Введение
Общие представления о нейропептидах.
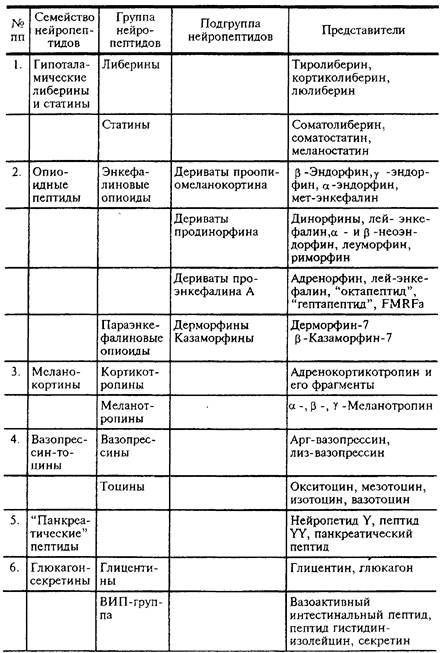
Классификация
Первое семейство
Второе семейство - опиаты.
Третьего семейства НП
Четвертое семейство НП
Пятое семейство
Шестое семейство
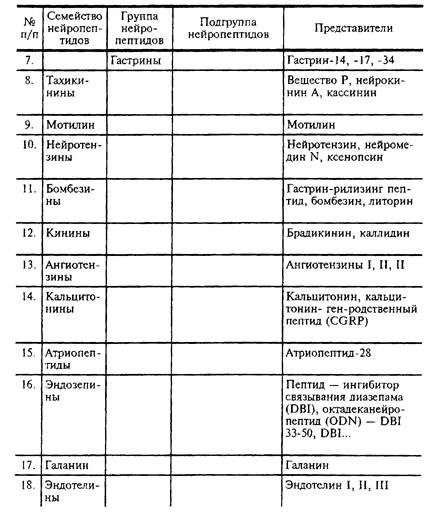
Седьмое семейство
Восьмое семейство
Десятое семейство
Одиннадцатое семейство
Двенадцатое и тринадцатое семейства
Четырнадцатого семейство
Пятнадцатое семейство
Шестнадцатое семейство
Пептиды - коннекторы
Процессинг и посттрансляционная модификация нейропептидов.
Метаболизм энкефалинов и ферменты их процессинга.
Краткие характеристики карбоксипептидазо-В-подобных ферментов.
Сarboxypeptidase U (M14.009)
M14.004: carboxypeptidase N
S10.005: serine carboxypeptidase D
M14.005: carboxypeptidase E
M14.006: carboxypeptidase M
PMSP-ингибируемая карбоксипептидаза
Заключение.
Литература
Введение
“Я не видел еще такой проблемы,
какой бы сложной она ни была,
которая при верном взгляде на нее
не становилась бы еще более сложной”.
Поль Элдестон
Вступление современной биохимии и физиологии человека в двадцать первый век ознаменовалось бурным развитием сравнительно нового биологического направления – нейрохимии. Этому способствовали, прежде всего, современный уровень развития биохимических исследований, как в теоретическом, так и в методическом отношении. На интенсивное изучение нейрохимии огромное влияние оказали такие факторы, как широкое применение разнообразных психофармакологических, наркотических и других веществ, а также распространение нейрологических заболеваний, в основе которых лежат глубокие нарушения биохимических процессов, протекающих в нервной ткани. Достижения нейрохимии на сегодняшний день могут служить показателем уровня наших знаний о мозге, его структуре и функциях его компонентов. Кроме того, в связи с резким увеличением потока информации и повышением интеллектуальной деятельности человека в настоящее время приобретают значение исследования, посвященные изучению основ памяти, обучения, оптимальных биохимических условий функционирования головного мозга.
Однако не стоит наивно полагать, что с помощью изучения только молекулярных и биохимических механизмов, можно разрешить все загадки живой природы. Живые организмы это набор интегрированных структурных элементов, и было бы ошибочно утверждать, что на основе изучения функциональности, скажем, одного нейрона, можно делать выводы о функциональности всего мозга. К сожалению, некоторые недобросовестные научные работники часто пытаются строить теории, опираясь лишь на результаты биохимического скрининга, не утруждая себя изучением взаимодействия микросистем на тканевом и органном уровнях.
Тем не менее, современная нейрохимия тесно связана с физиологией, гистологией, нейробиологией, биохимией и многими другими биологическими дисциплинами и совместно с ними разрабатывает направления исследования фундаментальных и прикладных вопросов.
Нервная система представляет собой исключительно сложную, гетерогенную и при этом уникальную биологическую систему, как в структурно-морфологическом, так и в функциональном отношении. Одной из важнейших функций центральной нервной системы является её регулирующая и интегрирующая роль по отношению к биологическим процессам, происходящим в целом организме человека и животных. Этим и определяются специфические особенности состава и метаболизма, происходящего в нервной такни, а также наличие в нервной системе сложных компенсаторных и регуляторных механизмов.
При изучении функциональной специфичности различных отделов головного мозга представляет исключительный интерес состав и структура пластических веществ. Функциональная же специфичность зависит от особенностей пластических веществ, которые образуют структуры, определяющие деятельность различных отделов мозга. В связи с этим немаловажный интерес представляют белки, пептиды и ферменты регуляции их процессинга и посттрансляционной модификации.
Общие представления о нейропептидах
Речь идет о химических соединениях пептидной природы, выполняющих в организме роль регуляторов разнообразных физиологических функций. Они логично и последовательно связаны между собой.
Первая — организация коммуникации между разными клетками посредством специализированного химического импульса.
Вторая — обеспечение “настроя” клетки, которая реагирует на воздействия того или иного рода. Это так называемая модуляция функции нервной или другой клетки организма.
Третья — участие в реализации отдельной физиологической реакции или сложного акта.
Сегодня мы можем говорить о классе универсальных химических регуляторов, значимость которых простирается от влияния на функции отдельных групп клеток до управления работой целых систем и органов, включая сложные акты поведения.
Попытаемся показать причины постулируемой “универсальности” нейропептидов, которые находятся сегодня в центре внимания широкого круга специалистов.
В природе существуют такие структуры, которые оказались на редкость удачными в организации систем любой сложности. Одна из них — аминокислота. Это минимально сложное органическое соединение, одновременно и кислота, и основание, потому что в него с двух концов вмонтированы амидная и карбоксильная группы. Они помогают аминокислотам соединяться друг с другом, образуя относительно прочные и в то же время лабильные структуры. Известно около 150 аминокислот. Живая природа использует только 20 из них. Однако представьте, какое количество комбинаций можно сделать лишь из 20 исходных единиц! Из них созданы все белки, которые составляют основу любого организма — структурные, каталитические (ферменты), регуляторные. В результате серии последовательных химических реакций, осуществляемых с помощью специальных ферментов (пептидаз), в клетках образуются олигопептиды, которые обладают высокой биологической активностью и которые были классифицированы как регуляторы разнообразных физиологических процессов.
Таких физиологически значимых пептидов было открыто несколько сотен. Но основной “костяк” — не более 40—50, остальные — их комбинации, дополнения. Как правило, регуляторные пептиды — это молекулы с различным набором аминокислот: большинство из них — до 30, больше не надо. Есть какой-то энтропийный уровень, уровень упорядоченности систем, оптимальный для выполнения регуляторной миссии. Однако все более углубленное исследование соотношения структуры и функции показывает, что части целой пептидной молекулы, ее фрагменты, также могут обладать физиологической активностью, подчас еще большей или качественно инвертированной.
Существует сложный биохимический процесс биосинтеза пептидов в клетке: экспрессия соответствующих генов, образование mRNA , считывание, как с конвейера, ленты аминокислотных “кирпичиков”, образующих белковую молекулу, далее разрезание этих белков на блоки определенной структуры.
Все это — точно прослеженные биохимические процессы, изученные для большинства физиологически значимых пептидов. Более того, тонкая генно-биохимическая “кухня” работает в строгой ПРИУРОЧЕННОСТИ к функции органа и к определенному биоритму его деятельности. Вот здесь — в “привязке” к работе органа, в соответствии его функциональной “ритмике” — главное содержание регуляторной роли нейропептидов.
