Вступ. 2
1. Походження і розвиток нервових клітин. 3
2. Регенерація нервових волокон. 7
3. Дія центра росту на нервові волокна. 9
4. Регуляція росту нервових волокон. 11
5. Дія периферичних областей на нерви. 16
6. Розвиток функціональних зв'язків. 18
7. Специфічність відносин центра і периферії 20
Висновок. 24
Список літератури. 25
Вступ
Нервова тканина - це високодиференційована спеціалізована тканина, яка формує інтегруючу систему організму — нервову. її структури здатні сприймати подразнення, трансформувати його в нервовий імпульс, швидко його передавати, зберігати інформацію, синтезувати біологічно активні речовини. Завдяки цьому нервова система регулює взаємозв'язок органів і систем організму та адаптацію його до екологічних умов середовища. Нервова тканина складається із двоякого роду органічно зв'язаних клітинних елементів: нервових клітин (нейронів, нейроцитів), здатних утворювати нервове збудження та проводити нервовий імпульс; нейроглії — клітини її забезпечують опорну, трофічну, розмежувальну, секреторну та захисну функції.
У зачатках органів, що розвиваються, існує взаємодія різних тканин, що визначає утворення функціонуючої структури. У цій главі такого роду взаємодії в розвитку анатомічних і функціональних зв'язків будуть розібрані на прикладі взаємозв'язків у нервовій системі, що розвивається, і кінцівці, що розвивається.
Мета роботи полягає в тому, щоб проаналізувати розвиток нервової тканини у безхвостих земноводних та процеси, які цей розвиток супроводжують.
Основні завдання:
1) охарактеризувати походження та розвитку нервових клітин;
2) з`ясувати в чому полягає регенерація нервових волокон;
3) проаналізувати дію центру росту на нервові волокна;
4) розглянути як протікає регуляція росту нервових волокон;
5) охарактеризувати дію периферичних областей на нерви;
6) проаналізувати розвиток функціональних зв`язків та специфічність відношення центру та периферії для нервової тканини.
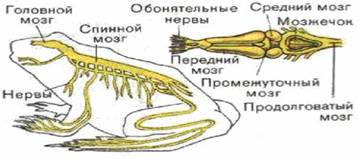
1. Походження і розвиток нервових клітин
Первинні нервові клітки (нейробласти) походять з декількох джерел. Нервова трубка є основним джерелом нейробластів (рис. 1). Вона містить великі округлі нейробласти, що пізніше утворять велику частину нервових кліток і нервові волокна головного і спинного мозку, а також рухові нерви. Крім того, нейробласти виникають із кліток нервового гребеня і черепних плакод. Ці клітини утворять нейрони чуттєвих гангліїв. Деякі нейробласти, що відбуваються з кліток нервового гребеня, мігрують вентрально, утворити вегетативні ганглії. Таким чином, другим джерелом нейробластів служить нервовий гребінь. Нарешті, нейробласти беруть початок з різних плакод: наприклад, нюхові плакоди, розташовані перед мозком, являють собою сенсорні елементи, що утворять нюховий нерв, що вростає в мозок [2].
Розглянемо тепер як розвивається кінцівка, що забезпечується нервами, які виходять з нервової трубки і спинальних гангліїв (рис. 1). Кінцівка, що розвивається, являє собою власне кажучи швидко зростаючу мезодермальну бруньку, у якій диференціюються м'язи і кісти. Мезодерма покрита ектодермой, з якої утворяться зовнішні покриви кінцівки. Задача, отже, полягає в тім, щоб виявити фактори, що забезпечують вростання нервів між гангліями і кінцівкою, між гангліями і спинним мозком, а також між черевним відділом спинного мозку і кінцівкою. Для того щоб кінцівка могла функціонувати, цей нервовий ланцюг, що представляє собою найпростішу форму рефлекторної дуги, повинна замкнутися.
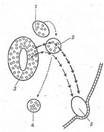
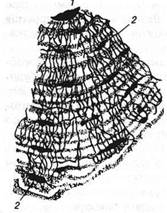
Рис. 1. Зв'язок кінцівки, що розвивається, з нейробластами, що утворюють нерви кінцівки.
Нейробласти зображені у виді маленьких кружків. Пунктирна стрілка вказує шлях міграції нейробластів від нервового гребеня в область утворення спинальних і симпатичних гангліїв. Стрілки, що з'єднують спинальний ганглій зі спинним мозком, позначають шлях нервових волокон, що розвиваються, а стрілки від спинального ганглія до кінцівки показують шлях нервів кінцівки. 1 - нейробласти нервового гребеня; 2 - нейробласти спинального ганглію; 3 - нейробласти спинного мозку; 4 - симпатичні нейробласти; 5 - кінцівка, що розвивається.
Розвиток маленької клітини, діаметр якого виміряється частками міліметра, у нервову клітку з відростком, що відходить від тіла клітини на 60-90 см, можна простежити, спостерігаючи поведінку ізольованих нейробластів у культурі. Невелика ділянка нервової трубки раннього зародка, у якого ще не розвилися нерви, можна помістити в кров'яний згусток у вологій камері (рис. 2). У культурі тканини насамперед спостерігається міграція клітин; при цьому клітини утворюють тонкий шар, що дозволяє спостерігати в мікроскоп за кожною з них. Перше повідомлення про такий дослід було опубліковано в 1907 р. Р. Гарісоном. Цей дослід був проведений для з'ясування питання про те, чи є нервові волокна відростками окремих нейробластів або вони утворюються численними клітками, що складають оболонку волокна, або ж, навпроти, виникають у результаті "кристалізації" заздалегідь сформованих протоплазматичних містків. Досвід Гарісона дозволив існуючий довгий час протиріччя, що стосуються походження нервових волокон, а також стимулював вивчення тканин тварин у культурі. У сучасній біології метод культури тканини є одним із самих перспективних методів дослідження [2].
