Содержание
1. Спорогенез и гаметогенез у растений
2. Норма реакции генотипа
3. Бесплодие отдаленных гибридов, его причины и способы преодоления
Список литературы
1. Спорогенез и гаметогенез у растений
Способность к размножению, или самовоспроизведению, является одним из обязательных и важнейших свойств живых организмов. Размножение поддерживает длительное существование вида, обеспечивает преемственность между родителями и их потомством в ряду многих поколений. Оно приводит к увеличению численности особей вида и способствует его расселению. У растений, подавляющее большинство которых ведет прикрепленный образ жизни, расселение в процессе размножения - единственный способ занять большую территорию обитания.
Различают два типа размножения: бесполое и половое. В, бесполом размножении участвует только одна родительская особь, которая делится, почкуется и образует споры. Размножение при помощи вегетативных органов у растений называется вегетативным. В случае полового размножения особи нового поколения появляются при участии двух организмов - материнского и отцовского.
Вегетативное размножение основано на способности организмов восстанавливать (регенерировать) недостающие части. Этот способ размножения широко распространен в природе, но с наибольшим разнообразием оно осуществляется у растений, особенно у цветковых.
Бесполое размножение характеризуется тем, что для воспроизводства потомства образуются специализированные клетки - споры, каждая из которых прорастает и дает начало новому организму. Спорообразование встречается у простейших (малярийный плазмодий), грибов, водорослей, мхов, плаунов, хвощей и папоротников. У голо - и покрытосеменных растений споры непосредственно в процессе, размножения не участвуют.
Споры образуются путем митоза или мейоза в обычных вегетативных клетках материнского организма или специальных органах - спорангиях и представляют собой микроскопические одноклеточные образования.
При любой форме бесполого размножения - частями тела или спорками - наблюдается увеличение численности особей данного вида без повышения их генетического разнообразия: все особи являются точной копией материнского организма. Эта особенность используется человеком для получения однородного, с хорошими признаками, потомства у плодово-ягодных, декоративных и других групп растений. Новые признаки у таких организмов появляются только в результате мутаций.
Половое размножение существенно отличается от бесполого тем, что в данном случае генотип потомков возникает в результате перекомбинации генов, принадлежащих обоим родителям. Это повышает возможности организмов в приспособлении к меняющимся условиям среды.
Половое размножение характеризуется наличием полового процесса, одним из важнейших этапов которого является слияние половых клеток, или гамет, специализированных гаплоидных клеток, одетых плазматической мембраной. Гаметы различаются по строению и физиологическим свойствам и делятся на мужские (подвижные - сперматозоиды, неподвижные - спермин) и женские (яйцеклетки). В отличие от спор одна гамета, за исключением случаев партеногенеза, не может дать начало новой особи. Этому предшествует процесс слияния двух половых клеток - оплодотворение, в результате которого образуется зигота. В дальнейшем из зиготы развивается зародыш нового организма.
Образование половых клеток (гаметогенез) у водорослей, многих грибов и высших споровых растений происходит путем митоза или мейоза в специальных органах полового размножения: яйцеклеток - в оогониях или архегониях, сперматозоидов и спермиев - в антеридиях. В процессе формирования половых клеток выделяют три стадии - размножения, роста и созревания[1].
Первичные половые клетки делятся путем митоза (период размножения), в результате чего их количество постоянно возрастает. В период роста деление клеток прекращается, и они начинают усиленно расти. При этом будущие яйцеклетки (ооциты) увеличиваются в размерах иногда в сотни и даже в тысячи раз за счет накопления в их цитоплазме запасных питательных веществ в виде желтка. Размеры незрелых мужских гамет (сперматоцитов) увеличиваются незначительно. Затем происходит их мейотическое деление, что приводит к образованию четырех гаплоидных клеток. При сперматогенезе все четыре клетки в дальнейшем превращаются в сперматозоиды.
