Навигация
Гаметоциды и их применение в селекции
126356
знаков
6
таблиц
1
изображение
Гаметоциды — химические соединения, применение .которых в определенные этапы развития растений приводит к гибели мужского гаметофи-та в результате нарушении метаболических процессов в период формирования пыльцевых зерен. Действие гаметоцидов на физиолого-биохимиче-ские процессы в клетке аналогично стерилизующей цитоплазме при перемещении в нее ядра, что вызывает мужскую стерильность.
Исследования, проведенные в ряде стран, в том числе и в СССР, позволили выявить ряд химических соединений, которые индуцируют мужскую стерильность у яровой и озимой пшеницы, ржи и других сельскохозяйственных культур. Использование гаметоцидов позволяет организовать Промышленное .производство гибридных семян зерновых и других сельскохозяйственных культур. Однако полученные гаметоциды вызывают ряд побочных нежелательных явлений: значительное ингибирование роста, задержку 'в прохождении .фенологических фаз и д'р. Поэтому необходим дальнейший поиск новых химических соединений, обладающих гаметоцид-ной активностью.
.
ВВЕДЕНИЕ
|Рост народонаселения планеты, необходимость повышения темпов производства продуктов сельского хозяйства определяют поиск новых направлений в селекции сортов и гибридов — основных средств сельскохозяйственного производства. В последнее время широкие исследования гетерозиса пшеницы, ржи, ячменя, подсолнечника, сахарной свеклы, хлопчатника, ряда овощных, бахчевых, цветочных, декоративных культур и кормовых трав показали, что гибриды первого поколения этих растений обеспечивают прибавку урожая на 20—25% и более по сравнению с районированными сортами [1,7, 35, 36].
Практическое использование гетерозисного эффекта возможно лишь при условии хорошо налаженного производства гибридные семян в значительном объеме. У ряда культур, обладающих высоким коэффициентом размножения семян и низкой посевной нормой (кукуруза, томаты и др.), производство гибридных семян может быть организовано при ручной кастрации материнских растений с последующим свободным или принудительным опылением. Однако для большинства возделываемых растений этот метод неприменим, так как затраты труда, необходимые для получения гибридных семян первого поколения, не окупаются дополнительно получаемой продукцией.
В настоящее время в результате использования явления цитоплазматической мужской стерильности (ЦМС) получены гибриды первого поколения пшеницы, сорго, кукурузы, сахарной свеклы, томатов, лука, сладкого перца и налажено массовое производство гибридных семян. Близки к завершению исследования по созданию гибридов ржи и подсолнечника [35]. Наряду с этим при селекции с томатами пытаются применить формы с функциональной мужской стерильностью [36]. 'Создание мужских стерильных линий, отбор линий — закрепителей стерильности и форм, восстанавливающих фер-тильность, значительно усложнили ведение селекции я семеноводства. Между тем основным недостатком использования системы «ЦМС — восстановление фертильноста» при получении новых гибридных комбинаций скрещиваний является постоянная необходимость создания стерильных аналогов и аналогов '— восстановителей фертильности новых высокопродуктивных сортов и самоопыленных линий.
Сравнительно недавно для индукции мужской стерильности на растения в определенные фазы развития воздействовали различными физическими факторами (сокращенный световой день, высокие и низкие температуры и др.) [2, 48, 97, 130]. Несмотря на полученные положительные результаты» ввиду отсутствия технических средств для массового производства гибридных семян эти приемы не были внедрены в практику.
•Определенные успехи в разработке методов мужской стерильности растений были достигнуты с помощью ряда химических соединений — растворов солей феноксиуксусной кислоты, гидразида малеиновой кислоты (ГМ1К), этрела, индо-лилуксусной кислоты (ИУК), нафтнлуксусной кислоты (НУК), гиббереллина (ГКз) и некоторых других, названных гаметоцидами. Этот термин — производное от двух слов:
гамета — по-гречески половая клетка, цидо — по-латински убивать. В настоящее время в ряде стран Европы и в США проводят исследования по выявлению химических соединений, обладающих гаметоцядной активностью к мужскому гамето-фиту растений, и разрабатывают способы их применения для массового производства гибридных семян различных сельскохозяйственных культур. Использование методов химической индукции мужской стерильности у зерновых, технических, овощных и кормовых культур позволит получить семена гибридов первого поколения, вовлекая в селекционный процесс наиболее совершенные сорта без создания их стерильных аналогов и линий, несущих гены восстановления фертильности.
Преодоление трудностей, препятствующих эффективному использованию гаметоцидов, в значительной мере зависит от определения сущности процесса стерилизации, в основе которого лежат конкретные физиолого-биохимические изменения в тканевом метаболизме спорогенеза, вызывающие анормальности в развитии пыльцевого зерна с последующей его абортивностью. В связи с практическими запросами физиология развития пыльцы привлекает особое внимание.
ЦИТОЛОГИЧЕСКИЕ, ЦИТОХИМИЧЕСКИЕ И ФИЗИОЛОГО-БИОХИМИЧЕСКИЕ ИССЛЕДОВАНИЯ ФОРМИРОВАНИЯ МУЖСКОГО ГАМЕТОФИТА
Мужской гаметофит покрытосеменных растений — микроскопическая структура, которая в период ранних стадий развития окружена тканями пыльника и непосредственно контактирует с клетками тапетума, метаболизм которых тесно связан с развитием пыльцевьгх зерен, особенно в критические периоды преобразования материнских клеток пыльцы в гап-лоидные микроспоры. Так как наибольшая эффективность действия гаметоцидов приходится на период формирования тетрад и связана с биохимизмом их развития, возникает необходимость в детальной информации о ранних фазах развития пыльцевого зерна в материнской клетке пыльцы.
С помощью электронной микроскопии установлено, что до мейоза материнские клетки пыльцы в пыльниках связаны между собой и с клетками тапетума цитоплазматлческими мостиками (плазмодесмами) [5, 59]. Перед стадией лептотены профазы 1 начинаются преобразовательные процессы в оболочках и мембранах микроспороцитов, которые быстро обволакиваются слоем каллозы, представляющей собой (3-1,3-свя-занный полимер глюкозы [130]. Это соединение локализуется между оболочками материнской клетки пыльцы и клеточной мембраной. С началом стадии лептотены плазмодесмы, соединяющие клетки тапетума и микроспороцитов, исчезают. Слой каллозы продолжает утолщаться, однако некоторые районы оболочек микроспороцитов не покрываются каллозой, здесь соседние материнские клетки пыльцы контактируют друг с, другом с помощью широких цитоплазматичесиих перетяжек, названных цитомиксическими каналами [28]. У большинства видов растений все микроспороциты мейотической профазы взаимосвязаны. Такая цитоплазматическая общность — важнейший фактор поддержания тесной синхронности мейотических стадий. Цитомиксические каналы начинают исчезать к концу мейотической профазы I, а в телофазе I их количество совсем незначительно, к началу же профазы II материнские клетки пыльцы полностью изолируются друг от друга каллозными оболочками. После мейоза каллоза образует между четырьмя гаплоидными спорами тонкую прослойку (стадия тетрад), и каждая спора становится полностью изолированной от других. Предполагают, что в формирование каллозных оболочек вовлечен аппарат Гольджи [137].
Процесс формирования оболочки пыльцевого зерна начинается вскоре после завершения мейоза и протекает между клеточной мембраной я каллозной оболочкой в каждой споре тетрады. Первичным материалом для построения оболочки является примэкзина. После развития прдмэкзины синтезируется предшественник зрелой экзины, морфология которого .зависит от его химического состава. По своим свойствам это вещество сходно с протоспоруллином. На данном этапе кал-лозная оболочка исчезает, споры быстро увеличиваются в размерах (в 3—5 раз) и интенсивно синтезируют спорополле-нин [70, 71]. После освобождения из тетрад в микроспорах формируется интина, содержащая микрофибряллы целлюло-5
зы, которые образуют матрикс с пектиновым материалом, ге-мицеллюлозой и незначительным количеством белка. До полного созревания пыльцевое зерно покрывается различными производными распадающегося тапетума — каротиноидами, липидами, белками и другими соединениями [130]. При дальнейших фазах развития пыльцы эффективность действия га-метоцидов снижается [47, 65, 75, 80, 102].
Ранние этапы развития пыльцы тесно связаны с функциональной активностью тапетума. Цитохимические и ультраструктурные исследования свидетельствуют, что тапетум функционирует как источник питательного материала при развитии пыльцевого зерна. У покрытосеменных установлено два основных типа тапетума: гландулярный, при котором клетки по мере развития пыльцевых зерен дегенерируют до П9лного автолиза, и амебоидный [76]. В последнем клетки теряют свою оболочку, и протопласт тапетума распределяется между развивающимися пыльцевыми зернами, образуя тапетальный периплазмодий [61]. Мембраны такого тапетума могут проникать в экзину пыльцы. Питательный материал транспортируется из клеток та'петума (возможно, предварительно метабо-ли'аируется им) к пыльцевым зернам .[62]. Взаимодействие тапетума и развивающихся микроопор максимально проявляется при растворении каллозных оболочек микроспор. Исчезновение каллозы сопровождается развитием каллазной активности. Энзим локализован в везикулах цитоплазмы тапетума [130].
Исследование (3-1,3-глюканазной активности в течение мей-оза я распада тетрад показало, что активность энзима резко возрастала, начиная со II мейотдческого деления, достигала максимума в фазу освобождения микроспор из тетрад и сохранялась затем в течение 48 ч [82, 131]. Доказательством взаимосвязи между развивающимися пыльцевыми зернами и тапетумом, а также подтверждением кардинальной роли тапетума в образовании пыльцы служат работы по ЦМС [59, Gl, 62, 76].
В связи с тем, что применение гаметоцидов имеет целью индукцию мужской стерильности растений, для выяснения предполагаемых механизмов действия веществ с гаметоцид-'ной активностью необходима сравнительная оценка и анализ морфологических, цитологических и физиолого-биохимиче-ских различий фертильных растений и линий с ЦМС. Сравнительная анатомия и биохимия тканей пыльников нормальных и стерильных растений свидетельствуют, что абортивность развивающихся пыльцевь»х зерен является результатом включения или выключения более чем одного механизма, а также нарушений регуляторных процессов на различных уровнях 6
контролирования метаболических путей [62, 94, 98, 111]. Отмечено, что у всех изученных линий с ЦМС 90% растений обла^-дали нормальным ходом микроспорогенеза до определенной фазы, на которой начиналась абортивность пыльцы, и только у 10% растений анормальность проявлялась еще до стадии, обусловливающей абортивность в результате более ранних нарушений нормальных метаболических циклов. У большинства видов (около 70%) деградация микроспор приходилась на период, начиная с мейоза 1 и включая интерфазу микроспор [94].
Одной из причин, вызывающих ЦМС растений, являются функциональные нарушения в метаболизме тапетума. Тщательное изучение этой ткани у фертильных и стерильных линий, а также у восстановителей фертильности сорго показало, что у фертильных линий после мейотической профазы I тапетум быстро уменьшался в радиальной протяженности [52]. В стерильных пыльниках радиально расширенный тапетум обусловливает абортивность микроспор, и период его разрастания приходится на стадию вакуолизации микроспоры. Тапетум стерильных пыльников по протяженности (в ширине) и морфологии имеет более широкую шкалу изменчивости, чем у фертильных линий. 'Размеры ширины тапетума стерильных пыльников связывают с дегенеративными процессами в микроспорах [62, 109, 111].
•При цитологических исследованиях клонов ежи сборной с ЦМС не было отмечено отклонений в развитии пыльника в течение архиспориальной стадии и стадии материнской клетки пыльцы по сравнению с нормальными растениями [94, 117]. Первые признаки различий в развитии появлялись в начальные фазы мейоза, что выражалось в разрушении некоторых микроспороцитов, в результате чего образовывалось малое количество тетрад у растений с ЦМС. Одновременно у стерильных растений наблюдалось более быстрое дегенерирова-ние тапетума, чем у фертильных. При сравнительном исследовании микроспорогенеза и развития пыльцы у фертильных и стерильных растений льна установлено, что стерильность пыльцы у этого растения достигалась анормальным разрастанием клеток тапетума, которые медленно дегенерировали по сравнению с тапетумом фертильных растений, у которых величина этой ткани быстро уменьшалась, особенно после стадии тетрад. Очевидно, преждевременное или позднее включение деградативньгх механизмов тапетума может быть причиной функциональной мужской стерильности. У разных видов растений эти механизмы имеют свой специфический характер, что необходимо учитывать при подборе гаметоцидов, их доз и сроков обработки.
• Цитологические исследования пшеницы показали, что на клеточном уровне ЦМС выражается в уменьшении размеров и числа органелл, характеризующихся низкой физиологической активностью и довольно быстрым дегенерированием [59]. У фертильных пыльцевых зерен органеллы клеток обильны.