Спостереження за нейробластами в культурі показують, що спочатку від однієї сторони тіла клітки відходить тонкий тяж цитоплазми (рис. 2). Що нагадує псевдоподію відросток, що зберігає зв'язок з тілом клітки, називають конусом росту. Його переміщення по покривному склу або будь-якому іншому твердому предметові нагадують руху амеби. В міру його видалення від тіла клітки з'єднуючий їх цитоплазматичний тяж подовжується й утворить нервове волокно. Таке волокно може розгалужуватися і мати два конуси росту.
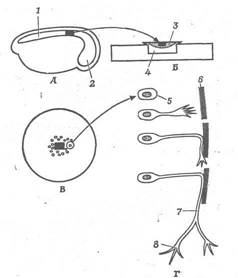
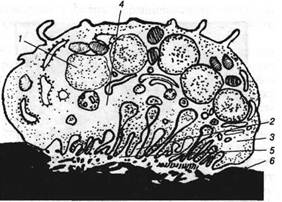
Рис. 2. Походження нервового волокна з одиночного нейробласта. А. Схема операції на зародку жаби. Чорним кольором позначена ділянка нервової трубки. Б. Ділянка нервової трубки в згустку крові, що утримує тканина на покривному склі. Покривне скло щільне закриває поглиблення на предметному склі, так що препарат залишається стерильним і вологим. В. Збільшена схема препарату, на якому видна міграція кліток. М. Поводження однієї з цих кліток; що нагадує псевдоподию виріст рухається по середовищу; стикаючись з перешкодою, виріст повертається і рухається уздовж. цієї перешкоди. Минаючи перешкоду, вона рухається у випадковому напрямку і може розгалузитися, утворити два конуси росту. 1 - нервова трубка; 2 - мозок; 3 - згусток крові; 4 - волога камера; 5 - нейробласт; 6 - перешкода; 7 - нервове волокно; 9 - конус росту.
Подовження нервового волокна не можна розглядати тільки як процес витягування. У тілі клітини утвориться нова цитоплазма, що переміщається дистально по серцевині нервового волокна. Це рух цитоплазми від місця її синтезу уздовж волокна виявляється в досвідах з перетяжкою нервового волокна, що приводить: до нагромадження цитоплазми перед цією перетяжкою (рис. 3).Звичайно розвивається волокно робить звивистий шлях у культуральному середовищі Це свідчить про те, що воно росте не в якомусь визначеному напрямку, а легко змінює його. Конус росту звичайно прагне до переміщення по твердих предметах; цю тенденцію до росту уздовж твердих предметів називають стереотропізмом. Стереотропічні властивості зростаючих нервових волокон виявляються при розвитку зорового нерва (рис. 4). Очний келих, що сформувався, залишається прикріпленим до очного стебельця, що з'єднує його з головним мозком. Очна стеблинка - це не зоровий нерв, а утворення, що з'єднує очний келих з мозком. Зоровий нерв утвориться групою волокон, що відходять від кліток внутрішнього шару очного келиха. Ці волокна ростуть уздовж черевної стінки очної стеблинки, входять у головний мозок і утворюють зоровий нерв.
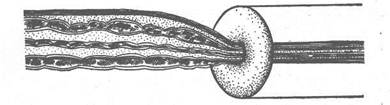
Рис. 3. Результат перетяжки нерва, що регенерує, за допомогою кільця.
Клітинні тіла нерва, що регенерує, знаходяться ліворуч (вони не показані на малюнку); регенерація нервових волокон відбувається праворуч від місця перетяжки. Нервові волокна, проходячи через кільце, що перетягає, стискуються і звужуються. У результаті відбувається нагромадження цитоплазми, що надходить з тіла клітки в нервове волокно. Зверніть увагу на вигин і розбухання волокон ліворуч від місця перетяжки.
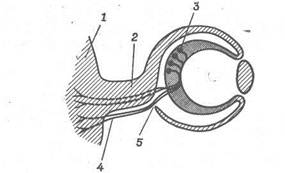
Рис. 4. Очна стеблинка направляє ріст нервових волокон від очного келиха до головного мозку.
Нейробласти, що дають початок зоровому нервові, знаходяться у внутрішньому шарі очного келиха. Конус росту нейробласта переміщається уздовж внутрішньої поверхні очного келиха до отвору - очної щілини, а потім по черевній стороні очної стеблинки назад до головного мозку. 1 - мозок; 2 - очна стеблинка; 3 - нейробласт; 4 - нервове волокно; 5 - очна щілина.
2. Регенерація нервових волоконТенденція нервових волокон рости уздовж твердих предметів має велике значення в процесі регенерації нервів. При перерізанні нерва дегенерує та його частина, що відділена від тіла клітки, а частина, з'єднана з клітиною, регенерує. При регенерації волокно росте, повторюючи хід частини нерва, що дегенерувала (рис. 5).