Половое размножение растений включает несколько физиологических процессов: цветение, опыление, оплодотворение и образование плода и семени.
При оплодотворении пыльца, попадая на рыльце пестика, прорастает, образуя пыльцевую трубку, достигающую семяпочки в завязи цветка. У растений может быть одна семяпочка или несколько. В нижней части пыльцевой трубки образуются спермин. Затем происходит так называемое двойное оплодотворение, при котором один из спермиев сливается с яйцеклеткой, а другой - с центральной клеткой семяпочки. После оплодотворения из яйцеклетки в результате многократного деления развивается зародыш семени, а из оплодотворенной центральной клетки образуется запас питательных веществ семени. Так в результате двойного оплодотворения образуется семя, состоящее из оболочки, зародыша и запаса питательных веществ, из которого впоследствии развивается новое растение. Полученное при половом размножении потомство наследует признаки обоих родителей[2].
2. Норма реакции генотипа
При формировании генетических представлений о связи между геном и признаком изначально предполагалось, что каждому признаку соответствовал особый детерминант (наследственный фактор), который обусловливал развитие своего признака. Однако такие представления далеки от истины, а прямые и однозначные связи гена с признаком на самом деле скорее исключение, чем правило. Было установлено, что на один признак могут влиять многие гены и, наоборот, один ген часто влияет на многие признаки. Кроме того, действие гена может быть изменено соседством других генов или условиями внешней среды.
В онтогенезе действуют скорее не отдельные гены, а весь генотип как целостная интегрированная система со сложными связями и взаимодействиями ее компонентов. Более того, эта система не является застывшей, она динамична, меняется, совершенствуется во времени, в результате генных мутаций постоянно появляются новые гены. Могут формироваться также качественно новые хромосомы за счет хромосомных мутаций и даже новые геномы за счет геномных мутаций. Вновь возникшие гены могут сразу же вступать во взаимодействие с уже имевшимися генами или менять, модифицировать характер работы последних, даже будучи рецессивными, т.е. не проявляясь сами по себе[3].
Таким образом, в каждый конкретный промежуток времени у каждого вида растений и животных генотип проявляет себя как исторически сложившаяся к данному моменту целостная система.
Характер проявления действия гена может изменяться в различных ситуациях и под влиянием различных факторов. Законы Менделя отражают законы наследования, то есть передачи генов в ряду поколений, только при обязательном соблюдении двух условий: гены должны быть локализованы в разных парах гомологичных хромосом (это дает им возможность независимо комбинироваться и наследоваться) и за каждый признак должен отвечать только один ген. Однако это далеко не всегда так. Для того, чтобы убедиться в том, что характер проявления генов разнообразен, рассмотрим свойства генов и особенности их проявления в признаках:
ген дискретен в своем действии, то есть, прерывист, обособлен в своей активности от других генов;
ген специфичен в своем проявлении, т.е. отвечает за строго определенный признак (на молекулярном уровне каждый ген отвечает за синтез одного конкретного белка);
ген может действовать градуально, то есть может усиливать степень проявления признака (например, увеличивать количество синтезируемого вещества) при увеличении числа доминантных аллелей (дозы гена);
один ген может влиять на развитие разных признаков - это множественное, или плейотропное, действие гена;
разные гены могут оказывать одинаковое действие на развитие одного и того же признака - это множественные гены, или полигены; при этом чаще всего наблюдается усиление или ослабление признаков - в таком случае это кумулятивное (накопительное) действие гена, которое обусловливает проявление так называемых количественных признаков;
ген может вступать во взаимодействие с другими генами, что приводит к появлению новых признаков. Поскольку гены дискретны и специфичны, они взаимодействуют не непосредственно, а продуктами своих реакций - веществами, синтезированными под их контролем;
действие гена может быть модифицировано изменением его местоположения в хромосоме (эффект положения) или условиями внешней среды и другими факторами[4].