Различный уровень метаболической активности тапетума в нормальных и стерильных пыльниках и продолжительность его функционирования контролируется цитологами реакцией Фёльгена. Ядра клеток тапетума нормальных пыльников интенсивно окрашиваются до формирования материнской микроспоры, после чего окраска прогрессивно уменьшается по мере коллапсирования тапетума. В тканях тапетума стерильных пыльников окрашивание ядер продолжает осуществляться интенсивно после формирования тетрад и незначительно снижается в процессе развития микроспор [94]. Следовательно, функция тапетума неразрывно связана с его определенной ролью при абортивности микроспор. У нормальных пыльников тапетум получает необходимый питательный материал, поставляемый растением, и преобразует его для формирующихся микроспор, что подтверждается поступлением Фёль-ген-положительных веществ в материнскую микроспору во время мейоза [93]. Тапетум стерильных пыльников имеет более низкий уровень метаболической активности, о чем свидетельствует как низкая энзиматическая активность, так и изменение поступления Фёльген-положительного материала в спорогенные клетки в критические периоды их формирования.
В процессе 'химической индукции мужской стерильности может происходить включение тех же механизмов, что и пр.и абортивности мужского гаметофита стерильных аналогов, где стерильность обусловлена ЦМС. Обработка растений кориандра, вики и лука 0,01%-ным раствором ГМК вызывала 100%-ную стерильность у всех трех растений [60, 89, 90]. После нанесения гаметоцида на растения кориандра тапетум увеличивался и существовал вплоть до стадии вакуолизации микроспор. У контрольных растений тапетум дегенерировал после мейоза. У лука Г1МК, включался в процессы формирования пыльцы, способствуя преждевременному дегенерирова-нию спорогенной ткани и материнских клеток пыльцы. У кориандра химическое индуцирование мужской стерильности проявлялось цитологически — размер ядер не изменялся, а клетки тапетума значительно удлинялись.
Предполагают, что вмешательство различных факторов в метаболизм клеток тапетума приводит к деградации и'х содержимого и, в первую очередь, дезокоирибозидов — основ-
8
ного синтетического пула ДйК, необходимого для нормального развития материнской клетки пыльцы и микроспор. Нарушение деятельности тапетума при снабжеаии питательным ^материалом спорогенной ткани, вызванное различными причинами, обусловливает начало процессов деградации материнской клетки пыльцы [134, 137].
Цитологические исследования пыльников растений с аук-син-индуцированной стерильностью, выращенных при продолжительном освещении, показали, что химические и физические факторы могут способствовать преждевременному де-генерированию тапетума с последующим снижением нормального передвижения питательного материала к спорогенной ткани. В процессе нормального развития пыльцы в начальных (самых ранних) этапах микроспорогенеза ткани тапетума разрастаются и только после завершения формирования микроспор в материнской клетке пыльцы дегенерируют. При гормональной индукции мужской стерильности гиббереллином (ГКз) и в условиях длительного естественного освещения
•отмечена обратная корреляция во взаимодействии микроспор .и тапетума. Большинство микроспор становятся крупными, сильно вакуолизированньши, с ядром, прижатым к клеточной .стенке. Клетки тапетума образуют плотную оболочку вокруг
•полости пыльцевого мешка. Часть микроспор дегенерирует до вакуолизации, но большее их количество абортируется при цветении [94].
Аберрация микроспорогенеза в результате продленного дня, по-видимому, связана с ранним распадом клеток тапетума. Подобный эффект может быть достигнут путем обработки ауксином, высокая концентрация которого обнаружена в тканях растений при длительном фотопериоде [78]. Это подтверждает гипотезу о влиянии фотопериода на пол путем регуляции метаболизма ауксинов в растительных тканях [84, 88].
Считают также, что нарушение функций тапетума в питании развивающихся микроспор не является непосредственной и единственной причиной абортивности пыльцы [46, 107].
При сравнительных исследованиях пыльцы и развития
•пыльников линий пшеницы с ЦМС и нормальных растений установлено, что деградативные процессы в развивающихся пыльцевых зернах начинаются с несбалансированного поступления питательных веществ в тапетум в результате нарушений функциональной активности васкулярной системы тычиночных нитей [31, 32, 87]. Последние у стерильных растений имели слабо дифференцированные васкулярные тяжи (либо они отсутствовали), в то время как у нормальных тычинок сосудистые элементы были хорошо дифференцированы на
9
ксилему и флоему. Наблюдаемое снижение аккумулирования крахмала в тканях тапетума и отсутствие запасного крахмала в зрелых микроспорах является следствием редукции передвижения растворов (в частности углеводов) в тычинки стерильных растений [87]. Нарушение структуры васкулярных элементов при воздействии веществ с гаметоцидной активностью может быть одним из аспектов их действия, тем более, что обработка гаметоци.д'а'ми, как правило, сопровождается уменьшением размеров пыльников и длины тычиночных нитей [20, 59].
Преждевременное деградирование тапетума 'или. более длительное существование интактных клеток тапетума в пыльниках с мужской стерильностью, индуцированной химическими препаратами, по-видимому, является результатом взаимосвязи химически активных веществ с деградативными энзимами (несвоевременная их индукция или блокирование) [56, 60, 74].
При исследовании причин абортивности пыльцы установлено повышение активности кислых фосфатаз в тканях нормальных пыльников до стадии тетрад, после которой активность энзимтав резко снижалась с последующим дегенерирова-нием тапетума [56]. В тканях стерильных пыльников активность энзимов была гораздо ниже, и набухший тапетум раздавливал микроспоры [82].
.Подобный дисбаланс в энзиматической системе стерильных пыльников был отмечен S. Izhar и R. Frankel [82] при сравнительном изучении каллазной активности. В связи с тем, что период развития каллазной активности четко установлен во времени и имеет определенный оптимум кислотности (р.Н 5), авторы измерили in vivo pH различных стадий микроспорогенеза. У фертильных пыльников вплоть до поздней стадии тетрад р.Н составляла около 7, затем снижалась до 6, что сопровождалось растворением каллозы. В пыльниках растений линий с ЦМС pH обычно или низкая — тогда энзиматический распад каллозы материнской клетки пыльцы начинается раньше нормальных сроков, или высокая — тогда наблюдается сильная активность энзима, начиная с профазы 1 в течение всего мейоза, вызывая цитологические нарушения. Преждевременное повышение каллазной активности может быть причиной немедленного разрушения материнской клетки пыльцы. Торможение энэиматической активности кал-лазы блокирует распад каллозной оболочки материнских клеток пыльцы, в результате микроспоры теряют свои нормальные очертания и деградируют.
Эти исследования позволили выявить новые причины абортивности пыльцы и показали, что в основе индукции мужской
10
стерильности лежит дисбаланс в четко скоординированные биохимических реакциях и нарушения в последовательности -„.нзиматических циклов. Изменение четкого ритма энзиматической активности каллазы приводит к деградации развивающихся микроспор [2, 3, 4]. Экспериментально установлено, что период формирования тетрад является наиболее эффективным для применения препаратов с гаметоцидной активностью.
.Возможное участие гаметоцидов в процессах, обусловливающих торможение или .преждевременное освобождение микроспор из материнской клетки пыльцы, вероятно, связано с влиянием соединений с гаметоцндными свойствами на уровни энзиматической активности каллазы. Своевременная индукция и развитие каллазной активности — критический период для нормального развития мужского гаметофита.
Для успешных поисков новых веществ, обладающих гаметоцидной активностью, необходимо располагать достаточной информацией о метаболизме спорогенной ткани и тапетума, а также о взаимосвязи вх обмена. Электронно-микроскопические наблюдения показали, что при нормальной метаболической взаимосвязи между клетками тапетума и развивающимися пыльцевыми зернами не наблюдается никаких аномалий [78]. Согласно имеющимся данным трудно определить, тапетум или спороциты являются 'инициатором -в нарушении взаимосвязи.
Одним из возможных путей выяснения механизма индукции мужской стерильности является исследование биохимической активности тканей. При исследовании пыльников и экстр-актов листьев пшеницы установлено, что мужская сте' рильность пыльцы сопровождается репрессией активности терминальных оксидаз. Уровень активности цитохромоксида-зы в тканях фертильных растений по сравнению со стерильными гораздо выше [18, 27]. Подобное явление отмечено в тканях растений с генетической мужской стерильностью [46]. Цитохромоксидаза — неотъемлемый компонент мито-хондрий, поэтому низкую активность дыхательных энзимов растений с ЦМС можно объяснить дисбалансом митохондри-альных энзиматических систем [128].
Митохондрии, выделенные из стерильного материала (проростки пшеницы с ЦМС), характеризовались более высокой окислительной фосфорилазной активностью, чем мито-хондрии из проростков с нормальной цитоплазмой [129]. Биохимические исследования и наблюдения электронной микроскопии подтвердили, что мужская стерильность тесно связана с изменением функционального состояния этих структур клетки {15, 16, 63]. Восстановление фертильности в линиях
11
с ЦМ'С сопровождается одновременным устранением отклонений в структуре цитоплазматических органелл и различных нарушений в метаболических процессах [27, 46]. Получены дополнительные доказательства в подтверждение гипотезы, что изменения в метаболизме митохондрий связаны с механизмами индукции мужской стерильности. Предполагается, что включение последних вызывает репрессию синтеза белков внутренней мембраны митохондрий [63, 114].
Однако было бы ошибочным считать, что только функциональная активность митохондрий является основой возникновения цитоплазматической мужской стерильности. Ее индукция базируется на изменениях в широком диапазоне биохимических реакций, обусловливающих нарушения в молекулярных компонентах различных клеточных структур и цитоплазмы, что сопровождается появлением белков с анормальной эн-зиматической активностью. В результате этих изменений возникает дисбаланс в энзиматических системах или же полное их блокирование[8, 15, 20,82].
Для получения ЦМС путем химической индукции у сельскохозяйственных культур необходимо детальное изучение метаболизма ДНК, РНОК и белка в стерильном и фертильном материале с цитологическим и цитохимическим сопоставлением полученной информации. Изменения в содержании ДНК, РНК и белка в пыльниках кукурузы изучали методами цитохимии и микроденситометрии [101]. В процессе спорогенеза различали три пика интенсивного синтеза нуклеиновых кислот. Первый пик соответствует премейотическому периоду и ранней профазе, второй — приходится на интервал между поздним мейозом и стадией тетрад, третий (наименьший) — соответствует периоду, предшествующему митозу микроспор. В течение первого пика установлена самая высокая скорость роста спорогенной ткани и тапетума при активном синтезе молекулярных компонентов в обеих тканях. Во втором пике активный синтез ДНК, PlHiK и белка осуществляется в основном в тканях тапетума. Торможение роста и параллельно синтеза этих молекулярных структур в спорах начинается в период третьего пика, при дегенерировании тапетума. В настоящее время тапетум рассматривают как возможный источник предшественников нуклеиновых кислот для формирующихся микроспор, которые получают материал для синтеза в форме растворимых предшественников или в форме ДНК с низким молекулярным весом [111, 130].
Основная аккумуляция PlHiK в спорогенной ткани происходит в течение мейотической профазы, включая стадию па-хитены материнской клетки пыльцы. В тканях тапетума накопление РНК приходится на тот же период (профаза—зи-12
готена—пахитена) [98]. Таким образом, сразу после синтеза ДНК на ранних этапах спорогенеза до митоза в микроспо-пах продуцируется РНК спорогенной ткани, причем 75% пригодится на рибосомальную РНК — на формы 16S и 28S. Аккумулирование форм p:PHiK 4S и 5S осуществляется иначе, чем основных форм pPlHK, и пик их аккумуляций наблюдается в конце интерфазы микроспор [112].
В пыльцевом зерне основной синтез PHiK, ДНК и белка происходит на более поздней стадии — после митоза в микроспорах. .В цитоплазме пыльцевого зерна синтез всех форм РНК полиостью прекращается в последние 48 ч формирования пыльцы [94]. Биосинтез нуклеиновых кислот и процессы, связанные с их обменом, претерпевают определенные изменения под влиянием соединений с гаметощидными свойствами.
Установлено, что у фертильных соцветий кукурузы интенсивный синтез ДНК осуществляется на ранних этапах мик-роспорогенеза, но по мере завершения формирования пыльцы наступает торможение синтетических процессов [20]. При обработке растений гаметоцидами (0,15%-ным раствором три-атаноламиновой соли ГМК или 0,8%-ным раствором натриевой соли сф-дихлоризомасляной кислоты (FW-450), комбинированно 0,8%-ным раствором FW-450 и 0,1%-ным раствором гнббереллина) содержание ДНК на всех этапах формирования пыльцы несколько увеличилось независимо от применяемого гаметоцида. Это объясняется торможением клеточного растяжения под влиянием обработки гаметоцидами, в результате чего уменьшаются размеры клеток в спорогенной ткани и пересчет на взятую навеску дает завышенное содержание ДНК, не связанное с процессами аккумуляции нуклеиновых кислот [34].