Якщо допустити, що під час відсутності яких-небудь оформлених структур напрямок росту нервового волокна має випадковий характер, то не дивно, що при наявності вільного простору між волокнами, що регенерують, і колишнім шляхом частини нерва, що дегенерувала, відбувається атипічна регенерація його (рис. 5). Якби цей дефект нерва можна було якимсь чином заповнити, так щоб волокна, що регенерують, могли рости паралельно, то нерв відновив би структуру, дуже близьку до нормального. Одним із таких способів створення умов правильної регенерації може служити пересадження в цей простір ділянки ліофілізованого нерва, багато в чому зберігаючому свою вихідну структуру. Волокна нерва, що регенерує, намагаються рости уздовж мертвих волокон трансплантата, заповнюючи, таким чином, проміжок між перерізаними кінцями нерва. Іншим способом забезпечення росту по шляху нерва, що дегенерував, служить з'єднання двох кінців розрізаного нерва трубкою, що заповнена плазмою крові, обробленої таким чином, що її частки розташовуються паралельно один одному (рис. 5). За допомогою цих і подібних методів можна запобігти безладний ріст нервових волокон; нерв, таким чином, буде регенерувати більш нормально, і повніше відновляться його функції [4].
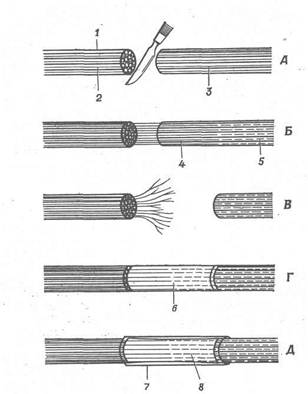
Рис. 5. Регенерація нерва.
A. У перерізаному нерві окремі нервові волокна дегенерують праворуч від розрізу (у дистальній куксі). Ліворуч від розрізу нервові волокна зв'язані з клітинними тілами; їхня дегенерація обмежується лише невеликою ділянкою, що знаходиться безпосередньо біля розрізу; незабаром вони починають регенерувати. Б. Конуси росту, що утворяться на кінці кожного нервового волокна, мігрують уздовж нервових волокон, що дегенерують.
B. Вилучений досить велика ділянка нерва, і волокна, що регенерують, відділені проміжком від частини нерва, що дегенерує. Волокна ростуть у всіх напрямках. Г. У проміжок між кінцями перерізаного нерва поміщають ділянка ліофілізованого нерва, і нервові волокна, що регенерують, ростуть уздовж цього мертвого нерва. Д. Два кінці перерізаного нерва з'єднані трубкою, у якій утримується плазма крові. 1 - нерв; 2 - нервове волокно; 3 - нерв, що дегенерує; 4 - нервове волокно, що регенерує; 5 - слід, залишений волокном, що дегенерувало; 6 - ліофілізований нерв; 7 - трубка; 8 - плазма крові.
3. Дія центра росту на нервові волокнаЗоровий нерв росте по існуючому вже шляху уздовж очної стеблинки і не має потреби в яких-небудь направляючих факторах при проходженні від ока до головного мозку. Однак між нервовою трубкою і кінцівкою такого шляху, що розвивається, немає. На цій ділянці розташовуються скупчення мезенхіми і деякі соміти, але немає нічого, що направляло б нерв до кінцівки. Таким чином, або напрямок росту рухових і чуттєвих нервів зі спинного мозку визначено, так що вони попадають у кінцівку, або кінцівка якимсь образом притягує до себе нерви. Досвіди з пересадженням зачатка кінцівки дозволили зробити вибір між цими двома альтернативами [3].
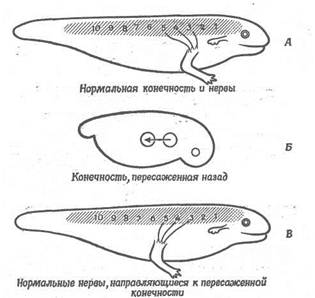
Рис. 6. Направляюча дія кінцівки на ріст 3-, 4- і 5-го спинальних нервів.
А. 3-, 4- і 5-й нерви нормально ростуть у напрямку до кінцівки. 5. Передню кінцівку пересаджують небагато назад. В. 3-, 4- і 5-й нерви повертають назад у напрямку пересадженої кінцівки й іннервують неї.
Як показано на рис. 6, передня кінцівка амфібій у нормі інервується нервами, що йдуть з 3-5-го сегментів спинного мозку. Якщо до початку росту нервів зачаток кінцівки пересадити назад, помістивши його проти 5-7-го сегментів, то нерви, що йдуть від 3-5-го сегментів, відхиляються від свого звичайного шляху, повертають назад і іннервують пересаджену кінцівку (рис. 6). Якщо зачаток кінцівки пересадити ще далі назад, проти 6- 8-го сегментів, то кінцівка інервується нервами, що йдуть з 6-8-го сегментів (рис. 7). Отже, існує межа для відстані, на яке можуть відхилитися нерви нормальної кінцівки, щоб урослися в пересаджену кінцівку.