Понятие “нейропептиды” тогда, когда в мозге открыли вещества, влияющие на центральные функции высших организмов. Они были названы эндорфинами и энкефалинами и дали начало изучению большой и значимой группы опиоидов, список которых пополняется и поныне. Нейропептиды оказались как бы над группами других, “периферических” пептидов, регулирующих работу сердца, почек, кишечника. Выяснилась также причастность к работе мозга ранее известных пептидных гормонов и их фрагментов — АКТГ, соматостатина, окситоцина. Со временем в мозге открывались новые и новые нейропептиды, влияющие на все многообразие его функций.
Стало давним штампом сравнение мозга с компьютером. Оно базируется на внешней оценке работы нервной клетки — приеме, переработке и передаче информации, а также на анатомическом подобии гигантской нейронной сети мозга, которую напоминали первые компьютерные агрегаты. Считается, что 1.5 кг мозга человека содержат около 100 млрд нервных клеток, а в 1 мм3 нервной ткани вмещается более 40 тыс. нейронов! Однако вся эта “коммуникационная анатомия” тысячекратно усиливается, если принять во внимание, что каждый нейрон имеет не менее 100—1000 синапсов. Поэтому вероятный запас коммуникационных возможностей мозга на два-три порядка больше. Значит, современный человек может оценивать собственный мозг как гигантский компьютерный зал, целое “государство” компьютеров высокой надежности, разнообразного класса и назначения. Но жизнь этого “государства” и работа его подразделений (зон, ядер мозга), даже отдельных нервных клеток нуждается в постоянном управлении — поддержании рабочего тонуса и его изменении в зависимости от меняющихся потребностей организма.
Исключительная насыщенность нервной ткани дает повод пойти по ложному следу. Срабатывает стереотип: это “много” связать с разнообразной работой мозга; перенести КОЛИЧЕСТВЕННУЮ сложность анатомической структуры на объяснение полифонии выполняемых функций.Стереотип связывался с возможностью синтеза специфических пептидов, ответственных за память, сон, страх, агрессивность и т.п. Впервые это случилось в 60-х годах, в пору интенсивного развития нейрокибернетики, когда была популярна идея, что специфичность получаемой мозгом информации определяется синтезом определенных химических молекул. Множественность функций, присущих мозгу, пытались ассоциировать с его возможностью синтезировать великое и разнообразное множество пептидов.
Эта посылка получила развитие в теории Г.Унгара, в 1965 г. сформулировавшего принцип: “Один пептид — одно поведение”. На первом этапе автор гипотезы будто бы получил весомые доказательства. Опыты с переносом закрепленного навыка от “обученных” крыс “необученным” посредством инъекции смеси пептидов произвели сенсацию. Более того, Унгар описал структуру пептидов, названных скотофобином и амелитином. В дальнейшем активный поиск веществ, ответственных за проявления страха, тревоги, устойчивости к стрессу, и радости оказался привлекательным... Однако в перечне пептидов, открытых и исследованных в последние 40 лет, нет веществ, “описанных” Унгаром. О скотофобине и амелитине к настоящему времени, по сути, забыто.
Пептиды синтезируются в нервных и железистых клетках, а также в эндотелии — особой моноклеточной ткани, выстилающей кровеносное русло. Кроме того, пептиды образуются в крови или выбрасываются в кровоток различными тканями. Вот почему исходное понятие “нейропептиды” оказывается как бы размытым: они (по крайней мере основная часть) синтезируются во всех точках организма и “работают” также везде — в мозге, в почках, в легких, в репродуктивных органах, в сердце... Например, вазоинтестинальный пептид (обнаруженный в кишечном тракте, но еще влияющий на тонус кровеносных сосудов, а также связанный с возрастными заболеваниями мозга) синтезируется и в нервной ткани, а давно известный пептидный гормон окситоцин имеет отношение не только к родовой деятельности, но и к регуляции памяти. Кроме того, пептиды могут транспортироваться: образующийся в гипоталамической области мозга аргинин-вазопрессин переносится по ходу нервного волокна в гипофиз, откуда попадает в кровь (целиком или в виде фрагментов). Циркулируя с током крови, пептиды становятся регуляторами по принципу “ВСЕМ-ВСЕМ-ВСЕМ” — доступными тому органу, который готов воспринять этот сигнал.
Теории “специального” пептида, о чем речь шла выше, противостоит идеология АНСАМБЛЯ регуляторных пептидов. Системы пептидов связаны биохимической “родословной” (структурой, биосинтезом, ферментами), сопряжены в функциональном отношении (“подстраховкой”, кооперацией, ограничением функции), наконец, они осуществляют обратную связь после реализации физиологического акта — передают сигнал генному аппарату клетки для стимуляции синтеза новых молекул...
Биохимическая цепочка биогенеза нейропептида завершается его взаимодействием со “своим” рецептором на основе химического закона “узнавания”. С этого момента процесс обретает новое качество — он превращается в результат. Обнаружение специфических для определенного нейропептида рецепторов в какой-либо клетке, ткани, органе означает его причастность к регуляции исследуемого процесса. Рецепторы обнаружены почти для всех физиологически значимых пептидов. Более того, для многих из них установлены подтипы рецепторов, специфичных для различных клеток и для определенных химических вариаций пептида. Типична, например, ситуация, когда фрагмент большой молекулы пептида, образовавшегося в результате ее ферментативного расщепления, становится антагонистом “своего же” рецептора.
Знаменательна история открытия опиоидных (морфиноподобных) нейропептидов. В 1972 г. шотландские фармакологи Костерлиц, Хьюз, Голдштейн и Снайдер обнаружили в нервной ткани рецепторы, лигандные (т.е. предпочтительно связывающиеся) для морфина — вещества, которое не синтезируется в клетках животных. Но это открытие означало: если есть воспринимающая субстанция — рецептор, в организме должен быть и его “контрагент” — вещество, специфически связывающееся с этим рецептором. Так были открыты энкефалины и эндорфины — первые эндогенные опиоидоподобные нейропептиды. Обнаружение специфичных рецепторов означает возможность фармакологического воздействия на физиологическую функцию с помощью веществ, имитирующих структуру пептида, которые будут действовать как антагонисты рецепторов или агонисты-усилители физиологического эффекта. Способ управления физиологической функцией, а значит возможность воздействия на патологические процессы с помощью таких препаратов представляет огромный интерес для современной фарминдустрии, где исследовательские, медицинские и финансовые интересы связаны короткой “упряжкой”.
Классификация
В категорию НП включают обычно малые и средние по размеру пептиды — от 2 до 50-60 аминокислотных остатков. Более крупные пептиды, в число которых входит ряд гормонов, некоторые факторы роста клеток и ряд других факторов, содержат, как правило, свыше 100 а.о., и их относят обычно к категории регуляторных белков. Большинство НП представляет собой линейные пептиды. С-концевые аминокислоты в них нередко амидированы, N-концевые остатки глутаминовой кислоты часто представлены в виде пироглутамата. Другие модификации аминокислотных остатков встречаются редко.
Многие из НП выполняют функции нейромедиаторов, передающих сигнал в пределах синапса, подобно классическим нейромедиаторам непептидной природы. При этом они, как правило, "сотрудничают" с непептидными медиаторами. В одном и том же нервном окончании локализованы определенные комбинации непептидного нейромедиатора с одним, двумя, а иногда и тремя НП. В зависимости от частоты и длительности импульсации они выделяются совместно или раздельно, иногда такие НП называют ко-нейротрансмиттерами или ко-нейромедиаторами. Кроме участия в передаче сигнала в синапсе НП способны осуществлять передачу информации и на более значительные дистанции — в небольших зонах, в органе и даже в пределах целого организма. В этом случае их функции неотличимы от функций гормонов (в том числе гистогормонов). Объектом дистантного действия НП являются пре- и постсинаптические зоны нейронов, а также другие клетки. НП могут при этом облегчать или тормозить передачу импульса и оказывать другие влияния на состояние нейрона, т.е. функционировать как нейромодудяторы.
Классификация нейропептидов основана на сочетании трёх принципов: функционального, структурного и топологического:
Первое семейство
Либерины и статины (релизинг-факторы) гипоталамуса — объединяет общность одной из главных функций, состоящей в регуляции выхода гормонов гипофиза и общность места образования установленного первыми исследователями этих НП. Либерины стимулируют выход (а в ряде случаев и синтез) определенных гормонов из клеток гипофиза, а статины тормозят его. По структуре они весьма разнообразны. Что касается места их образования, то синтез этих НП происходит не только в гипоталамусе, но и во многих других отделах мозга и организма в целом. В гипоталамусе синтезируется та часть либеринов и статинов, преимущественной (но не единственной) функцией которых является действие на гипофиз. Тем не менее ассоциация либеринов и статинов прежде всего с гипоталамусом стала традицией.