Комбинированная обработка с гиббереллином (FW-450 + +гиббереллин) вызывает качественные изменения в молекулах ДНК, что приводит к нарушениям митозов и деградации ядра. Отмечено, что растворы ГМК и FW-450 по-разному влияют на интенсивность биосинтеза РНК в мужских репродуктивных органах кукурузы [26]. Параллельно было установлено, что формы проявления стерильности (морфологиче-ские_йзменения) находились в зависимости от применяемого вещества. Применение ГМК на кукурузе вызывало торможение образования РНК, и гаметоцид выступал здесь как ингибитор синтеза РН.К, обусловливая торможение клеточного удлинения. Морфологически это выражалось в уменьшении размеров соцветий, отсутствии в большинстве случаев колосков на латеральных веточках и в редукции листьев. Споро-генные ткани или совсем не формировали пыльцы, или она была нежизнеспособна. Действие FW-450 при данной концен-
13
трации'не вызывала значительных изменений в синтезе ДНК и PiHK, процессы спорогенеза у кукурузы протекали нормально [20, 26].
У фертильных растений высокое содержание PiHIK отмечено на более ранних этапах формирования пыльцы я дальнейший синтез РНК осуществлялся более активно [94, 98, 137].
Специфичность действия гаметоцида обусловлена различными факторам.и: близостью его химических характеристик к аналогам синтетических пулов клетки; возможностью энэиматического преобразования в клеточной системе в субстрат-подобный продукт ил;и в активный промежуточный метаболит определенных энздматических систем; наличием гормональных свойств или конформационного подобия с молекулами индукторов 'или репрессоров. Воздействие препаратов с гаметоцидными свойствами связано с физико-химическими характеристиками соединений (высокая электрофильность, способность к хелатированию и т. д.), в результате которых могут изменяться р1Н клетки и 'ионная сила в критические стадии развития спорогенной ткани [82].
Исследования влияния этрела на мейотические процессы в спорогенной ткани пыльников пшеницы Triticum aestivum L. показали, что индукция мужской стерильности связана с распадом этрела в растительных тканях с выделением этилена, молекулы которого обладают гормональными свойствами [47, 97]. Предполагают, что этилен, как и многие гормоны, влияет на функциональное состояние мембран, изменяя активность РНК полимеразы. Таким образом он может воздействовать на процессы транскрипции, особенно участков ДНК, ответственных за синтез долгоживущих мРНК, транскрибирующихся до мейоза, но необходимых для нормального течения мейотических процессов[58, 98].
'В результате возможных нарушений в синтезе различных форм PlHiK при включении гаметоцида в метаболизм клетки неизбежно возникают анормальности в синтезе белка. В нормально развивающейся спорогенной ткани и в микроспорах активный синтез белка приходится на раннюю мейотическую профазу, его активность несколько снижается в зиготене — пахитене и совсем незначительна в период формирования тетрад [101]. Тапетум как ткань проявляет очень высокую метаболическую активность в течение всего периода микроспо-рогенеза вплоть до полного автолиза, поэтому отводить тапе-туму только секреторную роль — значит ограничить его многообразные физиологические функции [108]. Отмечено, что пул ДНК тапетума недостаточен, чтобы служить источником для формирующихся микроспор [98]. Возможно, тапетум снабжает микроспоры растворимыми ДНК предшественниками,
14
как это было показано с экзогенно добавленным меченым ти-мидином, который быстро проникал в ткани тапетума и включался в ДНК микроспор [72]. Наиболее вероятно, что синтез РНК в тапетуме и микроспорах материнской клетки пыльцы независим. Однако это не исключает, что тапетум, особенно в ранний период формирования микроспор, частично поставляет предшественников РНК в спорогенную ткань [94]. В тапетуме имеется довольно большой рибосомальный пул, который, по-видимому, полностью деградирует вместе с ним или может поставлять предшественников РНК для последнего периода синтеза РНК в развивающихся спорах [101].
Роль тапетума в белковом синтезе может быть объяснена с точки зрения синтеза специфических энзимов, связаннь1х с мейозом или другими процессами в спорогенной ткани. Допустимо участие тапетума и в распределении белкового резерва. Высокая пропорция piPHK в его тканях свидетельствует о синтезе белков de novo, часть ,из которьгх откладывается как запасные в микроспорак [28, 137]. Обработка гаметоцидными препаратами вызывает репрессию синтетических процессов белка в результате вмешательства этих физиологически активных веществ во взаимосвязь процессов ДНК— РНК—белок [13, 19,29, 30, 104, 120]. Выяснение сущности процессов индукции мужской стерильности лежит в этой области. Эффект ГМК на ростовые процессы посредством действия ингибитора на обмен нуклеиновых кислот — экспериментально установленный факт [29]. Значительное влияние оказывает ГМК на рибосомальную фракцию РНК путем изменений в процессах биосинтеза ДНК [19]. Возможно, что специфичность действия ГМК проявляется на уровне репрессии синтеза биокаталитически активных белков именно той фракции, которая ответственна за синтез и распад ДНК. При воздействии ГМК наступает уменьшение фосфорелированных богатых лизином гистоновых фракций, количество которых в активноделящихся клетках при нормальных физиологических условиях значительно выше. Такое изменение в соотношении гистоновых фракций оказывает влияние на матричную активность ДНК, что приводит к нарушению мейотического цикла. Не исключено, что изменение соотношения форм гистонов под влиянием ГМК создает условия для атаки и расщепления ДНК'азой доступных участков ДНК, о чем свидетельствует возросшая активность этого энзима у растений, обработанных ГМК [19].
ГМК как гаметоцид не проявляет высокой селективности действия, так как параллельно оказывает влияние на мери-стематическую ткань, где стимулирует процессы распада и ингибирует синтетическую активность клеток [29]. Очевидно,
15
'этим объясняются негативные эффекты, наблюдаемые при опрыскивании растений растворами ГМК: задержка в росте и развитии, морфологические аномалии и т. д. [20, 125].
Изменения в деятельности центров, программирующих природу синтезируемых белков и регулирующих их синтез, при включении механизмов стерильности вызывают сложные сдвиги в ферментативных системах. Низкая активность различных энзиматшческих комплексов стерильных аналогов озимой пшеницы и угнетение активности окислительно-восстановительных процессов, лежащих в основе метаболизма, проявляются на ранних этапах формирования ,и развития микроспор [6, 128]. Дезорганизация в энзиматических системах вызывает дисбаланс в аминокислотном, углеводном и нуклеиновом пулах.
Хлорированные производные аляфатических карбоновых кислот, применяемые в качестве гаметощидов, вызывают накопление в тканях пыльников большого количества аминокислот (а-аланина, р-аланина, аспарагиновой, глутаминовой, се-рина и др.), одновременно резко снижается содержание про-лиаа [8, 26, 43].
При сравнительных исследованиях мужской стерильности у различных видов растений отмечены изменения в соотношении отдельных аминокислот в аминокислотном пуле. Количественный и качественный состав аминокислот обусловлены механизмами цитоплазматической регуляции и спецификой метаболизма растения. Так, в стерильных пыльниках сорго по сравнению с фертильными установлена анормальная акку^-муляция глицина и заниженное содержание аспарагиновой .кислоты, серина и аланина. М. W. Brooks, J. S. Brooks и -L. Chien [52] считают, что в данном случае специфическим признаком, обусловливающим мужскую стерильность, является блокирование глицина в синтезе пурина. У других линий сорго с ЦМС обнаружено повышенное содержание трео-нина по сравнению с фертильными растениями. Пыльники последних включали больше аланина, глутаминовой кислоты и тирозина [43]. Вероятно, от типа стерильности и этапа, на котором возникают нарушения в синтетических процессах .клетки, зависит накопление или ингибирование образования определенных аминокислот.
.Сравнительные исследования содержания аминокислот у различных растений с разными формами мужской стерильности (функциональной, цитоплазматической и ядерной) показали, что во всех случаях было высокое содержание аспа-рагина' и низкое — пролина по сравнению с фертильными растениями [26, 43, 46]. Последующие эксперименты подтвер-
16
дили, что пыльники стерильных аналогов линий, полученных на основе цитоплазмы Т. timopheevi, характеризовались
-высоким содержанием свободного аспарагина и низким — пролина. Среди связанных аминокислот сохраняется та же закономерность [118].
^ Индукция мужской стерильности при использовании химических соединений, обладающих гаметоцидной активностью
'-(RMiK, FW-450 4- гиббереллин), сопровождалась аналогичными изменениями в аминокислотном пуле растений кукурузы: накоплением больших количеств a-алани'на, аспарагиновой
•и глутаминовой кислот, серина у стерилизованных растений с одновременным резким блокированием синтеза пролина [26]. Часть аспарагиновой кислоты подвергалась дальнейшему аминированию, что приводило к образованию амида, содержание которого у стерилизованных растений в отдельных случаях достигало 2%, или приблизительно половины от общей суммы всех свободных аминокислот [8]. Отмеченный многими авторами дефицит пролина у растений с различным типом мужской стерильности предполагает торможение синтеза белка путем включения механизмов мужской стерильности на более ранних этапах белоксинтетической системы (при синтезе аминокислот), в результате чего тормозятся процессы аминирования.
Полноценное питание в течение микроспорогенеза и развития пыльцы обусловливает нормальное развитие генеративных клеток. Массовое накопление свободного пролина начиналось после завершения мейоза, а в фазе тетрад он был обнаружен в заметных количествах [8]. Эта аминокислота принимает участие в ряде общих реакций метаболизма: активирует дыхание растительных тканей, регулирует поглощение кислорода, является донором NHz-групп при синтезе некоторых аминокислот и стимулирует синтез хлорофилла [8]. Про-лин относят к числу характерных аминокислот щелочных ядерных белков-гистонов и протаминов, несущих генетическую функцию в поддержании определенной структуры ДНК [9]. Влияние этой аминокислоты на активность генома связано с критическими этапами в развитии, когда незначительные структурные перестройки могут иметь решающее значение. Исключительно высокое содержание пролина в тканях гаме-тофита обусловливает определенную физиологическую «нагрузку», которую он выполняет в процессах формирования пыльцы.
В пыльце диплоидных сортов содержание пролина в два раза выше, чем в пыльце триплоидных, имеющих пониженную трертильность [8]. Эти данные свидетельствуют, что пролин .занимает особое место среди других аминокислот при форми-2—10287 17
ровании пыльцы и, возможно, является определяющим метаболитом нормальных физиологических процессов.
Пролин пыльцы, по-видимому, вовлекается в самые ранние фундаментальные реакции гаметогенеза, и его дефицит в пыльниках может иметь прямое отношение к абортивности пыльцы. Содержание пролина начинает .снижаться на стадии мейоза и становится прогрессивным к периоду интерфазы микроспор, когда уже наблюдается дегенерация пыльцевого зерна. Дефицит пролина — это следствие некоторых дефектов в мейозе или в предшествующей стадии микроспорогене-за [118]. Параллельно накопление глутаминовой кислоты в стерильных пыльниках свидетельствует о торможении процессов превращения ее в пролин, предшественником которого она является. Относительно высокое содержание аспарагина в пыльниках растений с мужской стерильностью также обусловлено метаболическими отклонениями.
Индукция мужской стерильности вносит свои коррективы в биосинтетические процессы не только генеративных органов, но и всего растения. Общее содержание аминокислот в семенах и вегетативных органах стерильных растений пшеницы выше, чем фертильных, на 8%, что можно объяснить спецификой метаболической активности митохондрий стерильных растений [46, 63]. У нормально развивающихся растений накопление свободного пролина начинается после редукционного деления и осуществляется за счет притока пролина из вегетативных органов [8].
Наблюдения за изменениями в динамике развития организма, происходящими под влиянием веществ с гаметоцидной активностью во взаимосвязанной системе ДНК—iPHiK—белок—аминокислоты, показали, что наиболее быстро и в значительной степени изменяются активности энзиматических систем, затем проявляются ответные реакции в обмене нуклеиновых кислот и белка и относительно с большим опозданием осуществляются процессы, приводящие к изменению соотношения свободных аминокислот, органических кислот и аммония [98].
Многообразие действия хлорированных алифатических кислот на растительный организм предполагает возможность индуцирования мужской стерильности путем блокирования .разнообразных метаболических путей.
Попытки связать механизм избирательного действия были сделаны при изучении ряда хлорированных производных али-фатических карбоновых кислот, проявивших гаметоцидные свойства. Одним из предполагаемых механизмов селективного действия ар-дихлоризомасляной (FW-450) и трихлоруксус" ной (ТХУ) кислот считают торможение ферментативного син-
18
теза пантотеновой кислоты из пантоевой и (3-аланина [20]. Физиологическая роль пантотеновой кислоты связана с тем, что она является функциональной группой конденсирующего фермента КоА. Хлорированные алифатические кислоты выступают антиметаболитами р-аланина, поэтому синтез цантотено-вой кислоты является одним из чувствительных метаболических путей к этой группе соединений. Действие FW-450 определяется конкуренцией с 2,4-диокси-р-метилмасляной кислотой за локус, специфически активирующий фермент. Аналогичный эффект отмечен при действии 2, 3, 3-трихлормасляной к 2, 3-дихлормасляной кислот. Недостаток рибозы (одного из компонентов КоА) повышает токсичность FW-450 и этрела. Исследования гербицидного действия производных алифати-ческих карбоновых кислот показали, что эффект применяемых соединений обусловлен нарушением синтеза КоА. В результате возникает дисбаланс в соотношении ряда аминокислот трикарбонового цикла. При этом снижается количество лимонной и возрастает содержание яблочной кислоты, происходит более интенсивный синтез р-аланина и аспарагиновой кислоты.