Виникає питання, чи специфічно притягання нервів зачатком кінцівки, тобто чи обумовлено воно саме кінцівкою або ж подібна дія робить будь-як зростаючий орган. Для рішення цього питання зачаток кінцівки був пересаджений назад, як у раніше описаному досвіді, а нюхова плакода пересаджена на місце кінцівки. У цьому випадку нерви кінцівки росли в напрямку до нюхового плакоде, як якби вона була кінцівкою. Існує, очевидно, загальний стимул, загальне притягання нервів зростаючими органами. І не має особливого значення, який орган виявляє це притягання: кінцівка або нюхова плакода. 6-й,7-й a 8-й нерви постачають пересаджену кінцівку
-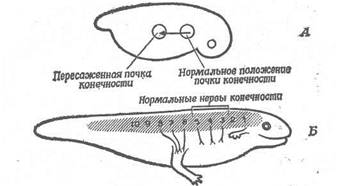
Рис. 7. Пересаджена кінцівка може направляти ріст 3-, 4- і 5-го нервів, якщо неї пересадити далеко назад від місця утворення цих нервів (А). У зазначеному положенні кінцівка інервується 6-, 7- і 8-м спінальними нервами (Б).
Потенційно нерви можуть рости в напрямку до структур, що відрізняється від структур, що вони в нормі іннервують. Цікаво, що хоча нерви і притягаються чужорідними структурами, але не завжди проникають у них. Очевидно, мається визначена специфічність, що обумовлює проникнення нервових волокон у тканину, - специфічність, що не залежить від сил притягання і проявляється не дуже різко. Так, нерви передньої кінцівки не проникають у пересаджену на її місце нюхову плакоду, але кінцівка, пересаджена на місце ока, одержує гілочки від V і VІІ черепно-мозкових нервів. Однак у цьому випадку характер іннервації лише наближається до характеру іннервації кінцівки в нормі.
4. Регуляція росту нервових волоконДля пояснення притягання нервів кінцівкою, що розвивається, були запропоновані різні теорії. Було висловлене припущення, що від кінцівки дифузно поширюється якась специфічна речовина і що конус росту орієнтується в напрямку збільшення концентрації цієї речовини (рис. 8, А). Це явище, називане хемотаксисом, відіграє визначену роль у процесах агрегації в миксоміцетів, однак даних на користь його участі в системі нерв - кінцівка недостатньо. Більш того, переконливо показано, що ріст нервів визначається не хемотаксисом. Ріст волокон, що регенерують, у напрямку до частини нерва, що дегенерувала, можна порівняти з ростом нервових волокон у напрямку до кінцівки; на підставі цього було висловлене припущення, що завдяки хемотаксисові волокна, що регенерують, ростуть назустріч речовинам, виділюваним частиною нерва, що дегенерувала. Однак в експериментальній системі, що дозволяє волокнам, що регенерують, рости по двох каналах, з яких тільки в одному знаходиться ділянка нерва, що дегенерує, волокна ростуть по обох каналах, не виявляючи хемотаксису [6].
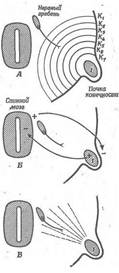
Рис. 8. Схема, що ілюструє три теорії, що пояснюють направляючий вплив почки кінцівки на ріст нервів кінцівки.
A. Деяка речовина (1), що утримується в кінцівці, дифундує через тканини до нервового гребеня. У процесі диференціювання нейробласта його конус росту попадає у зону слабкої концентрації цієї речовини, K1. Воно діє на мігруючий конус таким чином, що він завжди спрямований убік найвищої концентрації. У результаті конус росту мігрує до місця найвищої концентрації цієї речовини, Кт- Б. Між кінцівкою і нервовою трубкою встановлюється різниця потенціалів. Конус росту мігрує уздовж напрямку, у якому проходить струм.
B. Кінцівка, яка росте впливає на довгі вузькі частки, розташовані в середовищі між нею і нервовим гребенем. Конус росту мігрує уздовж орієнтованих часток, як це відбувається й у культурі тканини. Перші дві теорії (А и Б) не підтверджуються якими-небудь експериментальними даними.
Інша гіпотеза заснована на принципі гальванотаксиса, або орієнтації кліток відповідно до напрямку електричного струму, і виходить із представлення, що зростаюча структура заряджена негативно стосовно інших тканин організму (рис. 8, Б). При цьому допускається, що нерв, що розвивається, росте уздовж силових ліній електричного поля, що існує між спинним мозком і кінцівкою. Подібний механізм становлення іннервації кінцівки не підтверджений ніякими конкретними даними [7].
Концепція "контактної спрямованості" (contact guіdance) стверджує, що нервове волокно росте назустріч розвивається кінцівки під впливом механічних стимулів. Відповідно до цієї концепції, колоїдні частки в рідинах тіла зародка розташовані упорядковано; їх довгі осі орієнтовані в одному напрямку, і в цьому ж напрямку мігрує конус росту. Ця теорія заснована на результатах дослідів із тканинними культурами, у яких порівнювали ріст нервових волокон у середовищі з неорієнтованими й у середовищі з орієнтованими частками. У неорієнтованому середовищі нервові волокна ростуть з експлантата у всіх напрямках (рис. 9, А), а в орієнтованої - лише в напрямку орієнтації (рис. 9, Б). Якщо на одне покривне скло помістити дві культури нейробластів, то можна бачити, що під впливом їхнього росту середовище між ними стає орієнтованої, так що нервові волокна з обох культур ростуть в одному напрямку назустріч один одному (рис. 9, В). У концепції "контактної спрямованості" допускається, що зростаюча брунька кінцівки (або інший орган) викликає розтягання середовища між кінцівкою і спинним мозком і, таким чином, орієнтує вузькі довгі колоїдні частки в рівнобіжні ряди. Як тільки це відбувається, виникають свого роду "направляючі лінії", уздовж яких конус росту зустрічає менший опір, і нерви, що розвиваються, починають рости в цьому напрямку. Орієнтацією подібного роду можна також пояснити утворення нервового сплетення в підставі кінцівки.
