенцовые (незрелорождающиеся) остаются в нем довольно длительный срок. У первых почти сразу после вылупления четко действует рефлекс следования за самкой (или за обоими родителями), подчинение их сигналам (например, сбора, затаивания и т. д.), рефлекс подражания родителям и использования их для обогревания. Словом, у этой группы птиц сразу после выклева начинается активный контакт между родителями и потомством. На основе этого контакта образуются прочные семейные группы, примитивные формы которых существуют у рыб.
У вторых, рождающихся слепыми и беспомощными, первоначально действуют лишь безусловные рефлексы открывания рта и дефекации. Соответственно родительское поведение у этой группы птиц значительно сложнее, а контакт между родителями и потомством и прочные семейные группы здесь образуются и развиваются постепенно и сложно. Поскольку птенцовые птицы вылупляются из яиц слепыми, глухими и совершенно беспомощными, то их разделяют их ювенильный период жизни на три естественные стадии. Первой стадия - от момента вылупления до прозревания птенцов, когда последние еще лишены дистантной рецепции и преобладающими формами их поведения являются врожденные элементы. Второй стадией определяется период от включения зрительной и слуховой рецепции птенцов до оставления ими гнезда. В этот период у птенцов формируются многие начальные жизненно важные реакции. Наконец, третья стадия — от оставления гнезда до начала самостоятельной жизни. Для каждой стадии характерны особенности поведения как птенцов, так и родителей. Общей чертой всех стадий является кормление родителями птенцов. При этом резко возрастает активность сбора пищи при одновременном уменьшении ее потребления самими родителями. На первых двух стадиях происходит также уход за птенцами: чистка их покровов и вынос фекалия.
На всех трех стадиях родители охраняют и защищают птенцов самыми разными способами. Птицы ряда видов защищают потомство, самоотверженно и яростно нападая на приближающихся к гнезду врагов. Это характерно для многих хищных птиц, пускающих в ход когти и клювы. Чайки, крачки, а также дрозды-рябинники нападают на хищных птиц и зверей целой стаей и ударами с налета обычно обращают их в бегство. При нападении на приближающегося к гнезду человека они налетают на него и довольно точно попадают своими фекалиями. Чибисы около гнезд налетают на собак, лисиц и хищных птиц. Активно защищают своих птенцов (в гнездах) дрозды - белобровики в отличие от певчих и черных дроздов. Такая форма защиты потомства, характерна и для ряда рыб. Другая форма защиты заключается в затаивании самки на гнезде или около выводка и внезапном вылете на очень близком расстоянии от хищника (или человека). Часто при этом наблюдается так называемая «отвлекающая демонстрация. Она заключается в том, что птица так или иначе имитирует доступный для хищника пищевой объект, отвлекая его от гнезда или от выводка. Различают в этих отвлекающих демонстрациях несколько форм: 1) имитация убегающего зверька, когда птица бежит по земле с взъерошенными перьями спины, втянутой шеей и опущенным к земле хвостом; 2) имитация птенца с трепещущими крыльями и громким тонким ювенильным писком; 3) имитация раненой птицы с подлетами и падением на землю. Все эти формы известны и у других видов птиц, гнездящихся на земле или низко над землей. Так, например, хорошо известны отвлекающие демонстрации «раненой» птицы у самки рябчика около выводка. Элементы отвлекающего полета можно видеть у некоторых пеночек и славок.
Список использованной литературы
1. Баскин Л.М. Этология стадных животных. М., 1986.
2. Вагнер В.А. Психология размножения и эволюция//Сравнительная психология и зоопсихология: Хрестоматия/сост. Г.В. Калягина. Спб., 2001.
3. Гриффин Д. Перелеты птиц – М., Мир, 1966 г.
4. Дарвин Ч. Происхождение видов путем естественного отбора. М.; Л., 1937.
5. Зорина З.А., Полетаева И.И., Резникова Ж. И. Основы этологии и генетика поведения. М., 2002.
6. Карри-Линдалл К. Птицы над сушей и морем – М., Мысль, 1984 г.
7. Лоренц К. Агрессия (так называемое "зло"). М., 1994.
8. Михеев А.В. Биология птиц. Полевой определитель птичьих гнезд. Пособия для студентов пединститутов и учителей средних школ. М.: Топикал, 1996.
9. Михеев А.В. Перелеты птиц – М., Лесная промышленность, 1981 г.
10. Наумов С.П. Зоология позвоночных – М., Просвещние, 1973 г.
11. Панов Е.Н. Общение в мире животных. М., 1970.
12. Панов Е.Н. Сигнализация и "язык" животных. М., 1970.
13. Панов Е.Н. Механизмы коммуникации у птиц. М., 1978.
14. Панов Е.Н. Поведение животных и этологическая структура популяций, М,, 1983.
15. Панов Е.Н. Бегство от одиночества, М., 2002.
