Навигация
Строение бактериальной клетки
25373
знака
0
таблиц
0
изображений
3. Строение бактериальной клетки
Обязательными органоидами являются: ядерный аппарат, цитоплазма, цитоплазматическая мембрана.
Необязательными (второстепенными) структурными элементами являются: клеточная стенка, капсула, споры, пили, жгутики.
1. В центре бактериальной клетки находится нуклеоид - ядерное образование, представленное чаще всего одной хромосомой кольцевидной формы. Состоит из двухцепочечной нити ДНК. Нуклеоид не отделен от цитоплазмы ядерной мембраной.
2. Цитоплазма- сложная коллоидная система, содержащая различные включения метаболического происхождения (зерна волютина, гликогена, гранулезы и др.), рибосомы и другие элементы белоксинтезирующей системы, плазмиды (вненуклеоидное ДНК), мезосомы (образуются в результате инвагинации цитоплазматической мембраны в цитоплазму, участвуют в энергетическом обмене, спорообразовании, формировании межклеточной перегородки при делении).
3. Цитоплазматическая мембрана ограничивает с наружной стороны цитоплазму, имеет трехслойное строение и выполняет ряд важнейших функций- барьерную (создает и поддерживает осмотическое давление), энергетическую (содержит многие ферментные системы- дыхательные, окислительно- восстановительные, осуществляет перенос электронов), транспортную (перенос различных веществ в клетку и из клетки).
4. Клеточная стенка- присуща большинству бактерий (кроме микоплазм, ахолеплазм и некоторых других не имеющих истинной клеточной стенки микроорганизмов). Она обладает рядом функций, прежде всего обеспечивает механическую защиту и постоянную форму клеток, с ее наличием в значительной степени связаны антигенные свойства бактерий. В составе - два основных слоя, из которых наружный- более пластичный, внутренний- ригидный.
Основное химическое соединение клеточной стенки, которое специфично только для бактерий- пептидогликан (муреиновые кислоты). От структуры и химического состава клеточной стенки бактерий зависит важный для систематики признак бактерий- отношение к окраске по Граму. В соответствии с ним выделяют две большие группы- грамположительные (“грам+”) и грамотрицательные (“грам - “) бактерии. Стенка грамположительных бактерий после окраски по Граму сохраняет комплекс йода с генциановым фиолетовым (окрашены в сине- фиолетовый цвет), грамотрицательные бактерии теряют этот комплекс и соответствующий цвет после обработки и окрашены в розовый цвет за счет докрашивания фуксином.
Особенности клеточной стенки грамположительных бактерий.
Мощная, толстая, несложно организованная клеточная стенка, в составе которой преобладают пептидогликан и тейхоевые кислоты, нет липополисахаридов (ЛПС), часто нет диаминопимелиновой кислоты.
Особенности клеточной стенки грамотрицательных бактерий.
Клеточная стенка значительно тоньше, чем у грамположительных бактерий, содержит ЛПС, липопротеины, фосфолипиды, диаминопимелиновую кислоту. Устроена более сложно- имеется внешняя мембрана, поэтому клеточная стенка трехслойная.
При обработке грамположительных бактерий ферментами, разрушающими пептидогликан, возникают полностью лишенные клеточной стенки структуры- протопласты. Обработка грамотрицательных бактерий лизоцимом разрушает только слой пептидогликана, не разрушая полностью внешней мембраны; такие структуры называют сферопластами. Протопласты и сферопласты имеют сферическую форму (это свойство связано с осмотическим давлением и характерно для всех безклеточных форм бактерий).
L- формы бактерий.
Под действием ряда факторов, неблагоприятно действующих на бактериальную клетку (антибиотики, ферменты, антитела и др.), происходит L- трансформация бактерий, приводящая к постоянной или временной утрате клеточной стенки. L- трансформация является не только формой изменчивости, но и приспособления бактерий к неблагоприятным условиям существования. В результате изменения антигенных свойств (утрата О- и К- антигенов), снижения вирулентности и других факторов L- формы приобретают способность длительно находиться (персистировать) в организме хозяина, поддерживая вяло текущий инфекционный процесс. Утрата клеточной стенки делает L- формы нечувствительными к антибиотикам, антителам и различным химиопрепаратам, точкой приложения которых является бактериальная клеточная стенка. Нестабильные L- формы способны реверсировать в классические (исходные) формы бактерий, имеющие клеточную стенку. Имеются также стабильные L- формы бактерий, отсутствие клеточной стенки и неспособность реверстровать которых в классические формы бактерий закреплены генетически. Они по ряду признаков очень напоминают микоплазмы и другие молликуты - бактерии, у которых клеточная стенка отсутствует как таксономический признак. Микроорганизмы, относящиеся к микоплазмам - самые мелкие прокариоты, не имеют клеточной стенки и как все бактериальные бесстеночные структуры имеют сферическую форму.
К поверхностным структурам бактерий (необязательным, как и клеточная стенка), относятся капсула, жгутики, микроворсинки.
Капсула или слизистый слой окружает оболочку ряда бактерий. Выделяют микрокапсулу, выявляемую при электронной микроскопии в виде слоя микрофибрилл, и макрокапсулу, обнаруживаемую при световой микроскопии. Капсула является защитной структурой (прежде всего от высыхания), у ряда микробов- фактором патогенности, препятствует фагоцитозу, ингибирует первые этапы защитных реакций- распознавание и поглощение. У сапрофитов капсулы образуются во внешней среде, у патогенов- чаще в организме хозяина. Существут ряд методов окраски капсул в зависимости от их химического состава. Капсула чаще состоит из полисахаридов (наиболее распространенная окраска- по Гинсу), реже- из полипептидов.