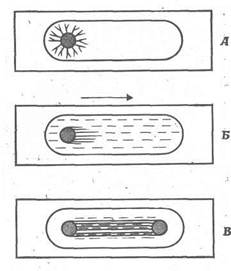
Рис. 9. Орієнтація довгих вузьких часток у середовищі і її вплив на ріст нервових волокон.
А. Культура нейробластів у неорієнтованому середовищі. Нервові волокна ростуть у всіх напрямках. Б. Середовище орієнтоване шляхом розтягування, після чого нервові волокна ростуть паралельно один одному. В. Дві культури орієнтують середовище, що знаходиться між ними, і нервові волокна ростуть паралельно один одному.
Концепція "контактної спрямованості" зустрічає менше заперечень, чим дві інші гіпотези про причини росту нервових волокон у напрямку органів. Однак вона, очевидно, не дає повного пояснення спрямованого росту нервових волокон. Орієнтований субстрат може забезпечити загальний напрямок росту волокон, але навряд чи цим можна пояснити численні випадки дуже точної орієнтації волокон і встановлення специфічних функціональних зв'язків. Цілком можливо, що очна стеблинка служить механічним субстратом, що забезпечує вростання нервів у головний мозок, а субклітинної орієнтацією субстрату цілком можна пояснити ріст нервів у напрямку до кінцівки, однак подальший ріст цих нервів у мозку і кінцівці припускає існування інших, хоча, бути може, і подібних, механізмів. Мігруючі пігментні клітини, що беруть початок від нервового гребеня, можуть переміщатися уздовж ліній орієнтації субстрату, але цим не можна пояснити виникнення упорядкованих візерунків пігментації. У культурі ці клітки розташовуються на визначених відстанях друг від інша імовірно, таке їхнє розташування в культурі, а також міграція від нервового гребеня по бічних сторонах тіла тварини порозуміваються взаємним відштовхуванням кліток. Таким чином, навіть незважаючи на те, що "контактна спрямованість", можливо, визначає основні шляхи переміщення кліток до місця їхнього призначення, концентрація продуктів обміну речовин у цих клітках може служити стимулом для їхньої міграції і забезпечувати остаточний розподіл і положення кліток [6].
Заслуговують також згадування дані про існування іншого роду специфічності. Звичайно, як тільки нерви досягають кінцівки, яка розвивається, вони розподіляються в ній нормальним образом. Примітно, що якщо нерви доростають до добре розвитий передньої кінцівки, цілком позбавленої нервів, то в ній виникає той же тип іннервації, що й у молодій кінцівці, що розвивається; отже, нерви, потрапивши в кінцівку, що знаходиться на ранній або пізній стадії розвитку, розгалужуються однаково. Тенденцію нервових волокон рости уздовж вже існуючих кровоносних судин можна пояснити тим, що це шлях найменшого опору. Остаточно характер іннервації кінцівки, очевидно, встановлюється в результаті взаємодії рано розвиваються м'язів з рано розвиваються нервовими волокнами. Ця можливість найкраще виявляється при розгляді питання про вплив кінцівки на нервову систему і про специфічність нейронів.
5. Дія периферичних областей на нервиДуже сильний вплив на нервову систему, що розвивається, роблять структури, з якими в неї установлюється функціональний зв'язок. Це знаходить своє відображення в розмірах нервів і гангліїв. Нерви кінцівок, що містять як рухові, так і чуттєві волокна, крупніше інших спинальних нервів; ганглії нервів кінцівок крупніше, ніж інші спинальні ганглії. Видаляючи кінцівку на одній стороні зародка і залишаючи кінцівку протилежної сторони недоторканої, можна показати, що велика величина нервів і гангліїв безпосередньо зв'язана з периферичним функціональним навантаженням на нерви.
На ранніх стадіях розвитку вентролатеральна частина спинного мозку диференціюється в рухову область, і волокна з нейробластів цієї області іннервують м'яза тіла і кінцівок. Спинальні ганглії містять клітинні тіла чуттєвих нервів, і рефлекторна дуга замикається через спинний мозок. Очевидно, що між кількістю інервованої тканини і диференціацією клітин у нервовій системі існує прямий зв'язок. Цей висновок підтверджується результатами досліду з пересадженням кінцівки в нове положення. У такому досліді пересаджена кінцівка інервується новими нервовими відростками. У результаті кількість волокон у цих нервах збільшується і наближається до нормального для волокон рухового нерва [5].
У більшості хребетних кінцівка індукує утворення різного числа рухових нейронів у спинному мозку і чуттєвих нейронах у спинальних гангліях. Установлено кількісні співвідношення між периферичним навантаженням і диференціюванням нейронів в амфібії.
Кінцівка, що розвивається, створює комплексну дію на спинний мозок і ганглії. Присутність кінцівки, очевидно, викликає посилену проліферацію кліток, а також збереження кліток, що звичайно дегенерують. Відомо, що загибель клітин і нормальне явище в розвитку різних систем, у тому числі і центральної нервової системи. Присутність власної або пересадженої кінцівки запобігає звичайній дегенерації деяких нейронів у центральній нервовій системі. Відсутність кінцівки або зменшення периферичної області кінцівки приводить до збільшення кількості клітин, що дегенерують. Ці фактори при незалежній або спільній дії можуть забезпечити відповідність величини нервових, центрів їхнім периферичним зв'язкам.