Помимо действия на выход гипофизарных гормонов каждый из либеринов и статинов обладает большим числом биологических активностей, осуществляемых прямым действием на определенные нейроны и другие клетки мозга и организма. Так, тиролиберин является мощным стимулятором эмоционального поведения, двигательной активности, дыхательного центра и др.
Люлиберин — главный фактор, усиливающий половое поведение не только через усиление секреции гонадотропных гормонов, но и непосредственным действием на отделы мозга. Кортиколиберин, открытый как индуктор выхода АКТГ, непосредственно подавляет половое поведение и потребление пищи, будучи одновременно антидепрессантом и стимулятором эмоционального поведения. Типичным статином широкого спектра действия является соматостатин. Помимо подавления выхода гормона роста он является умеренным ингибитором настолько большого числа других функций, что его называют иногда пангибином. В их число входит способность подавлять моторику и секрецию желудочно-кишечного тракта, что нашло уже применение при лечении тяжелых форм язвенной болезни, связанных с кровотечениями. Развитие некоторых форм рака (например предстательной железы) тормозится соматостатином.
Второе семейство – опиатыВ отличие от либеринов и статинов эти НП имеют общие особенности структуры. Они содержат в качестве активного центра Met- или Leu-энкефалиновые последовательности или (в группе параэнкефалиновых опиоидов) аналоги этих последовательностей. Первыми были выделены два аналогичных пентапептида, названные энкефалинами и имеющие аминокислотные последовательности:
Tyr-Gly-Gly-Phe-Met Met-энкефалин
Tyr-Gly-Gly-Phe-Leu Leu-энкефалин
Большинство из опиоидов обладает более или менее выраженным обезболивающим действием, реализуемым через рецепторы, впервые описанные в связи с изучением механизмов действия классических непептидных опиатов — морфина, налоксон и др. Однако не менее важными представляются их активности в отношении высших функций мозга: снижение эмоционального поведения ("внутренние нейролептики"), индукция "чувства удовлетворения, вознаграждения", т.е. функция внутренних факторов подкрепления, и др. Некоторые из представителей этого семейства (например "гепта-" и "октапептиды", FMRF и α-эндорфин) ведут себя как частичные антагонисты классических опиоидов. Особенно показателен в этом отношении α-эндорфин. Если β-эндорфин, γ-эндорфин и дез-тирозил-γ-эндорфин снижают эмоциональное поведение, то α-эндорфин его усиливает, вызывая эффекты, подобные эффектам такого психостимулятора, как фенамин. Наконец, такой НП, как β-эндорфин, оказался мощным активатором разновидности лимфоцитов, убивающих раковые клетки, т.е. одним из важных регуляторов системы иммунитета, химическим посредником между этой системой и нервной системой. Заметим, что осуществление такой гуморальной связи — "нервная система — сома" — характерно и для других НП.
Источники опиоидных НП в организме многообразны: большая часть эндорфинов и часть мет-энкефалина образуется в гипофизе; динорфины, неоэндорфииы, леуморфин, риморфин — преимущественно в мозге (или исключительно в нем); адренорфин, "окта-" и "гептапептиды" — преимущественно в надпочечнике; большая часть лей-энкефалина и часть мет-энкефалина имеет источником надпочечник и мозг. Особого упоминания заслуживание такой пищевой источник опиоидных НП, как продукты неполного гидролиза казеина и глютена в желудочно-кишечном тракте, среди которых обнаружены аналоги энкефалинов — казаморфин и глюторфин. Пока неясны количества, в которых они могут образоваться, степень их всасывания и их вклад в баланс опиоидов организма.
Для энкефалинов выполняются некоторые основные критерии для нейромедиаторов: они были определены гистохимически среди других тканей в дорсальном спинном мозге, т.е. в области, ответственной за проведение болевых сигналов. Они найдены в малых промежуточных нейронах, но не на главных нервных путях, где роль медиатора выполняет вещество Р. Это позволяет утверждать, что опиатные пептиды могут ингибировать пресинапсическое высвобождение вещества Р. Энкефалины, благодаря взаимодействию с опиатными μ-рецепторами, блокируют высвобождение субстанции Р, активируя кальций-зависимые калиевые каналы и уменьшая длительность потенциала действия. Другие опиоидные пептиды связываются со вторым подтипом опиатных рецепторов (κ-рецепторов). Они уменьшают высвобождение медиатора, ингибируя потенциалзависимые кальциевые каналы.
Высокие концентрации энкефалинов присутствуют также в лимбической системе, которая участвует в регуляции эмоций.
Опиатные рецепторы уже охарактеризованы как специфические медиаторные рецепторы. Опиатные пептиды конкурируют с синтетическими опиатами за участки связывания. Наиболее высокая связывающая способность обнаружена в нервных окончаниях среднего мозга и таламуса, миндалевидном теле и желатинообразной субстанции спинного мозга.
Также как опиаты, энкефалины тормозят нейрональную активность путём снижения проводимости для ионов Na+, оказывая, по-видимому, прямое влияние на натриевые каналы. Показано, что энкефалины эффективно ингибируют аденилатциклазу, что свидетельствует о роли циклических нуклеотидов как вторичных мессенджеров в каскадных механизмах энкефалинового действия.
Разнообразие биологических свойств энкефалинов позволяет рассматривать их в качестве эндогенных регуляторов многих физиологических и патологических процессов в организме. В связи с этим, изучение механизмов функционирования энкефалинов открывает новые возможности медицинской практики в связи с перспективами их экзогенного введения в организм в качестве средств, идентичных или близких по природе к эндогенным при тех или иных патологиях центральной нервной системы (ЦНС).
Не исключено, что такой широкий спектр биологических свойств опиоидов обусловлен функционированием протеолитических ферментов, участвующих как в процессинге, так и инактивации опиоидов.
Рассматривая пептидные опиоиды, нельзя не упомянуть обнаружение в организме опиоидов непептидной природы. К ним относятся сальсолинол , папаверолин и β-карболины, образующиеся особенно интенсивно при алкоголизме, при нарушение дофаминового цикла, а также некоторые количества кодеина, морфина, 6-ацетил-морфина и некоторых других алкалоидов, находимых в организме животных и человека вне связи с наркоманиями, алкоголизмом и особенностями диеты. Понятно некоторое недоумение, которое эти данные вызывают, но сейчас работы по выявлению этих соединений не вызывают методических возражений. Дискутируется вопрос о месте их синтеза. Предполагается, что синтез некоторых из них могут осуществлять какие-то бактерии в кишечнике из предшественников, поступающих с растительными продуктами.
Третьего семейства НП
Адренокортикотропвый гормон (АКТГ) — давно известен как гипофизарный гормон, функция которого состоит в индукции выделения кортикостероидов из надпочечника. Сейчас, однако, доказаны его синтез в ряде отделов мозга и прямое участие в таких функциях мозга, как уровень внимания к внешним сигналам, запоминание, обучаемость. Установлена ключевая роль для осуществления последних функций фрагмента молекулы АКТГ с 4-го по 7-й а.о. — МЕНF. Этот же фрагмент с той же функциональной характеристикой выявлен у меланотропных пептидов. Синтезирован ряд аналогов МЕНF, которые предполагается использовать для стимуляции внимания и запоминания человека.
Четвертое семейство НПВключает вазопрессины и окситоцины, четко объединяется общностью структуры и преимущественного места секреции — нейрогипофиза. Синтез этих НП происходит главным образом в гипоталамусе, и они представлены в ряде отделов мозга. Вазопрессин давно известен как гормон — дистантный ингибитор диуреза и вазопрессор, а окситоцин —как стимулятор сокращения матки при родовой деятельности. Однако сейчас стало очевидным их участие в формировании долговременной памяти. Дискутируется вопрос о том, является ли это действие прямым или осуществляется через действие на эмоциональное состояние и избирательное внимание. Вазопрессин является при этом стимулятором, а окситоцин — частичным ингибитором.
Пятое семейство
Пятое семейство НП получило название — панкреатические пептиды — в значительной мере по месту первичного обнаружения. Название это, хотя и общепринято, является иногда источником недоразумений, ибо, например, инсулин, являющийся типичным полипептидным гормоном поджелудочной железы, не имеет ничего общего с этими НП ни по структуре, ни по главным функциям. Наиболее изученный представитель панкреатических пептидов — иейропептвд Y — широко представлен не только в органах желудочно-кишечного тракта, но и в головном мозге. Он является участником поддержания тонуса сосудов мозга и организма в целом. Он же стимулятор пищедобывательного поведения; это его свойство неожиданным образом сочетается с некоторым анксиолитическим действием.