Множественное действие генов - это способность гена воздействовать на несколько признаков одновременно.
В процессе индивидуального развития организма фенотип может меняться, а генотип остается таким же, каким был получен от родителей при слиянии их гамет (процесс мутирования в данном случае во внимание не принимается). Как правило, роль генотипа в определении фенотипа является решающей. Это относится в первую очередь к проявлению ряда качественных признаков (красная окраска цветков, желтая и зеленая окраска семян гороха, голубой цвет глаз у человека, наличие ушной раковины и т.д.), а также к большинству простых биохимических признаков (синтез определенных специфических белков при наличии всех необходимых компонентов).
Однако роль условий внешней среды в реализации многих или даже большинства признаков игнорировать нельзя. Они могут модифицировать, то есть изменять, характер проявления признака, но только в определенных, наследственно обусловленных пределах, называемых нормой реакции. Такая изменчивость признака в одну и в другую сторону под влиянием условий внешней среды называется модификационной. Она не наследуется, а проявляется только в индивидуальном развитии данного организма. Влиянию условий внешней среды в большей степени подвержены количественные признаки. Поэтому по фенотипу часто невозможно определить, является ли он следствием только генотипа или генотипа и условий среды. Для того чтобы представить всю сложность взаимодействия генотипа с условиями среды, а также определить долю вклада генотипа и условий среды в фенотипическое проявление признака, обычно применяются специальные методы математического анализа.
3. Бесплодие отдаленных гибридов, его причины и способы преодоления
Так как одним из методов селекции является гибридизация, то большую роль играет выбор типа скрещиваний, т.е. система скрещиваний.
Системы скрещивания могут быть разделены на два основных типа: близкородственное (инбридинг - разведение в себе) и скрещивание между неродственными формами (аутбридинг - неродственное разведение). Если принудительное самоопыление приводит к гомозиготизации, то неродственные скрещивания - к гетерозиготизации потомков от этих скрещиваний.
Инбридинг, то есть принудительное самоопыление перекрестноопыляющихся форм, кроме прогрессирующей с каждым поколением степени гомозиготности, приводит и к распадению, разложению исходной формы на ряд чистых линий. Такие чистые линии будут обладать пониженной жизнеспособностью, что, по-видимому, связано с переходом из генетического груза в гомозиготное состояние всех рецессивных мутаций, которые в. основном являются вредными.
Чистые линии, полученные в результате инбридинга, имеют различные свойства. У них различные признаки проявляются по-разному. Кроме того, различна и степень снижения жизнеспособности. Если эти чистые линии скрещивать между собой, то, как правило, наблюдается эффект гетерозиса.
Гетерозис - явление повышенной жизнеспособности, урожайности, плодовитости гибридов первого поколения, превышающих по этим параметрам обоих родителей. Уже со второго поколения гетерозисный эффект угасает. Генетические основы гетерозиса не имеют однозначного толкования, но предполагается, что гетерозис связан с высоким уровнем гетерозиготности у гибридов чистых линий (межлинейные гибриды). Производство чистолинейного материала кукурузы с использованием так называемой цитоплазм этической мужской стерильности было широко изучено и поставлено на промышленную основу в США. Ее использование исключало необходимость кастрировать цветки, удалять пыльники, так как мужские цветки растений, используемые в качестве женских, были стерильны.
Разные чистые линии обладают разной комбинационной способностью, то есть дают неодинаковый уровень гетерозиса при скрещиваниях друг с другом. Поэтому, создав большое количество чистых линий, экспериментально определяют наилучшие комбинации скрещиваний, которые затем используются в производстве.