Вплив периферичних областей на диференціювання клітин нервових центрах, цілком ймовірно, здійснюється через нервове волокно. При видаленні кінцівки, що розвивається, від задньої кінцівки від нейробластів у спинному мозку, як звичайно, відходять цитоплазматичні волокна. Вони ростуть у напрямку до культи і розташовуються в тканині рубця безладним чином. Протягом декількох днів ці волокна, що не прикріпилися, атрофуються, а їхні клітинні тіла у вентральній частині спинного мозку дегенерують. Очевидно, для нормального розвитку нейрона необхідно, щоб аксон нерва, що розвивається, установив контакт із визначеною структурою. Областю контакту, видимо, є конус росту волокна; якщо контакт не установився, те відповідна інформація якимсь образом передається тілу клітки, центрам її метаболічної і синтетичної активності, у результаті чого припиняється диференціювання або відбувається дегенерація. Отже, установлення периферичних зв'язків досить істотно для підтримки нервових кліток, що розвиваються, у визначеному стані. Збереження і виживання нервових кліток, можливо, залежить також і від їхніх взаємин один з одним. Процес дегенерації може переходити з одного нейрона через аксон і синапс на інший нейрон, що спричиняє дегенерацію цього другого нейрона; таке явище називають міжнейронною дегенерацією; його, зокрема, можна спостерігати в слуховій і зорової системах. Наприклад, навіть саме незначне ушкодження сітківки приводить до помітної атрофії зорових часток середнього мозку шляхом міжнейронної дегенерації.
На диференціюванні клітин у нервових центрах, крім зовнішніх факторів, що діють через нервові волокна, впливають також гуморальні фактори. Дослідження іn vіtro показали, що фактор росту скоріше діє на тіло клітини, а не на аксон. Дією саме на тіло клітки порозумівається, очевидно, посилене розвиток нервових клітин під впливом тиреоїдного гормону. У процесі нормального розвитку гуморальні агенти (гормони) по кровоносних судинах можуть попадати в різні частини нервової системи і безпосередньо впливати на що розвиваються нейробласти [4].
Зв'язок нервових центрів, що розвиваються, з периферичними областями залишається багато в чому нез`ясованим. Та обставина, що зовнішні фактори впливають на диференціювання нейронів, яким би не був механізм цього впливу, можливо, ще раз підкреслює важливу роль, що відіграють різні форми індукційних взаємодій у процесі прогресивного диференціювання систем, що розвиваються.
6. Розвиток функціональних зв'язків
Спинний мозок зародка характеризується лабільною організацією. В амфібій він організований таким чином, що 3-5-і сегменти іннервують передні кінцівки, а 6-8-і сегменти - м'яза, розташовані за цими кінцівками. Ці дві ділянки на ранніх стадіях розвитку можна поміняти місцями, і вони замінять один одного. Як показано на рис. 10, у зародка можна видалити3-5-і сегменти і замінити їхній 6-8-ми сегментами, узятими в іншого зародка. При цьому функція передньої кінцівки цілком зберігається, а це означає, що на ранніх стадіях розвитку 6- 8-і сегменти лабільні і їх можна змусити функціонувати по типу3-5-х сегментів.
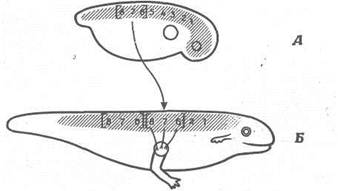
Рис. 10. Заміна 6-, 7- і 8-го сегментів спинного мозку 3-, 4- і 5-м сегментами. Зачаток передньої кінцівки залишається в колишнім положенні. Кінцівка, що розвивається, притягає нерпи з пересаджених 6-, 7- і 8-го сегментів і нормально функціонує. Отже, нове середовище викликало розвиток 6-, 7- і 8-го сегментів по типу 3-, 4- і 5-го сегментів. А. Пересадження 6- 7- і 8-го сегментів на місце 3-, 4- і 5-го. Б. Пересаджені сегменти нормально функціонують.
Принцип лабільної організації прикладемо також до 3-5-му нервів. Додаткова брунька кінцівки, трансплантована поруч з нормальної, часто інервується відгалуженням 5-го нерва. У цьому випадку додаткова кінцівка функціонує нормально; якщо ж вона інервується 6-8-ми нервами без відгалуження від 5-го нерва, то функціонувати нормально вона не може. У той же час однієї гілочки кожного з 3-5-го нервів досить для нормального функціонування передньої кінцівки. Таким чином, на ранніх стадіях розвитку всі розходження в організації цих трьох нервів носять лабільний характер. Нагадаємо, що спінальні нерви утворять сплетення в основі кінцівки, так що сегменти, що іннервують кінцівку, мають тенденцію функціонувати як єдина група [5].