Жгутики. Подвижные бактерии могут быть скользящие (передвигаются по твердой поверхности в результате волнообразных сокращений) или плавающие, передвигающиеся за счет нитевидных спирально изогнутых белковых (флагеллиновых по химическому составу) образований- жгутиков.
По расположению и количеству жгутиков выделяют ряд форм бактерий.
1.Монотрихи - имеют один полярный жгутик.
2.Лофотрихи - имеют полярно расположенный пучок жгутиков.
3.Амфитрихи - имеют жгутики по диаметрально противоположным полюсам.
4.Перитрихи - имеют жгутики по всему периметру бактериальной клетки.
Способность к целенаправленному движению (хемотаксис, аэротаксис, фототаксис) у бактерий генетически детерминирована.
Фимбрии или реснички - короткие нити, в большом количестве окружающую бактериальную клетку, с помощью которых бактерии прокрепляются к субстратам (например, к поверхности слизистых оболочек). Таким образом, фимбрии являются факторами адгезии и колонизации.
F- пили (фактор фертильности) - аппарат конъюгации бактерий, встречаются в небольшом количестве в виде тонких белковых ворсинок.
Эндоспоры и спорообразование.
Спорообразование- способ сохранения определенных видов бактерий в неблагоприятных условиях среды. Эндоспоры образуются в цитоплазме, представляют собой клетки с низкой метаболической активностью и высокой устойчивостью (резистентностью) к высушиванию, действию химических факторов, высокой температуры и других неблагоплиятных факторов окружающей среды. При световой микроскопии часто используют метод выявления спор по Ожешко. Высокая резистентность связана с большим содержанием кальциевой соли дипиколиновой кислоты в оболочке спор. Расположение и размеры спор у различных микроорганизмов отличается, что имеет дифференциально- диагностическое (таксономическое) значение. Основные фазы “жизненного цикла” спор- споруляция (включает подготовительную стадию, стадию предспоры, образования оболочки, созревания и покоя) и прорастание, заканчивающееся образованием вегетативной формы. Процесс спорообразования генетически обусловлен.
Некультивируемые формы бактерий.
У многих видов грамотрицательных бактерий, не образующих спор, существует особое приспособительное состояние- некультивируемые формы. Они обладают низкой метаболической активностью и активно не размножаются, т.е. не образуют колоний на плотных питательных средах, при посевах не выявляются. Обладают высокой устойчивостью и могут сохранять жизнеспособность в течение нескольких лет. Не выявляются классическими бактериологическими методами, обнаруживаются только при помощи генетических методов ( полимеразной цепной реакции- ПЦР).
Похожие работы
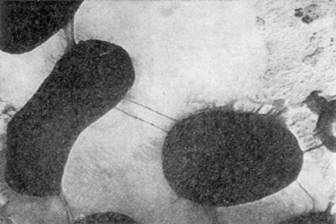
... обнаруживается большое количество типичных ветвящихся клеток. Следовательно, ветвление у микобактерий зависит в значительной степени от питательной среды [5]. 3. Особенности физиологии микроорганизмов рода Mycobacterium Микобактерии характеризуются высоким содержанием липидов (от 30,6 до 38,9 %), вследствие этого трудно окрашиваются анилиновыми красителями, но хорошо воспринимают краску ...
... . ЛИТЕРАТУРА: 1. Курс низших растений //Под ред. М. В. Горленко – М.: Высш. школа, 1981. 2. Жизнь растений, т. 1–3, - М.: Просвещение, 1974-77. 3. Н. Грин, У. Стаут, Д. Тейлор. Биология, т. 1-3. – М.: Мир, 1990. 4. Н.П.Горбунова , Е.С.Клюшникова, Н.А.Комарницкий. и др. Малый практикум по низшим растениям. – М.: Высш. школа, 1976. 5. Русанов В. А. Низшие растения. Методические указания ...
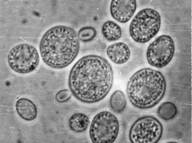
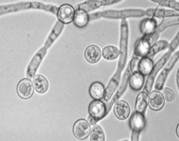
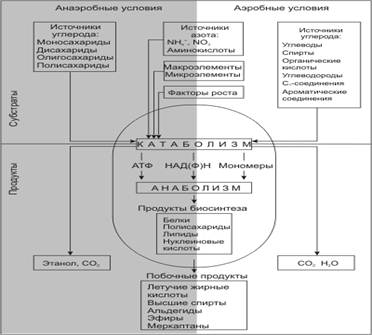
... так как от их состава зависит качество пищевого продукта, получаемого с помощью дрожжей. Такие соединения называют органолептическими. Обязательные побочные продукты метаболизма дрожжей – высшие спирты (сивушные масла). Механизм синтеза высших спиртов связан с образованием алифатических аминокислот. Чаще всего встречаются спирты: пропиловый, изоамиловый, бутиловый, изобутиловый. Летучие жирные ...
... попытку классифицировать растения, привлекая в дополнение к обычному в то время делению растений на деревья, кустарники и травы также признаки цветков, плодов и семян. Швейцарский ботаник Иоганн Баугин (Жан Боэн) в своей «Всеобщей истории растений», опубликованной (1650) после его смерти, описал около 5000 растений. Его брату Каспару Баугину ботаника обязана созданием бинарной номенклатуры, то ...
0 комментариев