Шестое семейство
Наиболее изученный представитель шестого семейства НП— вазоактивный интестинальный пептид (ВИП) — подобно НпY был впервые обнаружен в желудочно-кишечном тракте. Однако именно он (наряду с холецистокинином) содержится в особенно высоких концентрациях в коре головного мозга. Несомненной является его способность снижать тонус гладкой мускулатуры сосудов, в том числе мозга и бронхов. В последнее время показано его участие в формировании полового поведения и функциях органов репродукции. В связи с данными о регуляции сна ВИП отметим существование еще одного пептида, способного усиливать процессы, лежащие в основе сна. В данной классификации не удалось сформировать семейство, в которое входил бы так называемый пептид дельта-сна (ПДС). Он стоит по структуре и другим признакам особняком по отношению к другим НП. Его наиболее выраженное действие проявляется в индукции дельта-волновой активности мозга, регистрируемой электроэнцефалографически и характерной для ортодоксальной фазы сна. Практически важна также его способность облегчать стрессовые состояния.
Седьмое семействоВ коре головного мозга наряду с вазоактивным интестинальным пептидом очень высоко содержание представителя седьмого семейства НП — холецистокинина-8. Первично найденный опять-таки в желудочно-кишечном тракте, он оказался чрезвычайно мощным ингибитором пищедобывательного поведения. Введение его в количествах порядка 1010 — 1012 г в третий желудочек мозга длительно голодавших животных на несколько часов снимало пишедобывательное поведение. Еще меньший по размеру вариант холецистокинина, содержащий всего 4 аминокислотных остатка, оказался одним из внутренних факторов, индуцирующих состояние тревожности и страха.
Восьмое семействоНаиболее изученный представитель восьмого семейства НП — вещество Р —является первым из НП, который был идентифицирован как нейромедиатор в путях проведения сенсорных импульсов. Обладает очень сложным спектром центральных и периферических эффектов, из которых особенно интересно его участие в индукции нейрогенного воспаления. Вещество Р открыто в 1931 г. фон Эйлером и Гаддумом в экстрактах мозга и пищеварительного тракта. Было обнаружено, что оно вызывает сокращение гладких мышц кишечника и расширение кровеносных сосудов, причем для проявления эффекта у человека при внутривенном введении достаточно всего нескольких нанограммов пептида. Позднее вещество Р, выделенное преимущественно из гипоталамуса, было очищено до гомогенного состояния. Его физиологическая роль пока не выяснена. Высвобождение этого пептида после сенсорной стимуляции было показано на дорсальном роговидном отростке спинного мозга. Как уже отмечено, этот процесс ингибируется энкефалином. Вещество Р действует путем деполяризации постсинаптической мембраны и является, следовательно, медиатором сенсорной стимуляции.
Десятое семействоНейротензин — десятое семейство НП — сходен с опиоидными НП по обезболивающей активности (особенно при центральном введении), но реализуется этот эффект не через опиоидные рецепторы; структурным сходством нейротензин не обладает. Характерным является сочетание у нейротензина аналготической активности с гипотермическим и гипотензивным действием — интересным в медицинском плане.
Одиннадцатое семействоСамый мощный из известных гипотермических факторов — бомбезин — является представителем одиннадцатого семейства НП. Глубокое снижение температуры он вызывает при центральном введении. Интерес к нему связан, в частности, с его возможной ролью при зимней спячке.
Двенадцатое и тринадцатое семействаДва НП — брадикинин и ангиотензин II — давно исследуются главным образом как факторы, участвующие в формировании тонуса сосудов. Брадикинин и его аналоги являются вазодилятаторами, ангиотензин II — вазоконстриктор. Известные пути их синтеза не связаны с нервными клетками, хотя ряд эффектов реализуется посредством влияния на синаптическую передачу (например ангиотензин II —на адренергические синапсы). Оба пептида обнаружены в головном мозге, причем установлена способность ангиотензина II возбуждать жажду при центральном введении.
Четырнадцатого семействоНазвания НП четырнадцатого семейства нередко приводят к недоразумениям. Так, в англоязычной литературе они называются "calcitonin gene related peptides" и обозначаются как СGRP. В данном случае точный перевод — "пептиды, кодируемые геном, подобным гену кальцитонина" — является слишком громоздким. Поэтому все чаще используется в качестве
тривиального названия термин ко-кальцигенин. Биологическая активность ко-кальцигенина проявляется очень наглядно — введение всего лишь около 1014 моля этого НП под кожу человека вызывает довольно длительное расширение артериол, усиление кровотока и (особенно совместно с веществом Р) диапедез из сосудов форменных элементов крови. Вместе с ВИП и НпY ко-кальцигенин участвует в регуляции тонуса сосудов мозга, коронарных сосудов и др.
Пятнадцатое семействоНП пятнадцатого семейства — атрнопептиды — до последнего времени считались факторами сугубо периферического происхождения и действия. Главный их источник — кардиомиоциты предсердия. Атриопептиды значительно усиливают диурез и особенно натрий-урез. Сейчас, однако, доказано, что атриопептиды синтезируются и в головном мозге. Таким образом, история их исследования подобна описанной выше для ряда пептидов, обнаруженных сначала на периферии, а затем в головном мозге. На очереди — выявление их функций в ЦНС.
Шестнадцатое семействоНаконец, НП шестнадцатого семейства — эндозепины — являются негативными регуляторами рецепторов GABA. Если сама GABA является тормозным медиатором, участвующим в разнообразных процессах, в том числе в транквилизации — снижении тревожности, успокоении, то эндозепины, напротив, вызывают тревожность и проконфликтное поведение. По-видимому, подавление GABA-ергической трансмиссии, очень широко представленной в мозге, позволяет предполагать у эндозепинов и другие проявления биологической активности, но исследования этой группы НП начаты сравнительно недавно.
Пептиды - коннекторы
Описан целый ряд пептидов, так называемых пептидов-коннекторов, которые ряд исследователей считают непосредственными детерминантами формирования определенных условных рефлексов и довольно сложных навыков. Эту точку зрения разделяют не все исследователи. Так некоторые из них полагают, что пептиды-коннекторы являются не детерминантами памяти, а регуляторами некоторых специфических форм врожденного поведения животных. Пептиды-коннекторы представляют исключительный интерес. Наиболее изученными из них являются амелитин, скотофобин, хромодиопсины и катабатмофобин. Все они получены из мозга животных, тренированных к тому или иному навыку. При введении в мозг они сообщают необученному животному тот же навык.
Амелитин, по-видимому, имеющий строение Глу-Глу-Гли-Тир-Сер-Лиз образуется в мозгу белых крыс при привыкании к резкому звуку определенной частоты и продолжительности. После введения амелитина, выделенного из этого животного, необученной крысе, она не реагирует на резкий звук той же частоты и продолжительности. Эффективная доза — 10 нг.
Скотофобин, вероятно, имеет следующее строение:
Сер-Асп-Асн-Асн-Глн-Глн-Гли-Лиз-Сер-Ала-Глн-Глн-Гли-Гли-Тир. Он образуется в мозгу белых крыс при воспитании у них страха перед темной частью лабиринта. После введения природного препарата белые крысы и мыши, а также рыбы избегают темноты.
Хромодиопсины образуются в мозгу золотых рыбок при выработке у них рефлексов избегания синей или зеленой стенки аквариума. Этот рефлекс передается необученным золотым рыбкам после введения им хромодиопсина. Химический состав хромодиопсинов не установлен. Хромодиопсин «к зеленому цвету» расщепляется трипсином и химотрипснном, а хромодиопсин «к синему цвету» устойчив к трипсину и расщепляется химотрипсином.
Катабатмофобин имеет молекулярный вес 1700—1950, образуется в мозгу белых крыс при формировании двигательно-оборонительного рефлекса избегания определенной последовательности движений. Введение катабатмофобина сообщает необученным белым крысам этот рефлекс. Механизм действия этой интереснейшей группы соединений пока неясен. После введения животному скотофобин преимущественно локализуется в коре. Высказывались предположения, что действие этих пептидов основано на специфическом связывании с определенными небольшими группами синапсов, надолго повышающем их проводимость (отсюда и термин—пептид-коннекторы). Для изучения этого класса пептидов требуется четкое установление их структуры.