Лабільність нервів виявляється в явищі гомологічної функції. Якщо м'яз кінцівки личинки амфібій, пересаджений поруч з нормальною кінцівкою, інервується відгалуженням від нерва кінцівки (наприклад, відгалуженням від 5-го спінального нерва), то пересаджений м'яз завжди буде скорочуватися одночасно з таким же м'язом у нормальній кінцівці. Якщо пересадити в такий же спосіб цілу кінцівка, то кожен м'яз останньої буде скорочуватися одночасно з її гомологом у нормальній кінцівці і руху додаткової і нормальної кінцівок будуть синхронними. Ще більш дивно, що якщо додаткову кінцівку пересадити так, що її полярність стає протилежної полярності нормальної кінцівки, розташованої на тій же стороні, те їхнього руху будуть дзеркально відбивати один одного. Отже, нерви, що постачають гомологічні м'яза, тобто м'яза, що володіють тотожною конституціональною специфічністю, стають функціонально зв'язаними, хоча такі функціональні ефекти можуть виявитися марними або навіть шкідливими для тварини. Описане явище гомології функцій показує, що м'яз реагує на особливий вид стимулу незалежно від специфічності її нерва, що іннервує, а це означає, що м'яз, або периферична область, відіграє важливу роль у встановленні специфічних функціональних зв'язків у нервовій системі.
6. Специфічність відносин центра і периферіїЧисленні дані показують, що в ході диференціювання нейрони набувають визначеної біохімічної специфічності, що складає основу складних функціональних взаємозв'язків між нервовими центрами і периферичними областями. Ця проблема була найбільше повно вивчена на прикладі зорової системи хвостатих амфібій. Як у дорослих, так і в зародків цих тварин можна вирізувати ока, повернути їх і знову посадити на місце; при цьому зоровий нерв відновлюється. Однак одержувані результати різко розрізняються в залежності від того, на якій стадії була проведена операція. Так, до стадії пізньої хвостової бруньки око можна повернути на 180° без яких-небудь серйозних порушень зору або очноорухливої координації. Пізніше, однак, така ж операція різко порушує очних реакцій тварини [1].
Справедливість представлення про специфічність зв'язків між чуттєвим центром і периферичною областю підтверджується також даними, отриманими для систем, що відрізняються від зорової, котра, являє собою центральну систему, оскільки сітківка утвориться з частини нервової пластинки. Існування специфічного зв'язку між нервом і периферичною областю було також показане в досвідах зі шкірними нервами. Якщо в пуголовка жаби вирізувати дорзовентральну смужку шкіри, повернути неї на 180° і знову посадити на місце, вона приживляється і буде самодиференціюватися, так що після метаморфоза на спині тварини буде шматок світлої шкіри черева, а на череві - шматок темної шкіри спини. Якщо подразнювати ділянку темної шкіри на животі, то тварина намагається змахнути подразник зі спини або усунути подразник з живота, якщо подразнювати ділянку світлої шкіри на спині.
Дані, отримані в таких дослідах, пояснити не дуже легко, і будь-яке їхнє пояснення в даний момент буде чисто умоглядним. Однак пропоноване нижче пояснення співпадає з відомими фактами і показує, які процеси можуть брати участь у встановленні численних і різноманітних зв'язків у нервовій системі (рис. 11). Цілком імовірно, що тип центральних синаптичних зв'язків, утворених нервовими волокнами, визначається, принаймні частково, властивостями периферичної області, з яким зв'язані ці волокна. Таким чином, можна представити, що, як тільки конус росту волокна, що розвивається, досягає шкіри, в області контакту між нервом і шкірою відбувається індукційна взаємодія, за допомогою якого шкіра додає нервові особливу біохімічну специфічність, що визначає виникнення центральних зв'язків шляхом більш тонких індукційних впливів на рівні нейронів. Якщо розглянути з цих позицій зорову систему, то виявиться, що на визначеній стадії розвитку сітківка стає поляризованої і на більш пізніх стадіях зв'язок між будь-якою частиною сітківки і зоровою часткою закріплюється. Таким чином, можливо, що в процесі розвитку сітківка диференціюється до такого стану, коли кожна сама маленька ділянка її володіє особливою біохімічною специфічністю. Досягши зорової частки, волокна специфічних кліток наділяють відповідною специфічністю центральні нейрони. Отже, аксони, що проникають у відповідну частину зорової частки, можуть сприяти виникненню специфічності в тканині центральної нервової системи, а після цього зв'язок між сітківкою і зоровою часткою жорстко фіксується, подібно зв'язку між шкірою і нервом. Тут важливо нагадати, що порушення очнорухливих реакцій після повороту ока, що свідчить про втрату лабільності системи "сітківка - зорова частка", виникає в той час, коли перші волокна зорового нерва досягають зорової частки. У процесі розвитку відбувається просторове диференціювання нервової системи.
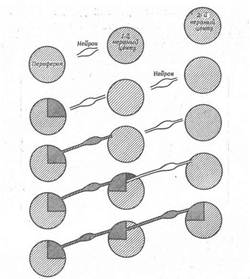
Рис. 11. Схема послідовних процесів, що приводять до виникнення специфічності нейронів і нервових центрів.
Специфічність, очевидно, уперше виникає в периферичній області. Потім зростаючі нейрони, установлюючи функціональні зв'язки з периферичною областю, також здобувають відповідну специфічність. Після цього вони передають свою специфічність нервовим центрам. Таким шляхом різні нейрони і нервові центри здобувають у ході розвитку взаємозалежну анатомічну і функціональну специфічність.
Нейрони з різних частин головного мозку часто розрізняються за формою і розміром. У мозку відбувається диференціальний розподіл холинэстеразы і катехоламинов, а тканина центральної нервової системи істотно відрізняється від периферичної по антигенному складі [8].