В настоящее время преждевременно судить об определенной специфичности действия конкретных гаметоцидов, о «месте первичного действия» соединения. Возможно, механизм гаметоцидного действия определяется конкуренцией между веществом, обладающим гаметоцидньши свойствами, и определенными естественными метаболитами тканей генеративных органов.
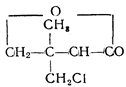
На основании исследований конкуренции между пантоте-натом и далапоном появилась возможность предсказать новые аналоги пантоата в виде хлорзамещенных алифатаче-ских кислот. Были синтезированы 4 соединения, биологическая активность которых (в данном случае гербицидная) варьировала в зависимости от степени хлорирования и место
|
|
положения хлора: (далапон); при кон
центрации 0,005 М активность далапона составляла 76%, при 0,05 М—99%.
| У соединения |
| активность при тех же |
концентрациях составляла соответственно 77 и 100%. 19
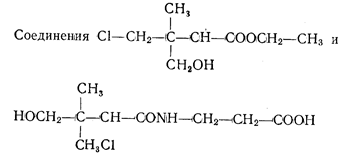
проявляли фитотоксический эффект: при концентрации 0,05 М он был равен 97 и 12%, при 0,05 М — соответственно 100 и 83%.
Таким образом, степень биологической активности препарата не имеет прямо пропорциональной зависимости от концентрации вещества, что свидетельствует о сугубо физиологической активности соединения, связанной с особенностями метаболизма растения.
В исследованиях по биохимизму действия ряда гербицидов установлено существенное влияние их на трансформацию энергия в клетке, повышение интенсивности окислительных процессов и угнетение фосфорелирования в митохондриях [10, 14, 21, 23]. Нарушение сопряженности окисления и фосфорелирования — результат угнетения активности многочисленных ферментов цикла Кребса и дыхательной цепи мито-хондрий. Получены дополнительные сведения о гербицидах, обладающих одновременно и гаметоцидной активностью. В частности, при нанесении далапона на растения люпина .изменялось соотношение сульфгидрильных и дисульфидных групп, входящих в состав активных центров многочисленных энзимов, участвующих в разнообразных ферментативных комплексах [13]. Кроме того, установлено повышенное содержание изофлавоновых глюкозидов и изменение их состава при o6pai6oTKe растений 2,4Д [22].
Появление хинонов — продуктов окислительного превращения фенольных соединений с высокой биологической активностью и их взаимодействие с амино- и сульфгидрильны-ми группами белков, сульфгидрильными группами аскорбиновой кислоты и другими SH-содержащими компонентами клетки обусловливают блокирование целых систем энзимати-чески взаимосвязанных комплексов. От окислительно-восстановительных условий и энергетических возможностей ткани, особенно спорогенной, зависят синтез и обмен важнейших органических соединений.
20
Характерными признаками ЦМС у сорго являются угнетение окислительно-восстановительных процессов и снижение энергетического обмена [41]. Различия в активностях АТФ'азы обнаруживались у стерильных аналогов уже в фазе тетрад и сохранялись в дальнейшем на всех фазах развития микроопор [16]. Среди соединений с гаметоцидными свойствами 2,4Д снижает содержание АТФ и АДФ — адениннук-леотидов, основных аккумуляторов энергии в клетке. Установлено, что 2,4Д ингибирует активность аденилаткиназы — фермента, осуществляющего равновесное соотношение компонентов аденилатного пула: 2 АДФ ^ АМФ+ДТФ [21].
Растительные гормоны, проявившие гаметоцидные свойства (2,4Д, ИУК, НУК, ГКз и т. п.), могут индуцировать мужскую стерильность на тех уровнях метаболических процессов, на которых они оказывают свое регуляторное действие:
на уровне генома, мембран, аллостерического эффекта. Возможно и одновременное влияние их на разные уровни, но во всех случаях отмечена взаимосвязь физиологически активных веществ, к которым относятся гаметоциды, с изменениями в энергетическом обмене клетки.
Существование специфических рецепторов в клеточных структурах и мембранах, способных обратимо связывать ауксины [25], может служить молекулярно-биологической интерпретацией действия ряда соединений, проявивших гаметоцид-ную активность на различных культурах и относящихся к ауксинам (ИУК, НУК, 2,4Д, Г.Кз, кинетин и др.) [11, 12, 14, 17, 58, 135].
Отмечено, что растительные гормоны (2,4Д, ИУК, ГКз), вызывающие при определенных концентрациях различную степень индукции мужской стерильности, влияют на активность энзимов, связанных с метаболизмом углеводов, определяющих структуру клеточных оболочек, с такими как р-1,4-глюканаза, р-1,3-глюканаза, (3-1,6-глюканаза я гемицеллю-лаза, а также а-1,3- и а-1,6-глюканазы [73, 99, 136]. Повышение активности глюканазных энзимов взаимосвязано с процессами деструкции их субстратов, а следовательно, и с изме-нениями в каллозной оболочке материнской клетки пыльцы и формирующихся тетрад, так как она является Р-1,3-свя-занным полимером глюкозы. Установлено, что ИУК и 2,4Д способствуют увеличению р-1,3-глюканаз'ной активности, в результате чего разрываются перекрестные связи в пределах клеточных стенок и оболочек, что обусловливает возрастание их эластичности и проницаемости [55].
Введение ИУК в растительную клетку повышает утилизацию глюкозы путем активации энзима УДФ-зависимой глю-кансинтетазы, локализованной в пределах аппарата Гольд-21
жи, что способствует формированию и повышенному содержанию глюканов, галактанов и пентозанов [42]. Подобным образом 2,4Д включается в один из уровней метаболизма клетки (через углеводы, путем активации плазменной, связанной с мембранами глюкансинтетазы), что способствует утилизации УДФ-арабинозы я увеличению количества связанных остатков арабинозы с галактаном [136]. Повышение числа сшивок в молекулах галактана изменяет пластичность клеточных стенок. Вместе с тем аккумуляция 2,4Д в мембранах вызывает нарушение комплекса связанного с мембранами белкового фактора, который обусловливает активность PHiK полимеразы, транскрибирующей определенные мРНК [66].
В опытах по конкурентному вытеснению связанных эффек-торов (производные феноксиуксусной кислоты и ИУК) наглядно продемонстрировано, что связывание биологически активных хлорированных производных мембранами (эффектор-рецептор) носит специфический характер [14]. Изменяя функциональную активность мембран и связанных с ними энзи-мов, ауксины с гаметоцидными свойствами могут вызывать индукцию синтеза определенных мРНК, ответственных за •продуцирование ряда энзимов, среди которых имеются ферменты, преобразующие углеводные компоненты мембран и клеточных оболочек. Возрастающая при этом проницаемость может вызывать нарушение селективной изоляция формирующихся тетрад с последующим их деградированием. Предполагают, что каллозная оболочка функционирует как «молекулярный фильтр», позволяющий проникать внутрь материнских клеток пыльцы основным питательным элементам, за исключением больших молекул. Последние в эту раннюю фазу могут помешать установлению автономии ядра гаплоид-ной споры в пределах собственной цитоплазмы [98].
Химическая изоляция материнских клеток пыльцы в стадии тетрад от окружающей диплоидной цитоплазмы является необходимой предпосылкой нормального развития пыльцы [94]. Установлено, что меченый тимидин поступает в материнские клетки пыльцы только до формирования каллозной оболочки, но не проникает, если они заключены в каллозу [72]. При изменении последней и освобождении тетрад метка свободно поступала в микроспоры. Эти наблюдения позволили сделать вывод о функционировании каллозной оболочки как «молекулярного фильтра».
Каллозное покрытие материнских клеток пыльцы функционировало как молекулярное сито: каллоза пропускала глюкозу и углекислый натрий, но задерживала фенилаланин, размер молекулы которого гораздо меньше глюкозы и он 22
должен был бы легко проникать в материнские клетки пыльцы [123]. Выводы J. Heslop-iHarrison и A. Mckenzie [72] также сомнительны, так как метка могла не включаться ввиду отсутствия синтеза ДНК. В других исследованиях показано, что роль каллозной оболочки значительно сложнее, чем «простого молекулярного фильтра» [4, 130].
ПОСТУПЛЕНИЕ В РАСТЕНИЯ И РАСПРЕДЕЛЕНИЕ В НИХ ФИЗИОЛОГИЧЕСКИ АКТИВНЫХ ВЕЩЕСТВ
Для успешного поиска химических соединений, обладающих высокой селективностью гаметоцидного действия, необходима информация о поступлении и распределении экзрген-но наносимого на стебли и листья препарата. Степень поступления и скорость распределения химических соединений зависят от морфо-физиологических особенностей листовой пластинки, химических характеристик наносимого препарата, окружающих условий и других факторов. У различных видов культурных растений, с которыми проводили эксперименты с целью химической индукции мужской стерильности, в значительной степени варьирует толщина кутикулы и соотношение ее компонентов (воск, кутан, пектин, целлюлоза), а также внешняя эпидермальная оболочка [44, 56, 60, 65, 69, 75].
С помощью химических методов анализа исследовали ку-тикулярный слой листьев 24 видов овощных и плодовых культур [77]. Установлено разнообразие в составе и соотношении веществ листовой поверхности и значительные различия в толщине и составе мембраны кутикулы. Однако основные структурные компоненты кутикулы характерны для всех растений. Кут.ин представляет собой полимолекулярную сеть гид-рокси-карбокоильных кислот с простыми и сложными эфирными связями. С кутанным матриксом связаны полисахар.иды и кутикулярный и этикулярный воск, богатый алканами. Так как в кутикуле имеются полярные и неполярные группы, она обладает как гидрофильными, так и липофильными свойствами [45].
Одним из важных факторов, определяющих степень эффективности действия гаметоцида, является зависимость его. активности от климатических и погодных условий, которые необходимо учитывать при разработке доз и концентраций для конкретных возделываемых культур и представляющих их сортов. Однако до сих пор не удалось установить четких взаимосвязей между влиянием условий среды и проницаемостью листовой поверхности, хотя развитие кутикулы коррели-
23
рует с продолжительностью светового периода, инсоляцией, влажностью почвы и воздуха, температурой [81].
Температурный режим является одним ,из самых определяющих условий развития кутикулы и распределения наносимого препарата [83]. При нанесении препарата в разные фазы органогенеза следует учитывать, что с возрастом листа снижается абсорбционная способность. Р. М. Nelson и R. К. Reid fl05] доказали, что метиловые эфиры жирных кислот — Lg-ii вызывают гибель меристематических тканей, но не разрушают более зрелые ткани. Механизм такой селективности основан на способности кутикулы действовать как барьер проницаемости по отношению к наносимому препарату. Это было подтверждено разрушением кутикулы до нанесения эмульсии (4%-ный метил-деканоат+0,1%-ный твин 20). Гибель ткани (ожоги) — результат нарушения структуры мембран. Авторы считают, что твин снижает проницаемость кутикулы и плохо проникает через кутикулу зрелых листьев.
Поверхностное применение гаметоцидных препаратов с использованием радиоактивной метки показало, что при нанесении на лист гаметоциды быстро транслокализуются в репродуктивные органы. Нанесение химикатов на базальную часть листа обеспечивало более полное поглощение препарата, чем на апикальную часть. В репродуктивных тканях активно аккумулировались гаметоциды.
Наиболее подробно абсорбционные особенности химических препаратов изучали на гербицидах. Детально были исследованы ответные реакции растений двух сортов капусты, характеризующиеся различной чувствительностью к нитрофе-ну (2,4-дихлорфенил-р-нитрофениловый эфир) [113]. У обоих сортов не было обнаружено заметных различий в скорости прорастания семян, росте растений, плотности устьиц листьев и в транслокализации меченого '^С гербицида. Однако у растений устойчивого к препарату сорта листья обладали более восковидной поверхностью, что позволило сделать вывод о корреляции между блокированием абсорбции и содержанием воска в кутикуле.
Проникновение соединений в ткани листа обусловлено и другими факторами, характеризующими применяемый раствор: температурой, концентрацией, продолжительностью времени его соприкосновения с поверхностью растений, поверхностным натяжением и др. Проникновение веществ возрастает пропорционально времени и концентрации. Поступление соединения из раствора в ткани листа осуществляется до тех пор, пока его доза на поверхности листа или под кутикулой не станет предельной. В полевых условиях эффективность действия и поступление нанесенного препарата зависят от
24
сложного взаимодействия климатических факторов и внутренних физиологических тканевых параметров (водный дефицит, значение рН в клетке, концентрация цнтоплазматиче-ского сока и т. д.).