Система регуляторных пептидов образует так называемый функциональный континуум. Это обозначает то, что, с одной стороны, каждый из пептидов обладает уникальными свойствами, уникальным комплексом активностей. С другой стороны, многие проявления биоактивности каждого из пептидов совпадают или близки к таковым ряда других пептидов. В результате каждый пептид выступает как созданный эволюцией "пакет программ" для включения или модуляции определенного комплекса функций. Набор таких комплексов настолько велик, что создается возможность относительно плавного, непрерывного перехода от одного к другому комплексу совместимых функций. Представление о функциональном континууме пептидных регуляторов позволяет понять биологический смысл их необычайного многообразия. Указанное выше число уже открытых пептидных регуляторов, по-видимому, значительно меньше их действительной численности, если учесть, что каждый год приносит открытие не только ряда пептидов, относящихся к уже известным семействам, но и новых семейств пептидов.
Кроме обеспечения разнообразнейших комплексов биоактивностей пептидный континуум выполняет ещё одну функцию — образование сложных регуляторных цепей и каскадов. Каждый из НП обладает способностью индуцировать выход в кровь, в цереброспинальную жидкость, в межклеточные среды организма определенных НП. Частным случаем этой системы взаимной индукции является описанное действие либеринов и статинов гипоталамуса на выход гормонов гипофиза. Каждый НП, выход которого индуцирован другим пептидом, в свою очередь может индуцировать выход ряда следующих НП, так что возникает цепной, каскадный регуляторный процесс. Сейчас трудно судить о том, сколь длинной может быть такая цепь. Известно, однако, что многие НП, период полуэллиминации которых измеряется минутами, способны вызывать многочасовые и даже многосуточные эффекты после введения в организм, Вероятно, основой этого и являются такие цепные процессы. Биологический смысл существования длительных регуляторных процессов, складывающихся из кратковременных звеньев, очевиден. В отличие от систем, основанных на долгоживущих регуляторах, такая система является более гибкой в меняющейся ситуации, при поступлении новых сигналов и т.п.
В сложной системе индуцирующего действия НП на выход других НП необходимо отметить ряд элементов иерархии, "взаимоподчинения" одних регуляторов другим. Например, ВИП и ХЦК-8, локализованные в значительных количествах в коре головного мозга, способны индуцировать или тормозить выход ряда либеринов и статинов гипоталамуса, который, в свою очередь, регулирует выход гормонов гипофиза. Многие из последних тоже служат индукторами выхода периферических гормонов (щитовидной железы, надпочечников и др.). Такая иерархия, контроль и управление "сверху-вниз" сочетается со сложной системой прямых действий всех НП на многие функции организма.
Сколь бы сложной ни представлялась эта картина действия и взаимодействия пептидов, следует постоянно помнить о существовании диалектического сочетания их прямых и опосредствованных каскадных эффектов.
Процессинг и посттрансляционная модификация нейропептидовОбщий принцип синтеза всех достаточно изученных НП состоит в образовании относительно больших пептидов-предшественников, из которых после завершения трансляции выщепляются протеазами соответствующие НП. Как правило, в состав пептида-предшественника входит одна или большее число последовательностей НП (иногда весьма различных по структуре) и так называемая сигнальная последовательность, способствующая миграции предшественника внутри клетки после завершения трансляции в шероховатой эндоплазматической сети и отщепляемая в конце этой части пути.
В 1984 году Gainer сформулировали гипотезу секреторных везикул, согласно которой процессинг биологически активных пептидов осуществляется в процессе передвижения молекул пропептидов по гранулярному ЭПР, комплексу Гольджи и в секреторных везикулах. Последние содержат полный набор ферментов процессинга, специфичный для данной клетки (ткани), а также специальные системы поддержки оптимальной рН среды.
Процессинг нейропептидов внутри секреторных гранул включает в себя эндо- и экзопротеолитические реакции, а также гликолизирование, сульфатирование, амидирование и фосфорилирование N- и С-концевых аминокислотных остатков, что предохраняет образовавшийся продукт от дальнейшей деградации.
Многие пептиды-предшественники образуют промежуточное соединение с гликозидами, которые оказывают стабилизирующее действие на некоторых стадиях процессинга и влияют на выбор мест атаки протеиназами. Протеиназы, обеспечивающие выщепление НП в течение нескольких десятков минут — часа после трансляции, не являются высокоспецифичными. В значительной мере точное выщепление обусловлено наличием в последовательности аминокислот пептидов-предшественников парных остатков. Они в первую очередь атакуются протеиназами, близкими по характеру действия к катепсину В и трипсину. Выщепляемые в этой фазе процессинга фрагменты во многих случаях уже представляют собой активные НП. Нередко эти фрагменты подвергаются дальнейшему протеолизу опять-таки с образованием новых НП. На этой фазе описано участие не только трипсиноподобных протеиназ, но и ряда других протеиназ, специфичность которых тоже не является очень высокой. Однако в различных нейронах и, возможно, даже в разных терминалях одного нейрона их набор может быть неодинаковым, что обеспечивает в конечном счете ту или иную специализацию на продукции строго определенных НП (единичных или их комплексов).
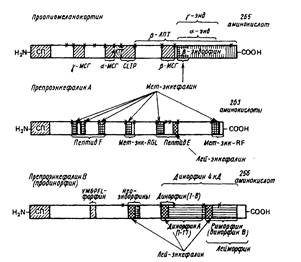
Так например, многие эндопептидазы, участвующие в процессинге энкефалинов обладают очень узкой субстратной специфичностью. Так, эндопептидаза, специфичная к связи Lys-Arg, не расщепляет связи Arg-Arg и Arg-Lys, что объясняют специфичностью фермента не только к гидролизуемой связи, но и, вероятно, к ближайшему аминокислотному окружению. Расщепление может происходить иногда не по парам остатков основных аминокислот, а по моноостаткам аргинина, лизина или каких-либо других аминокислот. Например, тиоловая прогормон-конвертаза расщепляет предшествен-ники энкефалинов с обеих сторон по моно- и диосновным остаткам аминокислот, но более высокую специфичность проявляет к моноосновным сайтам. Все этапы процессинга хорошо иллюстрируются схемой превращений проопиомеланокортина, из которого могут формироваться β-липотропин, АКТГ1-39, АКТГ1-24, АКТГ18-39 (так называемый CLIP), α, β- и γ-МSH, β-, γ- и α-эндорфины, Met-энкефалин, дезтирозильные производные эндорфинов, С-концевой дипептид β-эндорфина и, возможно, еще некоторые другие НП. Еще раз подчеркнем, что имеется в виду не одновременное образование всех этих НП в одном нейроне или терминали, а более или менее избирательное формирование их в зависимости от набора протеаз в данной клетке.
Обращает на себя внимание, что синтез ряда опиоидных пептидов, содержащих в качестве минимальной активной последовательности Met-энкефалин, обеспечивается по крайней мере двумя пептидами-предшественниками — проопиомеланокортином (локализованным главным образом, но не исключительно в гипофизе) и препроэнкефалином А (локализованным преимущественно, но опять-таки не исключительно в надпочечниках). Сходным образом лей-энкефалинсодержащие опиоидные НП выщепляются из мозгового пептида-предшественника продинорфина (препроэнкефалина В) и из уже упоминавшегося препроэнкефалина А, совмещающего в себе последовательности НП с лей-энкефалиновыми и мет-энкефалиновыми активными центрами. Однако общность минимальных последовательностей опиоидных НП, заключенных в разных пептидах-предшественниках, не означает идентичности содержащих их НП большого размера: продинорфин образует лей-морфин и перекрывающийся с ним каскад динорфинов разного размера (динорфин В (риморфин) и 4кД-ди-норфин), а также α- и β-неоэндорфины. Из препроэнкефалина А формируются самостоятельные НП, содержащие и лей-энкефалин, и мет-энкефалин (например так называемый пептид Е).
Сходные закономерности структуры можно отметить и для некоторых других пептидов-предшественников, хотя они, как правило, исследованы в гораздо меньшей степени, чем предшественники опиоидов, АКТГ и МСГ. Можно полагать, что дальнейший анализ биологической активности их фрагментов позволит выявить источники новых НП.