Становлення особливої біохімічної специфічності зв'язків між центральною і периферичною областями не означає, що внутрішня організація частин нервової системи цілком залежить від тонких індукційних впливів, що торкаються специфічність нейронів; воно не виключає необхідності направляти ріст волокон і не є незалежним від таких факторів, як тимчасова послідовність розвитку і відповідні просторові зв'язки.
Висновок
Підводячи підсумки, можна сказати, що загальні принципи розвитку організму на ранніх стадіях застосовні також до розвитку нервової системи.
Як для центральної нервової системи, так і для периферичних нервів характерні вихідна лабільність і взаємодія, що детермінує, з периферичними областями.
Розвиток специфічної функції, що починається з найпростішої форми організації і перехідне до самої витонченої взаємодії між нейронами в соматосенсорної області кори головного мозку, характерно для нервової системи. Відповідно м'язові рухи і пристосування тварини до середовища розвиваються від загальних до більш специфічного. Індукційні й інші взаємодії кліток і вироблення специфічних біохімічних властивостей у нервовій системі й у зв'язаних з нею структурах уражають своєю складністю і для їхнього остаточного аналізу буде потрібно застосування безлічі різноманітних методів.
Список літератури
1. Александровская О.В., Радостна Т.П., Козлов НА. Цитология, гистология и эмбриология. - М.: ВО «Агропромиздат», 1987 - 448 с.
2. Белоусов Л. В. Введение в общую эмбриологию: Изд-во Московского университета, 1980. — 210 с.
3. Бодемер Ч. Современная эмбриология. — М.: «Мир», 1971. — 446 с.
4. Вельш У., Шторх Ф. Введение в цитологию и гистологию животных. — М.: «Мир», 1972. — 259 с.
5. Иванов И.Ф., Ковальский П.А. Цитология, гистология, эмбриология. М.: «Колос», 1976. — 447 с.
6. Международная гистологическая номенклатура / Под ред. Ю.И.Копаева — К., 1980. - 113 с.
7. Свенсон К., Уэбстер П. Клетка. — М.: «Мир», 1980. — 303 с.
8. Токин Б.П. Общая эмбриология. М.: «Высшая школа», 1977. — 509 с.
9. Хэм А., Кормак Д. Гистология. — Т. 1—5. — М.: «Мир», 1983. — 272 с.
10. Ченцов Ю.С. Общая цитология: Изд-во МГУ, 1984. — 350 с.
11. Шмидт Г.А. Эмбриология животных. Ч.1: Общая эмбриология: Государственное изд-во «Советская наука». — М., 1951. — 354 с.
Похожие работы
... проривні пучки і канали з кровоносними судинами та нервами, розгалужуються, з'єднуються з центральними каналами остеонів і анастомозують між собою. Остеоцити лежать між кістковими пластинками, а кісткові канальці пронизують системи пластинок. Розвиток кісткової тканини — остеогенез. Існує два різновиди остеогенезу — безпосередньо із мезенхіми та заміни кісткою гіалінового хряща. Ділянки мезенх ...

... Розселення трав'яних жаб по суші визначається їх залежністю від вологості. У цьому відношенні вони займають проміжне положення між зеленими жабами і ропухами. Вони здатні без шкоди для себе втрачати більшу кількість води, ніж ставкові жаби, але значно менше, ніж ропухи, особливо зелені. У північних широтах свого існування в зв'язку з низькими температурами повітря і на півдні, де велика сухість ...
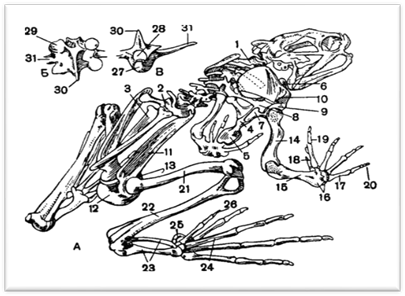
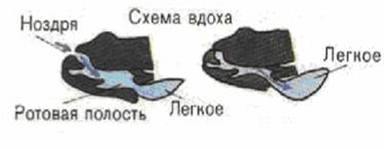
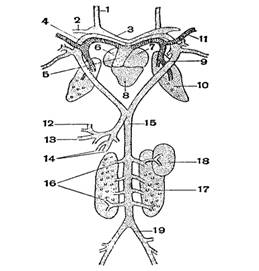
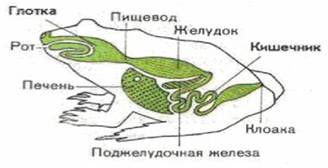
... хвостатих земноводних зябра зберігаються на все життя. [3] 2.3 Кровоносні органи З розвитком наземного органу дихання тісно пов'язана і перебудова системи кровообігу. Серце земноводних складається з двох цілком відокремлених передсердь, загального шлуночка і артеріального конуса, від якого відходить загальний стовбур аорти, що розділяє потім на три пари артеріальних судин. Передні з них - ...
лярною пластинкою решітчастої кістки. Вільний задній край леміша розмежовує хоани. Вилична кістка (os zygomaticum) формує рельєф обличчя. З нею сполучаються одноіменні відростки скроневої, лобної та верхньощелепної кісток. Скроневий відросток виличної кістки та виличний відросток скроневої кістки утворюють виличну дугу. Нижньощелепна кістка (mandibula) єдина рухлива кістка черепа. Склада ...
0 комментариев