16. Птицы. Животный мир СССР. Бутурлин С.А., Гептнер В.Г., Дементьев Г.П., Житков Б.М., Огнев С.И., Промптов А.Н., Туров С.С., Формозов А.Н., Шульпин Л.М. Под ред. А.Н.Формозова и Б.М. Житкова. Москва - Ленинград, Детиздат, 1940.
17. Растения и животные: Руководство для натуралиста. Пер. с нем./ К.Нидон, д-р И. Петерман, П. Шеффель, Б.Шайба. М., Мир, 1991.
18. Северцов А.С. Введение в теорию эволюции. М.: Изд-во МГУ, 1981.
19. Слоним А.С. Инстинкт. Л., 1967.
20. Смирин В.М. Смирин Ю.М. Звери в природе, М., 1991.
21. Тинберген Н. Осы, птицы, люди. М., 1970.
22. Тинберген Н. Мир серебристой чайки. М., 1974.
23. Шилов И.А. Эколого-физиологические основы популяционных отношений у животных. М., 1977.
24. Шовен Р. Поведение животных. М., 1972.
25. Чельцов-Бебутов А.М. Экология птиц - М., Издательство МГУ, 1982 г.
26. Хрестоматия по зоопсихологии и сравнительной психологии/сост. М.Н. Сотская Учебное пособие МГППУ, 2003.
[1]Птицы. Животный мир СССР. Бутурлин С.А., Гептнер В.Г., Дементьев Г.П., Житков Б.М., Огнев С.И., Промптов А.Н., Туров С.С., Формозов А.Н., Шульпин Л.М. Под ред. А.Н.Формозова и Б.М.Житкова. Москва - Ленинград, Детиздат, 1940, стр. 120-121.
[2] Панов Е.Н. Механизмы коммуникации у птиц. М., 1978., стр. 17-18
[3] Вагнер В.А. Психология размножения и эволюция//Сравнительная психология и зоопсихология: Хрестоматия/сост. Г.В. Калягина. Спб., 2001., стр. 34.
[4] Панов Е.Н. Сигнализация и "язык" животных. М., 1970, стр. 56-57.
[5] Панов Е.Н. Сигнализация и "язык" животных. М., 1970, стр. 60-62.
[6] Панов Е.Н. Бегство от одиночества, М., 2002, стр. 172-173.
[7] Птицы. Животный мир СССР. Бутурлин С.А., Гептнер В.Г., Дементьев Г.П., Житков Б.М., Огнев С.И., Промптов А.Н., Туров С.С., Формозов А.Н., Шульпин Л.М. Под ред. А.Н.Формозова и Б.М.Житкова. Москва - Ленинград, Детиздат, 1940, стр. 173-174.
[8] Растения и животные: Руководство для натуралиста. Пер. с нем./ К.Нидон, д-р И.Петерман, П.Шеффель, Б.Шайба. М., Мир, 1991, стр. 34-35.
[9] Панов Е.Н. Поведение животных и этологическая структура популяций, М,, 1983, стр. 78-79.
[10] Хрестоматия по зоопсихологии и сравнительной психологии/сост. М.Н. Сотская Учебное пособие МГППУ, 2003, стр. 220-222.
[11] Михеев А.В. Биология птиц. Полевой определитель птичьих гнезд. Пособия для студентов пединститутов и учителей средних школ. М.: Топикал, 1996, стр. 37-38.
Похожие работы

... отличаются обычно высокой численностью и большой экологической устойчивостью. Можно вслед за другими исследователями установить два основных типа несемейных группировок животных, которые различаются по особенностям взаимоотношении и поведения животных, составляющих группу: 1) с доминированием одних особей над другими (такие группы называются иерархическими); 2) стаи без доминирования. В ...
... из них обладают некоторым внутренним ритмом, или часами, для предсказания периодических изменений и подготовки к ним. Существуют три основных способа синхронизовать физиологию и поведение с циклическими изменениями среды: 1) прямой ответ на разные изменения во внешних геофизических стимулах; Рис.6. Векторно-аддитивные процессы при физиологической регуляции. Вектор акклиматизации а ...
... обычной в то время сельди – каспийского пузанка (Caspialosa caspia caspia). Волжская сельдь (C. kessleri volgensis) и черноспинка (C. k. kessleri) до настоящего времени в пищевом спектре этих птиц из Саратовской области занимают немаловажное место: в 1-й и 2-й декадах июля в средней зоне Волгоградского водохранилища в добыче гнездящихся здесь орланов доля сельдей достигает 85 %. В зимний период ...
... сосуды, которые ничто не способно после этого возвратить к жизни. Норвежские биологи экспериментально доказали, что среди животных, живущих на земле, наиболее морозоустойчивым организмом наделены именно гусеобразные. Эти птицы могут выжить при температуре 110°С ниже нуля! В то же время белые медведи и тюлени сохраняют жизненную активность при температуре до –85°С. Правда, такие морозы в природных ...
0 комментариев