В ткани листа легче проникают недиссоциирующие молекулы. Повышение температуры от 10 до 30° увеличивает проницаемость кутикулы и мембран, за исключением промежутка между 15" и 25°, в пределах которого поступление носит относительно стабильный характер. Если опрыскивание проводят под давлением, растворы проникают в основном через устьица, однако вещества с поверхностным натяжением, близким к поверхностному натяжению воды (70 дин/см2), через устьица не проникают ,[54].
При селективности абсорбции у наиболее распространенных видов сельскохозяйственных культур по отношению к 2,4Д (который обладает и гаметоцидными свойствами) установлено, что толщина кутикулы является решающим фактором, а толщина и структура оболочек эпидермальных клеток не имеют определяющего значения для степени абсорбции препарата. При этом абсорбционные характеристики старых и молодых листьев значительно отличались и в обоих случаях зависели от химических свойств препарата.
На проростках конских бобов поглощение листьями меченого по углероду '^'^^-хлор-о-толил) окиси] бутиловой кислоты было одинаковым для листьев всех возрастов. Однако замена бутиловой кислоты на радикал уксусной кислоты вызывала различия в поглощении молодыми (скорее) и более зрелыми (медленнее) листьями [92]. Таким образом, даже слегка измененная конфигурация молекулы может в значительной степени повлиять на ее способность проникать через мембраны [126].
Пока не установлено четкой взаимосвязи между химическими, физическими характеристиками и проницаемостью препарата в системе клеток.
С помощью энзиматически изолированной кутикулы листьев была создана шкала проницаемости кутикулы по отношению к ряду алифатических спиртов и их амидов [138]. Соединения выбирали по способности растворять липиды и по моле< кулярньш весам. Результаты опытов показали, что кутикула функционирует в основном как липоидная мембрана, позволяющая веществам проникать в клетку пассивной диффузией согласно их растворимости в липидах. Модель молекулярного сита больше подходит для молекул малых размеров. Коэффициенты проницаемости алифатических спиртов располагаются от этанола к пентанолу: этанол<метанол<пропа-
25
нол<бутанол<пентанол, а в случае амидов: ацета1мид<про-пионамид<.бутиламид<1валерам:ид [121].
Однако при подборе алкильных групп для улучшения абсорбция препарата следует также учитывать возрастание фитотоксичности соединения. При исследовании действия четырех о-алкилметилфосфонофторидов на проростках пшеницы была отмечена зависимость фитотоксичности от природы о-алкильной группы, которая усиливалась в следующем порядке: этил<изопроп'ил<пипаколил<циклогексил. При этом продукты разложения были менее токсичны [79].
Методами газовой хроматографии и с помощью сцинтил-ляционного счетчика изучали различия в абсорбции буто-ксиэтилового эфира и соли аммония 2,4,5-Т (2,4,5-трихлорфе-ноксиуксусная кислота). Сравнимые результаты показали, что абсорбция 2,4,5-Т была в 2—3 раза выше при использовании препарата в форме эфира, чем в форме солей. Однако их передвижение в проводящих системах растения не зависело от химической формы соединения [100].
В опытах с энзнмат.ически изолированной кутикулой плодов томата, которая по своим характеристикам близка к кутикуле листа, степень хлорирования (но не положение хлора при замещении) феноксиуксусной кислоты увеличивала проницаемость ее галогенопроизводных в следующем порядке:
2,4,5- и 2,4,6-трихлорфеноксиуксусная кислота >2,3- и 3,5-ди-хлорфеноксиуксусная >2-хлорфенокоиуксусная>фенокс.иук-сусная кислота. Чем больше полярность феноксиуксусной кислоты, тем легче она проникает через мембраны [53]. Хлорирование увеличивает растворение феноксиуксусной кислоты в л.ипидах и тем самым способствует ее проникновению через кутикулу [127].
Хлорирование бензойной кислоты, наоборот, снижает степень проникновения через кутикулу, и шкала ее проницаемости располагается в нисходящем порядке: 2-хлорбензойная кислота, 2,4- и 2,5-дихлорбензойная и 2,3,6-три.хлорбензойная кислота. J. L. Stoddart [132] объясняет это низкой растворимостью галогенопроизводных бензойной кислоты при заданных рН (2,5; 3,5 и 5,2) в липидах. Степень хлорирования, фто-рирования и метилирования препаратов обусловливает их биологическую активность и токсичность действия на растительный организм.
Избирательная способность по отношению к абсорбции различных веществ растительными тканями связана с физическими свойствами соединений, включая константу их диссоциации при различной степени хлорирования, фторирова-ния, метилирования и т. д., а также скорость их распределения в липидной фазе при данной рН [23, 24, 91]. При обработ-26
ке растений препаратами их проницаемость могут улучшать некоторые вспомогательные вещества, особенно поверхностно-активные (ПАВ), которые улучшают контакт между препаратом и поверхностью листа, а также повышают степень проникновения препарата в растительный организм. Так, абсорбция 2,4Д возрастала в 7—8 раз при введении в раствор тви-на 80 (концентрация 1%) [57].
С помощью '^I" и "'I" метки изучали «крепление» на поверхности листьев сои препарата ТИБА (2,3,5-тр.ийодбензой-ная кислота) в концентрации 200 мг [110]. (ТИБА относится к веществам, обладающим гаметоцидной активностью). Были взяты четыре формы солей ТИБА (натриевая, диметил-амин, диэтиламин и триэтила'мин) в сочетании с четырьмя ПАВ: твин 20 (полдокоиэтилен сорбитан монолаурат), тритон 100 (октил фенокоиполиэтоксиэтанол) — оба неионные ПАВ; арквад 50 (алкил четвертичный аммоний хлорид) —ка-тионное ПАВ и игепон Т-77 (натрий 1\Г-метил-1\Г-омоил та-урат) — анионное ПАВ. Все ПАВ исследовали в двух концентрациях — 500 и 2000 мг/кг.
Взаимосвязь физико-химических свойств ПА1В и их концентраций с формами солей ТИБА носила сложный характер. ПАВ оказывали значительное влияние на закрепление препарата на растениях, а наибольшая разница в степени «крепления» солей ТИБА встречалась в пределах неионных ПАВ. Следовательно, это лимитирует выбор ПАВ среди неионных классов, свидетельствуя о их неспецифичности по сравнению с ионорганическими классами ПАВ. Физиологическая активность препарата зависела от подбора ПАВ. Низкие концентрации ионных ПАВ (500 мг/кг) достаточны для достижения максимума прикрепления нанесенного препарата. Не было отмечено взаимосвязи между формами солей и ПАВ. Во всех случаях для эффективной абсорбции требовалась высокая концентрация неионного ПАВ (2000 мг/кг), высокие же концентрации ионных ПАВ не улучшали степени закрепления препарата, что свидетельствует о физико-химической взаимосвязи, включающейся в комплекс проницаемости [11 б].
Активность препарата в основном проявляется при возрастании концентрации этиленокоида в ПАВ, когда его молекулы становятся более гидрофильными или соотношение гидрофильных (ГФ) группировок к липофильным (ЛФ) в молекуле ПАВ довольно высокое. Таким образом, для каждого химического соединения с гаметоцидной активностью подбор ПАВ и его концентраций зависит от физиолого-химических свойств активного ингредиента.
27
Степень абсорбции препарата растительными тканями во многом зависит от соотношения группировок ГФ/ЛФ в молекуле ПАВ для определенных концентраций гаметоцида. В опытах с энзиматически изолированной кутикулой листьев груши получены результаты, свидетельствующие об изменении проницаемости 2,4Д в зависимости от значения ГФ/ЛФ (106]. При величине соотношения у ПАВ ГФ/ЛФ, равной 16,7, не отмечено изменений проницаемости в шкале концентрация 2,4Д от 0,05 до 1%. Другое неионное ПАВ с ГФ/ЛФ=8,6 способствовало увеличению абсорбции 2,4Д в 10 раз при значениях его концентраций, близких к 1%. Третье неионное и высоколипофильное ПАВ (ГФ/ЛФ =4,3) увеличивало поглощение 2,4Д в 15 раз при всех заданных концентрациях от 0,1 до 1%. Эти исследования показали важную роль активного ингредиента, сопутствующего ПАВ, в регуляции абсорбции.
Опыты по поглощению и распределению метазола [2-(3,3-дихлордион)] в смеси с ПАВ (полисборбатом) при различных соотношениях ГФ/ЛФ подтвердили, что эффект ПАВ обратно пропорционален значению ГФ/ЛФ. Среди различных параметров для достижения высокого эффекта абсорбции гаметоцида тканями соотношение ГФ/ЛФ в молекулах ПАВ— наиболее влиятельный фактор, определяющий степень абсорбции. Трудность подбора такого ПАВ для каждого определенного гаметоцида связана с тем, что соотношение ГФ/ЛФ может в значительной степени координироваться сложной взаимосвязью химических и физических свойств активности ингредиентов смеси, морфологическими и цитологическими особенностями листа. Поэтому для каждой культуры необходим дифференцированный подход при подборе ПАВ для получения оптимального эффекта химической индукции мужской стерильности, вызываемой гаметоцидом.
Некоторые неионные ПАВ силикон-гликолевой природы по сравнению со стандартными неионными органическими ПАВ могут в большей степени повышать эффективность химически активных веществ благодаря улучшению абсорбции тканями. Однако эта группа ПАВ, обладающая большей эффективностью, чем катионные аминосиликоны, имеет отрицательное свойство — низкую растворимость в воде [86]. Несмотря на это при всем разнообразии применяемых в сельском хозяйстве химически активных веществ, в том числе и гаметоцидов, имеется возможность объединить препараты на основании одинакового характера абсорбции, что облегчило бы поиск и рекомендации ПАВ для этих групп.
Характер абсорбции некоторых фторсоединений подобен абсорбции 2,4,5-Т. Препараты наносили на листья капельным методом в смеси с ПАВ, характеризующимися различным со-
28
отношением ГФ/ЛФ: полиоксиэтилен (ГФ/ЛФ=20), сорби-тан монолаурат (ГФ/ЛФ=16,7), сорбитан моностеарат (ГФ/ЛФ =9,6) и полиоксиэтилен (ГФ/ЛФ=4). В июне наиболее эффективным было применение ПАВ в соотношении ГФ/ЛФ =9,6, в июле—августе — с соотношением 16,7. Абсорбция веществ, нанесенных на лист, осуществляется через трихомы и устьица, откуда соединения распределяются латераль-но через эпидермальные клетки. При исследовании по подбору .ПАВ для эффективной абсорбции веществ с гаметоцидной активностью для каждого вида необходимо учитывать (кроме .физико-химических характеристик самого препарата) стадию .развития растения и возрастные изменения морфологических характеристик листа, обусловливающие смачивающую способность его поверхности и относительное значение специфической абсорбции [100].
ПРИМЕНЕНИЕ ГАМЕТОЦИДОВ(КОНЦЕНТРАЦИИ,ДОЗЫ И СРОКИ ОБРАБОТКИ)
Химические препараты как источники гаметоцидной активности были выделены из биологически активных веществ различного физиологического действия (ростактивирующие вещества, ретарданты, гербициды, растительные гормоны .и гор-моноподобные вещества и т. п.). Среди хорошо известных физиологически активных соединений гаметоцидная активность была обнаружена у веществ, обладающих ретардант-к'ыми свойствами: этрела (этефон) — 2-хлорэтилфосфоновая кислота, далапона — 2,2-дихлорпро.пионовая кислота, ГМК.
Некоторые соединения, кроме фирменных названий, получили определенные шифры как вещества, проявившие гамето-цидные свойства: мендок, или FW-450 (2,3-дихлоризомасля-ная кислота), FW-676 (кальциевая соль 2,3-дихлоризомасля-ной кислоты), G-315 (магниевая соль 2,3-дихлоризомасляной кислоты), u'niroyal D-513 (пропаргил 2-октосульфит), OCDP [N (р-алорфенил) - 2,4-диметил-6-оксо-3,6-дигидроникоти'новая кислота], RH-531 [натриевая соль 1-(р-хлорфенил)-1,2-дигид-ро-4,6-ди'метил-2-оксоникотино'вая кислота].
Первые опыты с применением этрела в качестве гаметоцида для мягкой пшеницы были выполнены в 1961 г. К. В. Porter и A. F. Weise [116]. Предварительные эксперименты в вегетационных сосудах с сортами мягкой яровой пшеницы Marled и Thatcher дали обнадеживающие результаты. При обработке растений в фазы кущения, выхода в трубку и колошения растворами в концентрации 100, 250, 500, 750, 1000, 2000 и 2500 мг/кг из расчета 30 мл на сосуд с тремя растениями было установлено, что этрел, начиная с дозы 750 мг/кг, индуцировал 100%-ную стерильность у сорта Marfed. В этих
29
опытах исследователи впервые столкнулись с проблемой сортовой специфичности ответной реакции на обработку гамето-цидом.