Отдаленная гибридизация - это скрещивание растений, относящихся к различным видам. Отдаленные гибриды, как правило, стерильны, что связано с содержанием в геноме различных хромосом, которые в мейозе не конъюгируют. В результате этого формируются стерильные гаметы. Для устранения данной причины в 1924 г. советским ученым Г.Д. Карпеченко было предложено использовать удвоение числа хромосом у отдаленных гибридов, которое приводит к образованию амфидиплоидов по следующей схеме[5]:
Р: пшеница Х рожь
Р: пшеница Х рожь
2n = 42 2n = 14
(21 пара) (7 пар)
G: n = 21 n = 7
F1: n = 28 (все непарные)
G: Мейоз нарушен, гибрид стерилен, нормальных гамет нет.
Обработка колхицином, приводящая к удвоению числа хромосом.
F1 (колхицированное): 2n = 56
(28 пар)
G: n = 28
F2, F3...Fn 2n = 56
тритикале
Таким методом кроме тритикале были получены многие ценные отдаленные гибриды, в частности многолетние пшенично-пырейные гибриды и др. У таких гибридов в клетках содержится полный диплоидный набор хромосом одного и другого родителя, поэтому хромосомы каждого родителя конъюгируют друг с другом и мейоз проходит нормально. Путем скрещивания с последующим удвоением числа хромосом терна и алычи удалось повторить эволюцию - произвести ресинтез вида сливы домашней.
Подобная гибридизация позволяет полностью совместить в одном виде не только хромосомы, но и свойства исходных видов. Например, тритикале сочетает многие качества пшеницы (высокие хлебопекарные качества) и ржи (высокое содержание незаменимой аминокислоты лизина, а также способность расти на бедных песчаных почвах).
Это один из примеров использования в селекции полиплоидии, точнее аллоплоидии. Еще более широко используется автополиплоидия.
При использовании новых методов селекции получены новые сорта растений. Так, академиком Н.В. Цициным путем отдаленной гибридизации пшеницы с пыреем и последующей полиплоидизации выведены многолетние пшеницы. Такими же методами получены перспективные сорта новой зерновой культуры тритикале. Для селекции вегетативно размножаемых растений используются соматические мутации (они использовались и И.В. Мичуриным, но он называл их почковыми вариациями). Широкое применение получили многие методы И.В. Мичурина после их генетического осмысления, хотя некоторые из них теоретически так и не разработаны. Большие успехи достигнуты в использовании результатов мутационной селекции в выведении новых сортов зерновых, хлопчатника и кормовых культур. Однако наибольший вклад во все возделываемые сорта внесли образцы коллекции мирового генофонда культурных растений, собранные Н.И. Вавиловым и его учениками.
Отдаленная гибридизация домашних животных менее продуктивна, чем у растений, так как преодолеть стерильность отдаленных гибридов невозможно, если она проявляется. Правда, в некоторых случаях отдаленная гибридизация видов с родственными хромосомными наборами не приводит к нарушению мейоза, а ведет к нормальному слиянию гамет и развитию зародыша у отдаленных гибридов, что позволило получить некоторые ценные породы, сочетающие полезные признаки обоих использованных в гибридизации видов. Успешно завершились попытки улучшить породы местного крупного рогатого скота скрещиванием его с зебу и яками.
Следует отметить, что не всегда необходимо добиваться плодовитого потомства от отдаленной гибридизации. Иногда полезны и стерильные гибриды, как, например, веками использующиеся мулы - стерильные гибриды лошади и осла, отличающиеся выносливостью и долговечностью.
Задача 1.
У пшеницы карликовость доминировала над высокорослостью. В скрещиваниях получены расщепления по фенотипу 3: 1 и 1:
1. Определите генотипы и фенотипы родителей.
Ответ:
Расщепление по фенотипу 3: 1 говорит о моногибридном скрещивании, следовательно, можно сделать вывод, что у обоих родителей доминировала карликовость, то есть фенотип родителей такой же как и фенотип первого поколения. Во втором поколении гибридов от скрещиваемых наблюдается расщепление на исходные родительские признаки в отношении: 3 части доминантных и одна часть рецессивных.