Обращают на себя внимание большие области пептидов-предшественников, где пока не идентифицированы НП. Хотя часть из них выполняет (судя по β -эндорфину, АКТГ) функции дополнительных "адресных" последовательностей, уточняющих взаимодействие НП преимущественно с определенной тканью иди органом, есть основания предполагать и в них наличие последовательностей еще не идентифицированных НП. В ряде случаев образование НП и процессы их распада переплетаются друг с другом. Например, один из путей распада β-эндорфина - НП, имеющего самостоятельное функциональное значение, ведет, как уже упоминалось, к образованию γ-эндорфина и опять-таки НП, имеющего уникальную функциональную характеристику; отщепление концевого аминокислотного остатка от γ -эндорфина приводит к образованию α-эндорфина, весьма своеобразного по функциям, и т.д.
Регуляция метаболизма активных регуляторных пептидов определяется большим спектром воздействий, изменяющих гомеостаз на любом уровне - уровне клетки (транскрипция, трансляция и посттрансляционный процессинг), ткани (секреция и инактивация нейропептидов), а также на уровне организма в целом. Именно эти морфогенетические и биохимические особенности биогенеза и определяют уровень активных регуляторных пептидов в организме. При этом, несомненно, важная регулирующая роль в метаболизме нейропептидов принадлежит протеолитическим ферментам.
Метаболизм энкефалинов и ферменты их процессинга
Как было отмечено выше, известны три белковые молекулы, включающие в свою структуру последовательности энкефалинов: проопиомеланокортин, препроэнкефалин А (проэнкефалин), препроэнкефалин В (продинорфин). Последовательности, определяющие структуру энкефалина, входящего в состав молекулы предшественника, фланкированы парами основных аминокислот - аргинина или лизина. Структуры типа Arg-Lys, Arg-Arg, Lys-Lys, Lys-Arg узнаются ферментативными комплексами, осуществляющими протеолиз. В результате действия как эндо-, так и экзопептидаз секреторных везикул, так и внеклеточных ферментов, локализованных на внешней поверхности мембраны и в биологических жидкостях организма, происходит полное освобождение активного пептида из его предшественника.
Протеолитические ферменты являются специфичными на каждом этапе процессинга. Выделяют множество различных везикулярных эндопептидаз, осуществляющих специфическое ращепление: прогормон-конвертазы 1/3 и 2, динорфин-превращающий фермент, тиоловая прогормон-конвертаза, энкефалинобразующий фермент, IRCM-сериновая протеиназа I. Ряд внеклеточных ферментов также принимают участие в процессинге энкефалинов: эндопептидаза 24.15 (КФ 3.4.24.15), эндоолигопептидаза А (КФ 3.4.22.19), а также ангиотензинпревращающий фермент (АПФ, КФ 3.4.15.1), проявляющий наряду с эндопептидазной и дипептидилкарбоксипептидазную активность.
После действия эндопептидаз образуются пептиды, фланкированные остатками основных аминокислот, для отщепления которых необходимы специфические амино- и/или карбоксипептидазы. В процессинге энкефалинов важную роль играют экзопептидазы секреторных везикул, такие как аминопептидазо-В-подобные и карбоксипептидазо-В-подобные ферменты.
Одним из основных ферментов конечной стадии процессинга энкефалинов в мозге является карбоксипептидаза H. Этот фермент катализирует расщепление остатков основных аминокислот с С-конца белков пептидов предшественников, превращая их в активные формы. Локализация КП Н совпадает с распределением проэнкефалинов, регуляторных пептидов и их предшественников (адренокортикотропного гормона, вещества Р, вазопрессин, окситоцина, инсулина, глюкагона , натрийуретического фактора предсердий).
Сходной с КП Н субстратной специфичностью обладает фермент, получивший своё название от индивидуального ингибитора: PMSP(фенилметилсульфонилфторид)-ингибируемая карбоксипептидаза. Физико-химические свойство данного фермента сходны с таковыми у лизосомальной КП А. Известно, что карбоксипептидаза Н проявляет большее сродство к тем субстратам, у которых перед остатком основной аминокислоты находится остаток аланина или глицина, а PMSP-ингибируемая карбоксипептидаза преимущественно расщепляет субстраты, у которых предпоследними являются остатки лейцина и метионина. Кроме того, показано, что Кm КПН по отношению к DNS-Phe-Ala-Arg почти в 3 раза выше, чем по DNS-Phe-Leu-Arg, а Кm ФМСФ-ингибируемой КП по DNS-Phe-Ala-Arg почти в 2 раза ниже, чем по DNS-Phe-Leu-Arg. Поэтому можно предполагать, что КПН и PMSP-ингибируемая КП, участвуя в процессинге, отдают предпочтение различным регуляторным пептидам, при этом проэнкефалины являются, вероятно, более предпочтительным субстратом для PMSP-ингибируемой КП, нежели чем для КПН.
Помимо вышеперечисленных экзопептидаз секреторных везикул, в процессинг энкефалинов вовлекаются и внеклеточные экзопептидазы: карбоксипептидаза N (КФ 3.4.12.7) и карбоксипептидаза М.
Таким образом, в результате последовательного действия вышеуказанных ферментов, из высокомолекулярных предшественников высвобождаются активные энкефалины. Под влиянием какого-либо стимула (медиаторы, другие регуляторные пептиды, cAMP), возникающего по типу обратной связи или поступающего от других секреторных клеток, происходит изменение концентрации ионов Ca2+. Это приводит к выделению из клетки активных энкефалинов, которые мигрируют к клеткам-мишеням (через кровяное русло или синаптическую щель), где связываются со специфическими рецепторами. В последствии, они подвергаются расщеплению различными пептидазами, что приводит к их модификации или же к полной потере их биологической активности.
Дезактивация энкефалинов происходит при участии ферментов внешней поверхности мембраны: энкефалиназа А, нейтральная эндопептидаза 24.11, КФ 3.4.24.11), аминопептидаза М (КФ 3.4.11.2) и ариламидаза (энкефалинаминопептидаза, КФ 3.4.11.?).
Таким образом, очевидна взаимосвязь и взаимовлияние регуляторных пептидов и протеолитических ферментов в организме: энкефалины способны инициировать эндогенные механизмы регуляции активности ферментов, в свою очередь протеолитические ферменты способны регулировать уровень активных форм пептидов в организме, а также участвовать в «запуске» реакций регуляторного континуума, обуславливая нейромодуляторные свойства и биологическую роль регуляторных пептидов. Поэтому, учитывая тесную взаимосвязь пептидгидролаз с регуляторными пептидами и, в частности – с энкефалинами, можно заключить, что изменение активностей протеолитических ферментов при различных функциональных состояниях организма, развитии патологий и влиянии некоторых экстремальных факторов, является неотъемлемой частью биологического действия этих пептидов, в определенной степени объясняющей индивидуальные особенности их физиологических свойств в организме.
Краткие характеристики карбоксипептидазо-В-подобных ферментов
В связи с участием в процессинге, модификации и инактивации биологически активных пептидов, последнее время интенсивно исследуется семейство основных карбоксипептидаз. Они называются карбоксипептидозо-В-подобными ферментами и их субстратная специфичность связана с отщеплением остатков основных аминокислот с С-конца пептидов предшественников.
В организме млекопитающих обнаружено несколько форм этих ферментов.