'Степень стерильности при тех же дозах этрела у сорта Thatcher была значительно ниже, чем у Marfed, так как Thatcher был менее чувствителен к 0'бработке препаратом. В полевых условиях опыты проводили с сортом мягкой озимой пшеницы Nugaines. Обработку этрелом осуществляли в те же фазы концентрациями 500, 1000, 1500, 2000 и 3000 мг/кг. Опытные и контрольные варианты размещали рядом с сортами-опылителями, цветущими в разные сроки. Полная мужская стерильность была отмечена у растений пшеницы, обработанных растворами этрела в концентрации 1500, 2000, 3000 мг/кг. Максимальная стерилизация достигалась при нанесении на растения растворов этрела повышенной концентрации — 2000 и 3000 мг/кг в конце фазы выхода в трубку. При обработке в период колошения эффективность препарата снижалась. В опытах сохранилась высокая степень женской фертильности. Стерильные растения по морфологическим признакам были сходны с растениями пшеницы, обладающими UjMC. Применяемые концентрации вызывали укорачивание междоузлии и анормальное колошение отдельных растений [116].
Дальнейшие исследования были направлены на поиск эффективного сочетания оптимальных доз, концентраций и сроков обработки этрелом зерновых. Так, в экспериментах по 'ян-дуцированию мужской стерильности у растений мягкой пшеницы, проведенных в условиях вегетационного домика и в поле, раствор этрела в дозах 500, 1000, 2000 и 3000 мг/кг наносили в начальной фазе выхода в трубку и в конце ее. Наиболее эффективными в данных условиях были дозы этрела 1000 и 2000 мг/кг. Несмотря на нежелательные явления (усиленное кущение, замедленное колошение, торможение роста и развития, морфологические анормальности, приводившие в ряде случаев к гибели растений, и, как правило, к снижению урожайности), Р. L. Rowelil и D. G. Miller [122] считают, что надежды на этрел как на вещество, селективно вызывающее мужскую стерильность, довольно обоснованны и имеют практические возможности, которые лимитируются лишь разработкой конкретных методов обработки.
J. Law и N. С. Stoskopf [95] применяли этрел в полевых условиях в различные фазы морфогенеза ярового ячменя. Авторы пришли к выводу, что в условиях Канады лучшим периодом для обработки растений гаметоцидом является середина фазы выхода в трубку (фенологически — период появления предпоследнего листа) при одноразовом внесении до-
3J
зы 1,68 кг/га, которая давала приемлемый уровень мужской стерильности и не вызывала редукции женской фертильности. Этрел у ярового ячменя не проявлял гаметоцидного воздействия на яйцеклетку и являлся более потенциальным стерилизатором, чем другие применяемые с этой целью вещества, при условии его правильной комбинации с антиретардантны-ми компонентами.
При изучении оптимальных доз этрела (между 1000 и 2000 мг/кг) и времени его применения на пшенице (сорт Sirius) была поставлена цель — не только получить полную мужскую стерильность, но и устранить его побочное действие [80]. На основе предыдущих исследований было рекомендовано применение этрела до начала мейоза в материнских клетках пыльцы, когда достигалась наибольшая степень индукции мужской стерильности у пшеницы [47]. Нанесение этрела после мейоза вызывало нежелательные явления, приводящие к уменьшению количества колосков в колосе, которое сопровождалось определенной степенью стерильности макрота'мет (в зависимости от дозы препарата). Концентрация этрела 2000 мг/кг, вызывающая стерильность пыльцы, может также приводить к частичной или полной задержке колосьев во влагалищах листьев в результате сокращения соломины верхнего междоузлия. Наряду с этим, этрел стимулирует развитие, побочных стеблей, пыльцевые мешки которых часто находятся за пределами фазы применения этрела и не совпадают с премейотической фазой археспориальной ткани главного колоса.
Подобные затруднения практического характера возникают в любом случае при использовании различных гаметоци-дов. '"Определенную степень стерильности можно химически индуцировать почти на всех этапах развития растений, за исключением тех, когда уже сформировались зерна. Применение гаметоцкдов на ранних этапах развития растений не только индуцирует мужскую, а чаще общую стерильность, но и вызывает патологические формативные эффекты в растительных тканях. В зависимости от того, на каком этапе произошло «включение сигнала» механизмов мужской стерильности, можно получить различные типы мужской стерильности, которые классифицировали на 4 группы [64].
К первой группе относится стерильность, полученная за счет блокирования питания в результате прекращения развития и нормального течения процессов мейоза во внешне нормальных материнских клетках пыльцы, что приводит к образованию в открытых цветках пыльников, содержащих монады или диады. Ко второй группе относится стерильность, при которой отмечена дезорганизация процессов в материнских
31
клетках пыльцы в течение I и II мейотических профаз, что вызывает иррегуляцию при расхождения клеток в процессе двух мейотических делений. В материнских клетках в данном случае на стадии тетрад образуется изолированная группа клеток (более четырех), и микроспоры прекращают рост после ранней стадии развития экзины. Третья группа стерильности характеризуется анормально маленькой пыльцой с очень тонкой экзиной. К четвертой группе относится тип мужской стерильности, обусловливающий формирование нормальной пыльцы, но без терминальной поры.
При испытании веществ на гаметоцидную активность и поиске оптимального сочетания доз, концентраций и сроков их нанесения необходим цитологический контроль за микро-и макроспорогенезом.
Для каждой культуры существует критический период, при котором применение препарата обеспечивает наибольший эффект мужской гаметоцидной реакции. Согласно полученным данным, стадия мейоза микроспорогенеза наиболее критическая для применения этрела на зерновых, обеспечивающая максимум индукции мужской стерильности. Этот период у пшеницы приходится на конъюгацию хромосом в течение мейоза и формирование поры пыльцевого зерна [47, 48]. Однако очень важным периодом для индукций мужской стерильности является, по-видимому, и премейотическая фаза [94, 98].
В опытах с сортами озимой пшеницы Безостая I .и Nugai-nes цитологически контролировали изменения в микроспоро-генезе в зависимости от сроков нанесения этрела. Отмечено, что наибольшая эффективность .индукции мужской стерильности достигалась при обработке растений в фазе двух- и трехъядерной пыльцы [30]. 'Под влиянием этрела уже на стадии двухъядерной пыльцы наблюдались аномалии в развитии вегетативного ядра, контролирующего трофические функции при формировании пыльцевого зерна по отношению к спермиям [28]. Этим можно объяснить, что в пыльцевых зернах с возникающими под действием этрела митотически-ми нарушениями не накапливается крахмал [48]. Считают, что действие этрела связано с включением его в эпигенетические механизмы, действующие в период мейоза [30]. Это обусловливает нарушение в координации трофических функций вегетативного ядра, переключая его на митотическое деление, т. е. на клеточном уровне этрел действует не как ингибитор роста, а наоборот, стимулирует вторичное деление ядер, которые в норме никогда не делятся.
Точное определение критического периода воздействия га-метоцида имеет свои сложности, так как в литературе рекомендована широкая шкала доз при использовании одного и
32
| 1 |
того же препарата на определенной культуре. Для этрела рекомендуемые концентрации для обработки зерновых культур, включая тритикале, колеблются от 1500 до 10000 мг/кг [50, 116, 119, 125].
В условиях Италии почти полная мужская стерильность у яровой пшеницы была индуцирована при одноразовой обработке растворами этрела с концентрациями 4000 н 8000 кг/га в раннюю фазу выхода в трубку. При этом отмечено снижение урожайности до 40% [50]. В опытах с зерновыми, характеризующимися кущением, при котором появление боковых пооегов растянуто во времени как в пределах отдельного растения, так и у различных представителей в популяции, возникают трудности, связанные со спецификой данной культуры.
В опытах, проведенных в Великобритании в 1973 г., в посевах яровой пшеницы сорта Sirius на 1 апреля пыльники немногих растений находились в премейотической фазе (наиболее эффективный период для нанесения препарата), 5 апреля — 5/%, а 10 апреля — 100% пыльников всех растений находились в постмеиотической фазе и только третьи и четвертые цветки и молодые колоски имели более раннюю фазу развития [80].
С возрастом пыльники становятся менее чувствительными к этрелу, и для достижения нужного эффекта потерю чувствительности компенсируют повышением дозы гаметоцида. Однако при этом возникает опасность редукции женской фер-тильности. Сжатые сроки обработки (от 5 до 12 дней) между двумя периодами развития должны совпадать с премейотической фазой в археспориальной ткани и мейозом. Вторичную обработку следует проводить в конце мейоза. [47]. Таким образом, наиболее эффективный период воздействия этрела может включать мейоз и даже раннюю фазу развития пыльцы, но высокие концентрации препарата, необходимые для хорошей индукции мужской стерильности, влияют на рост и развитие самого растения. Разрыв во времени между двумя обработками может быть более длительным (до двух недель) в зависимости от климатических условий (осадки, пониженная температура).
Трудность подбора определенного гаметоцида для любой культуры включает и климатические факторы. В условиях районов Онтарио (Канада) установлено, что этрел действует больше как ретардант, чем гаметоцид, при тех же дозах, которые в других климатических зонах дают высокий уровень мужской стерильности [95]. При использовании этрела на яровой пшенице Sirius в условиях теплиц в концентрации 2000 мг/л перед окончанием мейотической интерфазы разви-
3—10287 33
тия микроспор достигалась полная'' мужская стерильность только в нижних цветках ранее образовавшихся колосков, и, хотя женская фертильность оставалась высокой, перекрестное опыление ограничивалось неполным выходом колоса .из влагалища верхнего листа вследствие сильного укорачивания стержня колоса [80]. Этого удалось избежать путем последующего опрыскивания растений (через 2—5 дней) растворами ГКз в концентрациях 100—300 мг/л. Данные дозы не оказывали влияния на индуцированную этрелом муждкую стерильность в условиях теплицы, но в полевых опытах ГКз в сочетании с этрелом действовала как синергист в процессах стерилизации пыльцы; в результате оказалось возможным уменьшить дозы этрела.
На участке гибридизации отмечено нормальное выколаши-вание 17 сортов пшеницы при дозе этрела 6,4—12,8 кг/га и ГКз — 1,1 кг/га. Выход гибридных семян составлял 60— 75%. Однако этот показатель можно улучшить, если обрабатывать этрелом одновременно не все сорта, а каждый в отдельности в соответствии с наступлением оптимального срока опрыскивания [80].
Обнадеживающие результаты, полученные с этрелом на яровой пшенице, не получили подтверждения в опытах с озимой пшеницей, в которых этрел (2000 и 4000 мг/кг) вызывал лишь частичную стерильность, редуцировал женскую фертильность, тормозил рост и развитие как всего растения, так и самого колоса, негативно влияя на формирование семян
[67].
Этрел, натриевую соль ГМК, а также новые препараты испытывали в качестве гаметоцидов в Нечерноземной зоне РСФСР на посевах сортов мягкой (Московская 21, Минская и др.) и твердой (Гордеиформе 432) пшениц. Установлена различная реакция сортов на действие препаратов. При обработке этрелом отмечено меньшее количество цветков с недоразвитыми пыльниками у сорта Минская и образца СВ 6009 по сравнению с сортами' Гордеиформе 432 и Московская 21. Так, в 1973 г. двукратная обработка растений раствором этрела в концентрации 0,6% снижала количество зерен в колосе у сорта Минская с 22,9 до 9,5, а у Гордеиформе 432 — с 19,2 до 0,2 [36].
Количество цветков с индуцированной мужской стерильностью увеличивалось по мере повышения концентрации раствора. Если принять количество зерен главного колоса контрольных растений сорта Московская 21 за 100%, то у обработанных этрелом (концентрация раствора 0,3%) растений их число составляло 64,4%. Повышение концентрации гамето-цида приводило к более резкому снижению количества зе-
34
рею при концентрации 0,4% получали 46% зерен по сравнению с контрольными растениями, при 0,45% — соответственно 32%, при 0,50% — 23%, при 0,55% — 15%, а при 0,60% — только 13,3% [36].
В опытах также было установлено ингибирование роста растений, торможение удлинения последнего междоузлия, в результате чего, особенно в засушливые годы, часть колосьев частично или полностью не выколашивалась. Побочное отрицательное действие этрела несколько снижалось при использовании раствора ГКз, наносимого отдельно и совместно с га-метоцидом [37].
Судя по редукции женской фертильности, которая сопровождает процессы абортивности пыльцы при обработке зерновых этрелом, это соединение не обладает высокой селективностью гаметоцидного действия. Многие авторы [50, 75, 85, 95, 116] при рекомендации этрела как препарата с гаметоцид-ной активностью не учитывают вносимой дозы, а указывают только концентрацию и сроки обработки. Целесообразно дифференцировать эти дозы в зависимости от климатических факторов, сортовых особенностей культуры и ее чувствительности. к препарату.