Расщепление по первому гену 1: 1 говорит о том, что одна родительская форма является гомозиготной, а другая гетерозиготной.
Задача 2.
От скрещивания устойчивого к головне фуркатного ячменя с восприимчивым к головне остистым ячменем получили гибриды F1 устойчивые к головне с фуркатным колосом. Что ожидают по фенотипу и генотипу в анализирующем скрещивании, если наследование признаков независимое?
Ответ:
В случае полного доминирования по внешнему проявлению признака (по фенотипу) судить о генотипе организма невозможно, так как и доминантная гомозигота (ААВВ) и гетерозигота (АаВв) будут обладать фенотипически доминантными признаками, в данном случае устойчивость к головне с фуркатным колосом. Для того, чтобы отличить доминантную гомозиготу от гетерозиготы, применяется анализирующее скрещивание, т.е. скрещивание гетерозиготы с рецессивной гомозиготой.
Анализирующее скрещивание позволяет определить генотип особей с доминантным фенотипом. Это возможно при скрещивании с рецессивной гомозиготой по всем анализируемым генам, причем независимо от их числа. При таком скрещивании каждый аллель анализируемой формы проявляется на фоне второго рецессивного аллеля: доминантный потому, что подавляет действие рецессивного, а рецессивный потому, что находится в состоянии рецессивной гомозиготы. Поскольку проявляется каждый аллель, получим расщепление 1: 1 по каждому гетерозиготному фактору и не получим по гомозиготным.
Задача 3.
У кукурузы растения нормальной высоты имеют в своем генотипе два неаллельных доминантных гена. Гомозиготность по рецессивным аллелям даже одного из этих генов приводит к возникновению карликовых форм. От скрещивания двух карликовых растений получили гибриды F1 нормальной высоты. Какие результаты по генотипу и фенотипу ожидаются от самоопыления растений F1 в потомстве F2?
Ответ:
В потомстве F2 после самоопыления будет наблюдаться расщепление. Карликовую форму имеют только растения, несущие доминантные аллели обоих генов. Карликовость растений возможна только при наличие обоих доминантных аллелей.
Задача 4.
Одна из цепочек ДНК имеет следующее чередование нуклеотидов: Г-Т-А-А-Т-Г-Ц-Ц-Т-Г-Ц-Ц... Укажите схему транскрипции и трансляции генетической информации с данного участка ДНК.
Ответ:
Транскрипция происходит не на всей молекуле ДНК, а лишь на небольшом ее участке, соответствующем определенному гену. Она может протекать одновременно на нескольких генах одной хромосомы и на генах, расположенных на разных хромосомах. В результате транскрипции образуется иРНК, последовательность нуклеотидов которой является точной копией последовательности нуклеотидов матрицы - одного или группы рядом расположенных генов. Так, если в молекуле ДНК имеется азотистое основание - цитозин (Ц), то в РНК - гуанин (Г), и наоборот. В ДНК комплементарной парой является аденин - тимин. Однако в состав РНК вместо тимина входит урацил.
Трансляция начинается со стартового кодона АУГ. Отсюда каждая рибосома прерывисто, триплет за триплетом движется вдоль молекулы иРНК, что сопровождается ростом полипептидной цепочки.
Выстраивание аминокислот в соответствии с кодонами иРНК осуществляется на рибосомах при помощи транспортной РНК. Благодаря определенному расположению комплементарных нуклеотидов цепочка тРНК имеет форму, напоминающую лист клевера. При реализации генетической информации каждая тРНК присоединяет и переносит к месту синтеза белка соответствующую аминокислоту.
Список литературы
1. Алтухов Ю.П. Генетические процессы в популяциях. - М.: Мир, 1983.
2. Биология. / Сост. Лемеза Н.А., Морозик М.С., Морозов Е.И. - Минск: Университетское, 1999.
3. Дубинин Н.П. Общая генетика. - М.: Колос, 1987.