Сarboxypeptidase U (M14.009)| Другие названия | HBCPB, TAFI, arginine carboxypeptidase, brain-specific carboxypeptidase B, carboxypeptidase R, plasma carboxypeptidase B, thrombin-activatable fibrinolysis inhibitor |
| Classification | Clan MC >> Subclan (none) >> Family M14 >> Subfamily A >> M14.009 |
| Activity | |
| Catalytic type | Металл |
| NC-IUBMB | Subclass 3.4 (Peptidases) >> Sub-subclass 3.4.17 (Metallocarboxypeptidases) (Metallocarboxypeptidases) >> Peptidase 3.4.17.2 |
| Activity status | Человек: активна Крысы: активна |
| Physiology | Фибринолитическая активность. Деградация активных пептидов и возможно β–амилоида 1-42. |
| Human genetics | |
| Gene symbol | CPB2 |
| Locus | 13q14.11 |
| Mouse genetics | |
| Gene symbol | Cpb2 |
| Position | 14:D2 |
| Substrate | Bz-Gly-Arg (hippuryl-Arg) Bz-Gly-Lys (hippuryl-Lys) coagulation factor X light chain coagulation factor X light chain fibrin derived peptide furylacryloyl-Ala-Arg (FAAR) furylacryloyl-Ala-Lys (FAAK) Protein S100-A10 |
| Другие названия | CPN1 g.p. (Homo sapiens), anaphylatoxin inactivator, arginine carboxypeptidase, carboxypeptidase K, creatine kinase conversion factor, kininase I, lysine carboxypeptidase, plasma carboxypeptidase B |
| Classification | Clan MC >> Subclan (none) >> Family M14 >> Subfamily B >> M14.004 |
| Catalytic type | Metallo |
| NC-IUBMB | Subclass 3.4 (Peptidases) >> Sub-subclass 3.4.17 (Metallocarboxypeptidases) >> Peptidase 3.4.17.3 |
| Activity status | Крыса: активна Человек: активна |
| Physiology | Деструкция вазоактивного пептида |
| Human genetics | |
| Gene symbol | CPN1 |
| Locus | 10 |
| Mouse genetics | |
| Gene symbol | Cpn |
| Position | 19:D2 |
| Substrate | Dns-Ala-Arg Hemoglobin subunit alpha furylacryloyl-Ala-Lys Protein S100-A10 C3A anaphylotoxin C4A anaphylotoxin bradykinin |
S10.005: serine carboxypeptidase D
| Другие названия | carboxypeptidase D, serine, carboxypeptidase II, wheat, carboxypeptidase WII |
| Классификация | Clan SC >> Subclan (none) >> Family S10 >> Subfamily (none) >> S10.005 |
| Каталитический тип | Serine |
| NC-IUBMB | Subclass 3.4 (Peptidases) >> Sub-subclass 3.4.16 (Serine-type carboxypeptidases) >> Peptidase 3.4.16.6 |
| Substrate | furylacryloyl-Ala-Lys furylacryloyl-Phe-Leu |
| Структура 3D |
|
M14.005: carboxypeptidase E
| Другие названия | carboxypeptidase H, cobalt-stimulated chromaffin granule carboxypeptidase, crino carboxypeptidase B, enkephalin convertase, insulin granule-associated carboxypeptidase |
| Классификация | Clan MC >> Subclan (none) >> Family M14 >> Subfamily B >> M14.005 |
| Каталитический центр | Металл |
| NC-IUBMB | Subclass 3.4 (Peptidases) >> Sub-subclass 3.4.17 (Metallocarboxypeptidases) >> Peptidase 3.4.17.10 |
| Физиология | Участвует в биосинтезе пептидных гормонов и нейротрансмиттеров |
| Human genetics | |
| Gene symbol | CPE |
| Locus | 4 |
| Mouse genetics | |
| Gene symbol | CPE |
| Position | 8:B3.1 |
Substrate
Ac-Tyr-Ala-Arg
Bz-Phe-Ala-Arg
Dns-Phe-Ala-Arg
M14.006: carboxypeptidase M| Классификация | Clan MC >> Subclan (none) >> Family M14 >> Subfamily B >> M14.006 |
| Каталитический тип | Metallo |
| NC-IUBMB | Subclass 3.4 (Peptidases) >> Sub-subclass 3.4.17 (Metallocarboxypeptidases) >> Peptidase 3.4.17.12 |
| Физиология | Деградация биоактивных пептидов |
| Human genetics | |
| Gene symbol | CPM |
| Locus | 12q15 |
| Mouse genetics | |
| Gene symbol | 5730456K23Rik |
| Position | 10:D2 |
| Substrate | Bz-Ala-Lys Bz-Gly-Arg Bz-Gly-Lys Dns-Ala-Arg dynorphin A1-13 [Leu5]-enkephalin-Arg [Met5]-enkephalin-Lys bradykinin |
PMSP-ингибируемая карбоксипептидаза
Пока ещё мало изученный фермент, который расщепляет dansyl-Phe-Leu-Arg и отличается по молекулярной массе (100-110 кДа) от других карбоксипептидаз. Он имеет близкий к КР Н оптимум pH = 5,6 – 6,0. Не является метало ферментом, т.к. не ингибируется ЭДТА. Возможная биологическая роль данного фермента, это процессинг нейропептидов, в частности энкефалинов, хотя и этот вопрос довольно спорный. В настоящее время ведутся работы по выделению и очистке этого фермента, с целью изучения кинетики его действия м активного центра.
Заключение
Пептиды — одна из важнейших систем регуляции ГОМЕОСТАЗА. Этот термин, введенный в 30-х годах американским физиологом У.Кенноном, означает жизненно важное равновесие всех систем организма. По мере усложнения наших представлений о нормальной, а тем более патологической, физиологии это понятие уточнили как ГОМЕОКИНЕЗ, т.е. подвижное равновесие, баланс постоянно меняющихся процессов. Организм соткан из миллионов “гомеокинезиков”. Эта огромная живая галактика определяет функциональный статус всех органов и клеток, которые связуются регуляторными пептидами. Как мировая экономическая и финансовая системы — множество фирм, производств, заводов, банков, бирж, рынков, магазинов... А между ними — “конвертируемая валюта” — нейропептиды.
Все клетки организма постоянно синтезируют и поддерживают определенный, функционально необходимый, уровень регуляторных пептидов. Но когда случаются отклонения от “стационарности”, их биосинтез (в организме в целом или в отдельных его “локусах”) либо усиливается, либо ослабевает. Такие колебания возникают постоянно, если речь идет об адаптивных реакциях (привыкании к новым условиям), выполнении работы (физических или эмоциональных действиях), состоянии предболезни — когда организм “включает” повышенную защиту от нарушения функционального баланса.
Говоря о заболеваниях мозга, следует заметить, что в последние годы снова появилась информация о пептидах, особо причастных к какой-либо центральной патологии. В этом перечне — аллергический энцефалитогенный пептид, β-амилоид болезни Альцгеймера, нейротрофический пептид поврежденного мозга и др. Все они выделены из мозга, который охвачен соответствующей патологией, и имитируют таковую при перенесении экспериментальным животным. Для каждого из них установлена точная химическая структура, выявлены специфические рецепторы; но речь не идет о каких-то особых субстанциях. Эти пептиды, как правило, “осколки” миэлина, основного белка мозга, необходимого для его нормальной работы. Их присутствие служит иллюстрацией, как при определенных заболеваниях, вероятно вследствие инверсии нормального биосинтеза пептидов, они становятся фактором “зла”, провоцирующим развитие болезни в новой и еще более пагубной форме. Однако их выявление помогает найти конкретные с точки зрения нейрохимии причины патологии и, значит, подойти к возможности ее предупреждения и лечения.
Таким образом, выявлена огромная, хотя еще почти не изученная роль пептидов в функционировании нервной системы. Они оказывают влияние на возбудимость нервной ткани, выполняя роль медиаторов и участвуя в возвратном ингибировании нейронов выступают в роли модуляторов различных процессов, участвуют в создании межнейрональной связи. Пептиды могут быть не только кратковременными медиаторами химической передачи, но и долговременными регуляторами свойств мембраны и синаптического действия. Их действие на нейрональную активность часто выражается в изменении поведенческих реакций животных. Влияние пептидов на различные функции организма — на сосудистые реакции, высвобождение моноаминов, болевые реакции организма, терморегуляцию и, наконец, на сон, память и т. д, позволило предположить регулирование важнейших функций животных пептидными факторами. Связь пептидов с трансмиттероактивными аминокислотами и с моноаминами открывает большие возможности для химического кодирования в нервной системе.
Приведенные характеристики биологической активности НП —представителей различный семейств — далеко не исчерпывают всего многообразия их функций.
В заключение хочется обратиться к перу нейрогистолога, нобелевского лауреата С. Рамон-и-Кахаля, мыслившего образно и точно: “...сад неврологии представляет исследователю захватывающий, ни с чем не сравнимый спектакль. В нем мои эстетические чувства находили полное удовлетворение. Как энтомолог, преследующий ярко окрашенных бабочек, я охотился в красочном саду серого вещества мозга за клетками с их тонкими элегантными формами, таинственными бабочками души, биение которых, быть может, когда-то — кто знает? — прояснит тайну духовной жизни”.
Литература
1. Malcolm Dixon, Edwin C. Webb “Enzymes” Longman Group Ltd. 1979.
2. Р. Шмидт, Г. Тевс «Физиология человека» Москва «Мир» 1996
3. Ferdinand Hucho “Neurochemistry. Fundamentals and Concepts” VCH Verlagsgesellschaft mbH, D-6940 Weinheim (Federal Republic of Germany), 1986
4. И. П. Ашмарин и соавт. «Нейрохимия» Москва Изд. Института биомедициеской химии РАМН, 1996.