В трехлетних опытах, проведенных в условиях Нечерноземной зоны с сортами озимой мягкой пшеницы Мироновская 808, Мироновская юбилейная и Немчнновская 121, была установлена возможность успешной стерилизации пыльцы 0,5% -ным водным раствором этрела при внесении его оптимальной дозы — 0,8 г действующего вещества на 1 м2. Срок обработки — конец фазы выхода в трубку (VI — начало VII этапа органогенеза). Лучшие результаты стерилизации были достигнуты при температуре 18°. Более высокие температуры снижали эффективность препарата. В. М. Сотник [33] рекомендует использование этрела в качестве гаметоцида озимой мягкой пшеницы с учетом разработанной им в данной климатической зоне дозы внесения препарата и сроков обработки районированных сортов.
Однако ряд исследователей [36, 67, 122, 125] на основании полученного экспериментального материала высказывают определенные сомнения в перспективности этрела как гаметоцида для зерновых.
Поиск гаметоцидов, селективно индуцирующих мужскую стерильность основных зерновых культур, остается острой проблемой сельскохозяйственной науки. Он ведется не только в направлении подбора оптимального сочетания дозы и сроков нанесения уже известных гаметоцидов, но .и развивается более интенсивно при выявлении новых веществ, гамето-цидные свойства которых проявляются без сопутствующих не-
З* 35
желательных действий, свойственных этрелу. Поэтому испытание новых химических соединений на гаметоцидную активность проводят параллельно с этрелом, являющимся контролем, по которому судят о наличия или отсутствии у химических соединений, наряду с гаметоцидным эффектом, того или иного негативного действия, свойственного этрелу.
Так, в штате Алабама (США) в течение вегетационного периода 1971—1973 гг. в полевых условиях на гексаплоидных тритикале 6ТА 131, 6ТА 385 и 6ТА 204 были проведены параллельно с этрелом испытания новых соединений, проявивших гаметоцидные свойства — RiH-531 д uniroyal D-513. В опытах 1971—1972 гг. этрел испытывали в концентрациях 100, 250, 500, 1000, 2000 и 4000 мг/кг путем опрыскивания в три срока:
начало, середина и конец фазы выхода в трубку. Так как при этих концентрациях этрела была достигнута недостаточная степень стерильности, в опытах 1972—1973 гг. его применяли в те же сроки, но в более высоких концентрациях — 2500;
5000, 7500 и 10000 мг/кг. Параллельно проводили испытание RH-531 я uniroyal D-513 в концентрациях 1500, 3000 и 6000 мг/кг с применением ПАВ orto-HDD (5 мл на 1л). Обработка высокими дозами этрела, особенно в ранний срок фазы выхода в трубку, оказывала отрицательное влияние на завязывание семян. Реакция трех линий тритикале была различной: для 6ТА 131 концентрация этрела 4000 мг/кг отрицательно действовала на рост и развитие, для 6ТА 385 и 6ТА 204 наиболее неблагоприятной была концентрация 10000 мг/кг [125].
Самыми чувствительными к обработке гаметоцидами были растения тритикале 6ТА 131. Высокий уровень стерильности у них достигался при обработке RH-531 (концентрации 1500 и 3000 мг/кг) перед фазой выхода в трубку или в ее ранний срок. Препарат uniroyal D-513 в данных условиях был наименее эффективным. Ценным качеством обоих препаратов являлось то, что они в меньшей степени, чем этрел, редуцировали завязывание семян. Однако оба препарата, особенно RH-531, вызывали некроз и пожелтение листьев после обработки, способствовали уменьшению длины колоса и высокой плотности упаковки семян в колосе [125].
Действие RiH-531 изучали также на двух сортах яровой пшеницы Anza и Ехора. Одноразовое опрыскивание дозой 2 кг активного вещества на 1 га до начала мейоза вызывало максимум аборт.ивности пыльцы. Сорт Anza был более чувствителен к этому гаметоциду. Дальнейшее испытание препарата необходимо было проводить с учетом его действия на женскую фертильность. В опытах отмечены отрицательные по-
36
бочные действия препарата на рост, развитие растений и женскую фертильность [85].'
На трех сортах озимой пшеницы параллельно с этрелом исследовали влияние препаратов uniroyal D-513, RiH-531, RH-532, RH-2956 и DPX-3778, комбинируя дозы и сроки обработки. Каждый препарат, наряду с гаметоцидньши свойствами, проявлял негативное действие, вызывая хлороз растений (RH-531, RIH-532), повышенную ломкость стеблей и преждевременное их пожелтение (DPX-3778), снижение продуктивности зерна до 60% от контрольного варианта [67]. Это свидетельствует о неперспективности применения данных соединений в качестве гаметоцидов в широких масштабах на зер-новы.х культурах.
•Использование этрела как селективного индуктора мужской стерильности на других сельскохозяйственных культурах -до настоящего времени не дало ощутимых результатов. Этрел применяли для индукции мужской стерильности сахарной свеклы с целью успешной гибридизации между любыми популяциями этой культуры и устранения длительного и дорогостоящего процесса создания аналогов селекционных линий с ЦМС [69]. В зависимости от генотипа этрел индуцировал различную степень стерильности пыльцы сахарной свеклы, но эти же концентрации вызывали фитотокоический эффект, что резко снижало урожайность семян. Поэтому данный гаметоцид едва ли может быть перспективным для этой культуры. При внесении гранулированного этрела в почву под сахарную свеклу также получены негативные результаты [68]. На основании опытов S. С. Phatak [115], который получил 100%-ную стерильность пыльцы у некоторых видов сорняков при внесении гранулированного этрела в почву, была сделана попытка избежать фитотоксического эффекта, вызываемого этрелом при опрыскивании.
,В вегетационных опытах этрел вносили в почву в разные сроки микроспорогенеза: в период премейотической стадии, 'мейоза и после окончания мейоза. Дозы этрела составляли 200, 400, 800 и 1600 мкг/г сухого вещества почвы. Все дозы тормозили рост и развитие растений пропорционально внесенному количеству препарата. При одноразовом применении эт-'рел был более эффективным, чем при внесении той же дозы по частям. Дозы 800 и 1600 мкг/г вызывали гибель всех растений. Двукратное внесение 25, 50 и 70 мкг/г (этрел частично деградирует в течение 15 дней), хотя и способствовало абор-тивности пыльцы, но не могло быть рекомендовано для практического применения из-за сильного фитотоксического действия. Таким образом, этрел пока не оправдал себя как гаметоцид для сахарной свеклы.
37
Однако опыты с этрелом могут служить примером того, как не следует окончательно отказываться от веществ с га-метоцидной активностью, не проявивших себя на основных сельскохозяйственных культурах. Реализация эффекта гетерозиса с помощью этрела была достигнута на растениях семейства Cucurbi'taceae. При опрыскивании листьев растений огурца Cucumis sativus L. растворами этрела в дозе 240 мг/л в 2 приема или 120 мг/л в 4 приема развивались только женские цветки [122].
У растений клещевины этрел не оказывал влияния на изменение пола цветков даже в более высоких дозах, чем те, при которых он вызывал превращение мужских цветков в гер-мафродитные у представителей семейства Cucurbitaceae [51, 84].
Очевидно, не менее важным условием, чем гаметоцидные свойства вещества, является специфика метаболизма культуры, функциональное состояние «первичного места действия» гаметоцида, что определяет способность клетки воспринимать индуцирующее действие гаметоцидного эффекта.
У растений огурца та.кже отмечено формирование пестдч-ных цветков вместо мужских при двукратном опрыскивания раствором этрела в дозе 50 мг/л [44]. У тыквенных образование мужских цветков достигалось путем опрыскивания растений раствором этрела в концентрациях от 50 до 100 мг/л. У дыни концентрация этрела 150 мг/л вызывала изменения в экспрессии пола у 84,6% .мужских цветков [49].
На основе проведенных исследований предполагают получать семена гибридов Cucurbita pepo L. и Cucurbita maxima Duch. на промышленной основе путем индукции мужской стерильности при двукратной обработке растений растворами этрела в концентрации 350 мг/л с учетом температуры и влажности [96].
•В настоящее время среди огромного числа соединений различных классов, обладающих физиологической активностью на растениях, выявлено относительно небольшое количество веществ с гаметоцидным действием. Важнейшими физиологически активными соединениями являются гибберел-лины (ГКз) 'и гетероауксины (2-индолилуксусная, 2-нафтил-уксусная, 2,3,5-трийодбензойная кислоты), соли ;и эфиры 2,4-дихлорфеноксиуксусной кислоты, 4-хлор-2-мет'илфеноксиук-сусной и 2,4,5-трихлорфеноксиуксусной кислот.
Из соединений алифатического ряда гаметоцидную активность проявили 2,3-дихлоризомасляная кислота, 2,3-дяхлор-пропионовая, 2,3,3,3-тетрахлоризомасляная, 2,2,3-трихлормас-ляная, 2,2,3-тpиxлopизolмacлянaя, 2,2-дихлорпроп.ионовая.
38
.Эти 'соединения, применяемые вначале как гербициды, проявили различную степень гаметоцидной активности, будучи апробированными на ряде сельскохозяйственных культур [102, 103, 124]. Среди перечисленных кислот рекомендованы как перспективные гаметоциды: натриевая соль 2,3-дихлор-изомасляной кислоты (фирменное название мендок, или FW-450), кальциевая соль (FW-676) и магниевая соль 2,3-ди-хлоризомасляной кислоты (G-315), 2,2-дихлорпропионовая кислота (далапон). При нанесении водных растворов этих соединений на листья препараты поглощаются тканями листовой пластинки и проникают через проводящую систему к генеративным органам. Опыты с применением радиоактивных изотопов показали, что FW-450 аккумулируется в пыльниках в больших количествах, чем в яйцеклетке [26]. Однако содержание препарата, отмеченное в яйцеклетке (при концентрациях, вызывающих гаметоцидный эффект), достаточно, чтобы вызвать побочные нежелательные действия — редукцию женской фертильности.
Препараты FW-676 и G-315 применяли для стерилизации пыявцы у овса в фазы выхода в трубку и начала колошения. Оба соединения оказались слабоэффективными для этой культуры [116].
'При изучении в полевых условиях возможности химической кастрации растений озимой ржи с помощью FW-450, ГМ)К.'и далапона установлено, что обработка растений FW-450 в концентрации 1% при дозе 2,5 мл на растение в конце фазы кущения и в начале выхода в трубку снижала фертильность пыльцевых зерен без значительного повреждения женских генеративных органов [102, 103]. При обработке растений 0,5°/о-ным раствором далапона в начале фазы выхода в трубку или 0,05%-ным раствором ЛМК в дозе 5 мл на растение в конце фазы выхода в трубку — начала колошения были получены оптимальные результаты для этих соединений. Однако и в данных условиях оба препарата вызывали нарушения в росте и развитии растений и стерилизацию не только мужских, но и женских генеративных органов, что сопровождалось снижением завязывания семян [102, 103]. По-видимому, эти явления связаны не только с концентрацией препарата, дозами растворов и числом опрыскиваний, но и с взаимодействием препарата с объектом обработки.
Z. Natrova [102, 103] считает, что результаты химической кастрации далапоном, FW-450 и ГМК, при указанных дозах;
концентрациях и сроках обработки могут быть использованы для замены ручного кастрирования при скрещивании с целью получения большего количества гибридных семян озимой рж.и, чем теоретически ожидаемые 50% при свободной панмик-39
сии обоих партнеров, а также в случае изучения гетерозиса прд диаллельном скрещивании.
НЕКОТОРЫЕ ОТРИЦАТЕЛЬНЫЕ ЯВЛЕНИЯ ПРИ ИСПОЛЬЗОВАНИИ ГАМЕТОЦИДОВ
Повышенная проницаемость тканей у растений с ЦМС является отрицательным фактором, способствующим проникновению различного рода инфекций. Доказано, что мембраны митохондрий растений кукурузы с цитоплазматически наследуемой мужской стерильностью характеризуются повышенной чувствительностью к патотоксину, выделенному из Helmin-thosporium maydic, раса Т., так как структура и проницаемость мембран митохондрий стерильных растений отличаются от митохондриальных мембран фертильных растений [46, 133]. Гибель стерильного аналога наступала в результате разобщения окислительного фосфорелирования патотоксином в митохондриях, что обусловливало дисбаланс энзиматиче-ских реакций [114]. Указанное явление связывают также с изменениями в мембранах митохондрий с мужской стерильностью и отводят в этом значительную роль белкам внутренней мембраны [63].
Повышенная чувствительность растений с ЦМС к инфекциям и факторам окружающей среды, вероятно, имеет более сложный биохимический базис, и трудность поддержания линий с ЦМС обусловливает необходимость поисков эффективных гаметоцидов, селект.ивно индуцирующих мужскую стерильность без сопутствующих побочных явлений, снижающих урожайность. Одним из нежелательных факторов при использовании рекомендованных в настоящее время гаметоцидов (этрел, ГМ'К, FW-450) является повышенная проницаемость мембран и клеточных стенок, способствующая развитию различного рода инфекций.