4. Замотайлов С.С., Бурдун А.М. Краткий курс генетики. - М.: Агропромиздат, 1987.
5. Основы генетики и селекции. / Под ред. Дубникова А.С. - М.: Колос, 1997.
[1] Основы генетики и селекции. / Под ред. Дубникова А.С. - М.: Колос, 1997.
[2] Алтухов Ю. П. Генетические процессы в популяциях. - М.: Мир, 1983.
[3] Замотайлов С. С., Бурдун А. М. Краткий курс генетики. - М.: Агропромиздат, 1987.
[4] Дубинин Н. П. Общая генетика. - М.: Колос, 1987.
[5] Биология. / Сост. Лемеза Н.А., Морозик М.С., Морозов Е.И. – Минск: Университетское, 1999. – с.-142.
Похожие работы
... фазы развития. Кроме того, применение указанных гаметоцидов задерживало прохождение фенологических фаз у растений, а также выколашивание части стеблей. Однако, несмотря на отрицательные побочные действия гаметоцидов, применение их в ближайшей перспективе может открыть путь к реализации эффекта гетерозиса у зерновых (особенно пшеницы), технических, овощных и кормовых культур. Поэтому весьма ...
... . ЛИТЕРАТУРА: 1. Курс низших растений //Под ред. М. В. Горленко – М.: Высш. школа, 1981. 2. Жизнь растений, т. 1–3, - М.: Просвещение, 1974-77. 3. Н. Грин, У. Стаут, Д. Тейлор. Биология, т. 1-3. – М.: Мир, 1990. 4. Н.П.Горбунова , Е.С.Клюшникова, Н.А.Комарницкий. и др. Малый практикум по низшим растениям. – М.: Высш. школа, 1976. 5. Русанов В. А. Низшие растения. Методические указания ...
... название "волшебных". Тот факт, что белки всех организмов построены из одних и тех же аминокислот - еще одно доказательство единства живого мира на Земле. В строении молекул белков различают 4 уровня организации: 1. Первичная структура - полипептидная цепь из аминокислот, связанных в определенной последовательности ковалентными пептидными связями. 2. Вторичная структура - полипептидная цепь в ...
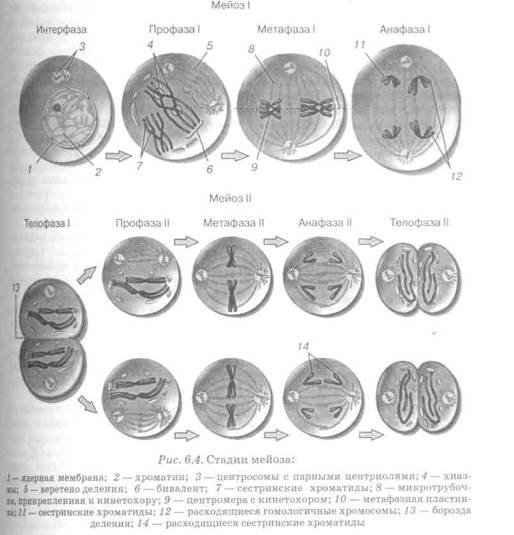
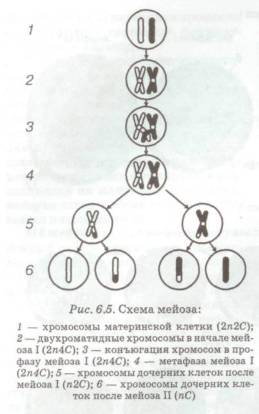
... яйцеклеток и сперматозоидов количество хромосом увеличивалось бы в два раза. Мейоз служит также основой комбинативной изменчивости. мейоз деление клетка пищеварительная система 2 (28) Строение и значение пищеварительной системы Пищеварительная система представляет собой комплекс органов, осуществляющих процесс механической и химической обработки пищи, всасывание переработанных веществ и ...
0 комментариев