5. М. И. Прохорова «Нейрохимия» Ленинград, 1979
6. А. Уайт, Ф. Хендлер, Э. Смит, Р. Хилл, И. Леман «Основы биохимии» Москва: «Мир», 1981
7. Albert L. Lehninger “Principles of biochemistry” Worth Puplishers, Inc 1982
8. В. К. Плакунов «Основы энзимологии» М.: Логос, 2001.
9. Дж. Г. Николс, А.Р. Мартин, Б. Дж. Валлас, П.А. Фукс «От нейрона к мозгу» М.: Едиториал УРСС, 2003.
10. О.А. Гомазков «Нейропептиды — универсальные регуляторы. Почему?» //Природа № 4, 1999.
11. Л.Ф. Панченко, Н.В. Митюшина, Н.В. Фирстова, М.Т. Генгин «Метаболизм энкефалинов при различных функциональных и патологических состояниях организма» http://medi.ru/pbmc/8890401.htm
12. А.Н.Вернигора, Н.В. Щетинина, М.Т. Генгин «Исследование активности основных (отщепляющих остатки аргинина и лизина) карбоксипептидаз у крыс разного возраста» // «Биохимия» № 10 т. 61, вып. 10, 1996
13. Н. Грин, У. Стаут, Д. Тейлор «Биология» М.: Мир, 1996.
14. Марри Р., Греннер Д., Мейес П., Родуэлл В. «Биохимия человека» М.: Мир, 2004
15. A.N. Vernigora, N.N. Nikishin, M.T. Gengin “Proteolytic Enzymes and Regulation of the levels of Active Neuropeptides” //Biochmistry (Moscow), Vol. 60, No. 10, 1995
16. Walker,J.B. & Bajzar,L. Complete inhibition of fibrinolysis by sustained carboxypeptidase B activity. The role and requirement of plasmin inhibitors. J Thromb Haemost (2007)
17. Aviles,F.X. & Vendrell,J. Carboxypeptidase B. In Handbook of Proteolytic Enzymes, 2 edn (Barrett,A.J., Rawlings,N.D. & Woessner,J.F. eds), p.831-833, Elsevier, London (2004)
18. .N. Vernigora, M.T. Gengin «Proteolytic Enzymes: Subcellular Location, Properties, and involvement in Neuropeptide Turnover. A Review» // Biochemistry (Moscow) Vol. 61, No 5. 1996
19. Ferrer,M., Zuck,P., Kolodin,G., Mao,S.S., Peltier,R.R., Bailey,C., Gardell,S.J., Strulovici,B. & Inglese,J. Miniaturizable homogenous time-resolved fluorescence assay for carboxypeptidase B activity. Anal Biochem (2003) 317, 94-98
20. Muller,C.A., Appelros,S., Uhl,W., Buchler,M.W. & Borgstrom,A. Serum levels of procarboxypeptidase B and its activation peptide in patients with acute pancreatitis and non-pancreatic diseases. Gut (2002) 51, 229-235
21. Mock,W.L. & Xu,D.H. Catalytic activity of carboxypeptidase B and of carboxypeptidase Y with anisylazoformyl substrates. Bioorg Med Chem Lett (1999) 9, 187-192
22. А.Н. Вернигора, М.Т. Генгин «Частичная характеристика основной фенилметилсульфонилфторид-ингибируемой карбоксипептидазы из головного мозга кошки» //Биохимия т. 60, вып 11, 1995
23. A.N. Vernigora, N.N. Nikishin, M.T. Gengin «On the interaction between the activities of carboxypeptidase H and angiotensin-converting enzyme» // Biochemistry (Moscow) Vol. 60, No 1, 1995
24. Marzban,L., Soukhatcheva,G. & Verchere,C.B. Role of carboxypeptidase E in processing of pro-islet amyloid polypeptide in {beta}-cells. Endocrinology (2005) 146, 1808-1817
25. Fricker,L.D. Carboxypeptidase E. In Handbook of Proteolytic Enzymes, 2 edn (Barrett,A.J., Rawlings,N.D. & Woessner,J.F. eds), p.840-844, Elsevier, London (2004)
26. Fricker,L.D., Plummer,T.H., Jr. & Snyder,S.H. Enkephalin convertase: potent, selective, and irreversible inhibitors. Biochem Biophys Res Commun (1983) 111, 994-1000
27. Fricker,L.D. & Snyder,S.H. Purification and characterization of enkephalin convertase, an enkephalin-synthesizing carboxypeptidase. J Biol Chem (1983) 258, 10950-10955
28. Fricker,L.D. & Snyder,S.H. Enkephalin convertase: purification and characterization of a specific enkephalin-synthesizing carboxypeptidase localized to adrenal chromaffin granules. Proc Natl Acad Sci U S A (1982) 79, 3886-3890.
Похожие работы
... сенсорную стимуляцию. Однако подобный дефект непостоянен, постепенно веточки из соседних, интактных нейронов возмещают дефект иннервации. Взаимодействие чувствительных и двигательных нейронов В нервной системе беспозвоночных синапсы между нейронами обычно располагаются не на соме, а на отростках в центральной части ганглия (область нейропиля). Синаптические потенциалы из нейропиля ...
... . Обзор работ по этим ферментам, который будет представлен ниже, является свидетельством большого интереса к пептидгидролазам нервной ткани нелизосомальной локализации, и вместе с тем это лишь первые шаги в выяснении функциональной роли этой группы пептидгидролаз. Характеристика протеолитических ферментов нервной ткани нелизосомальной локализации и их биологическая роль Пептидгидролазы ...
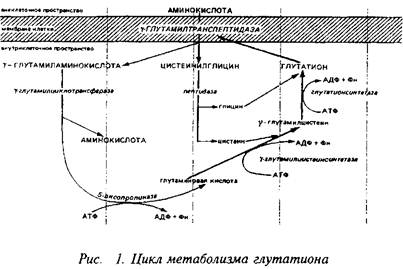
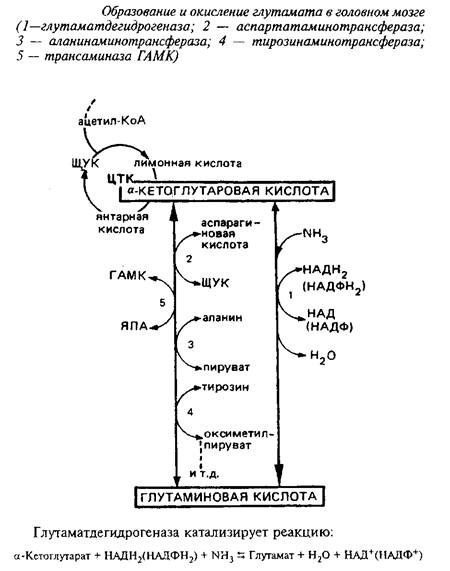
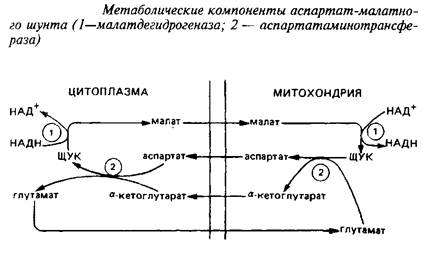
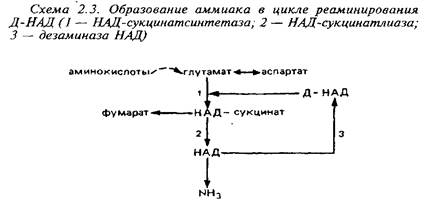
... видов животных. Региональные различия в концентрации метионина невелики. Влияние диеты на концентрацию метионина в мозге также незначительно из-за конкурентных отношений с нейтральными аминокислотами за транспортные системы. Метионин в пуле свободных аминокислот утилизируется на 80% для синтеза белка. Метаболизм свободного метионина до цистеина начинается с образования S-аденозилметионина, ...
... , кроме того, в олигодендроцитах до миелинизации и в миелине периферической нервной системы. В ЦНС человека он представлен тремя полипептидными цепями с Мг=92, 107, 113 кД, а в периферической нервной системе – одним белком с Мг=107 кД. МАГ относится к гликопротеинам с относительно низким содержанием углеводных остатков – около 30% от массы молекулы, но содержит характерный для гликопротеинов набор ...
0 комментариев