Определенная фитотоксичность производных феноксиуксус-ной кислоты и ал.ифатических хлорсодержащих кислот также способствует снижению урожайности, а часто и гибели растения. Под воздействием производных феноксиуксусной кислоты в растительных тканях изменяются соотношения самих фенольных соединений клетки. Разностороннее влияние фё-нольных компонентов в растениях и их участие в регулировании ростовых процессов посредством связи с фитогормона-м.и свидетельствуют о важности этой группы соединений для нормального или абортивного течения процессов спорогенеза. Наиболее чувствительными к воздействию производных феноксиуксусной кислоты и алифатических кислот являются флавоноиды, оксикумарины и бензойные кислоты [II]. Этим,
40
по-видимому, объясняется различная степень токсичности большинства гаметоцидов, относящихся к данным типам соединений, так как накопление свободных фенольных производных (агликонов), обладающих высокой токсичностью, должно инактивироваться в тканях путем образования глю-козидов [42, 136]. Однако низкое содержание моносахаров и высокая активность глюкозидаз при нанесении этого типа препаратов на растения приводят к тому, что у обработанных растений наблюдаются фитотоксические эффекты и степень поражения зависит от чувствительности вида или сорта к этим препаратам [22, 38]. Углеводный обмен у растений с ЦМС изменяется следующим образом. Цитоплазматическая мужская стерильность у растений сопровождается нарушением синтеза и обмена углеводных компонентов, что выражается в низком содержании крахмала (при параллельном угнетении активности амилазы) и значительном увеличении дефицита Сахаров по мере развития микроспор [41, 87].
Соединения, обладающие гаметоцидными свойствами и относящиеся к производным феноксиуксусной кислоты, наряду с гаметоцидным эффектом, как правило, оказывают тормозящее, действие на рост и развитие растения, что негативно сказывается на урожайности [33, 35, 37]. Сумма всех нежелательных .неустранимых до сих пор воздействий этой группы препаратов объясняется скорее всего аддитивным эффектом самого препарата и эндогенных фенольных соединений. Роль фенольных компонентов у обработанных растений изменяется существенным образом. Ауксины и гиббереллины, ответственные за рост и развитие растений, частично или полностью инактивируются в результате блокирования фенольными соединениями их ферментных систем, а эндогенные фенольные ингибиторы, особенно продукты их окисления — хиноны, оказывают непосредственное влияние на ростовые процессы и вызывают фитотоксические эффекты у растений [12, 18, 22].
Поскольку физиологическая активность большинства соединений, обладающих гаметоцидными свойствами, проявляется в меристеме, чувствительность растений к ним должна контролироваться генетически. Установлено, что чем чувствительнее сорт к действию далапона, тем интенсивнее ответная реакция, т. е. метаболические сдвиги в синтезе РНК и белка [13].
Физиологическая активность 2,4Д проявляется в первичных и вторичных меристемах. Интенсивность проникновения 2,4Д-—214C из листьев в стебель и к генеративным органам растений, чувствительных к этому препарату, существенно отличается от устойчивых видов [38]. Быстрая аккумуляция препарата меристемой наблюдается у растений в следующем по-41
рядке: горчица > подсолнечник > фасоль > соя > 'хлопчатник. Слабая аккумуляция меристематической тканью отмечена у культур: тимофеевка < пшеница < огурец < клубника. Эффект препарата (и его фитотоксичность) зависит от подвижности соединения, его иммобилизации и высокой скорости метаболизма вещества в тканях. У устойчивых растений энзиматическое преобразование препарата приводит к нивелированию эффекта его действия. Влияние препарата тем сильнее, чем длительнее его присутствие в тканях в неизменном виде. Этим объясняется широкая шкала доз гаметоцидов не только для различных культур, но и в пределах определенной культуры в зависимости от сортовых особенностей. Так, этрел расщепляется в тканях, имеющих слабокислую реакцию [47].
При сравнении действия 2,4Д на относительно чувствительную яровую пшеницу (сорт Лютесценс 758) и яровой ячмень (сорт Винер), отличающийся устойчивостью к хлорфе-ноксикислотам, установлено, что при обработке в одну и ту же фазу (5—6-го листа) у пшеницы наблюдалось угнетение роста, сопровождающееся частичным повреждением растений, у ячменя та же доза (0,7 кг/га) стимулировала рост и развитие растений [10].
В опытах по индукции мужской стерильности у проса обнаружена сортовая специфика при воздействии 2.4Д [40]. Скороспелые сорта при морфологически одинаковой фазе развития по сравнению с позднеспелыми сортами обладали иным этапом органогенеза. Эффективность гаметоцида зависела не только от концентрации и наносимой дозы, сорта и фазы развития в момент обработки, но и от погодных условий. Поэтому, наряду с основной концентрацией, рекомендуется применять две смежные. Кроме сортовых показателей, высокогете-розисное растение обладает защитным действием против химической обработки [361.
Отрицательным явлением при использовании гаметоцидов является .их высокая биологическая активность, что резко сужает практический диапазон концентраций препаратов. Кроме хорошо изученных фенольных соединений, производные тиокарбаминовой кислоты, диазинов, анилинов при рекомендуемых для борьбы с сорняками дозах вызывают нарушения в мейозе и гаметогенезе, в результате чего отмечена частичная или полная стерилизация пыльцы у диплоидных растений сахарной свеклы. Генеративные органы тетраплоидов отличались большей устойчивостью к действию этих препаратов Г39].
Все исследователи, работающие с гаметоцидами, пришли к выводу, что эффективность любого вещества с гаметбцид-
42
ными свойствами зависит от генотипа, окружающих -условий, дозы и концентрации препарата, этапа органогенеза, времени обработки, а также числа обработок. В экспериментах, проведенных в вегетационном домике с растительным материалом, выравненным по развитию, при контролируемых условиях опыта, оптимальных и константных условиях внешней среды получают очень хорошие результаты, которые в полевых условиях трудно повторить.
ЗАКЛЮЧЕНИЕ
В настоящее время в основном известны изменения метаболических процессов, происходящие в растительном организме в результате воздействия растворов химических соединений, обладающих гаметоцидной активностью. При этом эффект гаметоцидов по своему действию на проницаемость клеточных мембран аналогичен эффекту стерилизующей цитоплазмы при перемещении в нее ядра, т. е. применение этих препаратов вызывает мужскую, а порой — и частично женскую стерильность. Аналогичные явления наблюдали при изучении мужских стерильных аналогов сортов пшеницы на основе цитоплазмы Aegilops ovata L. и Ае. caudata L.
Для создания мужских стерильных (кастрированных) растений с целью получения гибридных семян первого поколения выявлены и предложены для исследований некоторые химические соединения — этрел (этефон), ГМК, нафтилуксусная кислота .и др. При испытании этих соединений в различных странах мира прежде всего с целью разработки некоторых элементов технологии их использования и определения эффективности не было получено 100%-ной стерилизации мужского гаметофита при обработке посевов в период выхода в трубку, а также в другие фазы развития. Кроме того, применение указанных гаметоцидов задерживало прохождение фенологических фаз у растений, а также выколашивание части стеблей.
Однако, несмотря на отрицательные побочные действия гаметоцидов, применение их в ближайшей перспективе может открыть путь к реализации эффекта гетерозиса у зерновых (особенно пшеницы), технических, овощных и кормовых культур. Поэтому весьма важно продолжать поиски новых веществ с гаметоцидньши свойствами и выявлять их эффективность путем тщательного изучения доз и сроков их применения для каждой культуры и сорта в определенной природно-климатической зоне.
43
Л ИТЕРАТУРА
1!. Анащенко А. В. Достижения и перспективы селекции подсолнечника. М., ВНИИТЭИСХ, 1977, 53 с.
2. Б а л и н а Н. В. Действие повышенных температур на рост пыльце-.вых трубок. — Физиология растении, 1976, т. 23, № 4, с. 805—8Я1.
3'. Барская Е. И., Балина Н. В. О роли каллозы в пыльниках растении. — Физиология растении, 1977, т. 18, № 4, с. 716—721.
4. Барская Е. И., Балина Н. В., .К а наш Е. В. Влияние засухи на динамику каллозы в пыльниках растений. — Доклады АН СССР, 1973 .т. 208, № 2, с. 476—478.
5. Батыгина Т. Б. Эмбриология пшеницы. Л., «Колос», 1974, 206 с.
6. Борисенко Л. Р., Дмитриева А. Н. Окислительно-восстановительные процессы у форм пшеницы с цитопаазматической мужской стерильностью. — Селекция и семеноводство. .Киев, «Урожай», 1975, вып. 30, с. 69-75.
7. Брежнев Д. Д., Шмарае® Г. Е. Селекция растений в США. М., «Колос», 1972, 296 с.
8. Б р и т и к о в Е. А. Биологическая роль пролина. М., «Наука», 1975, 87 с..
9. Буш Г., Кнехт М., О л сон М., С т арбу к В. К. Современное состояние исследований белков ядрышка. — В кн.: Клеточное ядро, мор-фолотия, физиология, биохимия. М., «Наука», 1972, с. 99—1!12.
10. Воеводин А. В., Н е в з о р о в а Л. И. Различия в действии 2,4Д и симазина на фосфорный обмен гороха и некоторых злаков. — Ме-.ханиэм действия гербицидов и синтетических .регуляторов и их судьба в биосфере. Х Международный симпозиум. Пущино. Издательство АН СССР, '1975, ч. 1, с. 67—70.
М. Вольте ц А. П., Пальченко Л. А. О взаимодействии гербицидов с фитогормонами. — Доклады АН БССР, 1972, т. 16, № 10, с. 930— 933.
'12. Волы не ц А. П., Пальченко Л. А. Изменения фенольного комплекса растении при воздействии гербицидами. — Механизм действия гербицидов и синтетических регуляторов и их судьба в биосфере. — Х Международный симпозиум. .Пущино. Издательство АН СССР, 1975, ч 1 с. 97—100.
J3. Деева В. П., Шелег 3. И. О механизме действия .'производных алифатических 'карбоновых кислот. — Механизм действия гербицидов и синтетических регуляторов и их судьба в биосфере. Х Международный симпозиум. Пущино. Издательство АН СССР, 1975, ч. 1, с. 60—64.
14. Дельштедт Р., Хюбнер Г., Хиршберг К. Связывание ауксинов с белками мембран проростков. — Механизм действия гербицидов и синтетических регуляторов роста растений и их судьба в биосфере. Х Международный симпозиум. Пущино. Издательство АН СССР 1975 ч. 1,_с. 54—57.
15. Дмитриева А. Н., Борисенко Я. 'В. Некоторые биохимические особенности ЦМС и восстановление фертильности у пшеницы. — Вестник сельскохозяйственной науки Украинской академии сельскохозяйственных наук. Харьков. Издательство АН УССР, 1973, № 1, с. 39—40.
44
16. Дмитриева А. Н., Хавжинская О. Е. Биохимические особенности растений сорго с ЦМС. — В кн.: Селекция и .семеноводство. Киев. «Урожай», !1975, вып. 30, с. 30—38.
'17. Д у л и н А. Ф. Влияние регуляторов роста на процесс фотофосфо-релирования. — Механизм действия гербицидов и синтетических регуляторов роста растений и их судьба в биосфере. Х Международный симпозиум. Пущино. Издательство АН СССР, 1975, ч. 1, с. 89—92.
КЗ.-Змртал 3., Махачкова А. Роль пероксидазы в метаболизме фенольных соединений и индолилуксусной кислоты. — Механизм действия гербицидов и синтетических регуляторов роста растений и их судьба в биосфере. Х Международный симпозиум. Пущино. Издательство АН СССР, 1975, ч. 1, с. 109—113.
•19. 'Калинин Ф. Л., Борейко В. К. Новые данные о механизме действия гидразида малеиновй кислоты. — Механизм действия гербицидов и синтетических регуляторов роста растений и их судьба в биосфере. Х Международный симпозиум. Пущино. Издательство АН СССР, 1975, ч. 1, с. 9—12.
'20. Кал1н1н Ф. Л., MixHO А. 'М. Бюсинтез пантотеново! кислоти в чолов1чих репродуктивних органах кукурудзи при стерилгзацп. — Нау-ков1 пращ!. КиГв, 1971, вью'. 50, с. 50—52.
21. Ладонин В. Ф., Пронина Н. Б. Действие 2,4Д на активность аденилаткиназы и метаболизм адениннуклеотидов в растениях гороха и ячменя в связи с условиями фосфорного питания. — Механизм действия гербицидов и синтетических регуляторов роста растений и их судьба в биоафере. Х Международный симпозиум. Пущино. Издательство АН ССОР, 1975, ч. 1, с.'47—49.
22. Лама н Н. А. Влияние 2,4Д на качественный состав флавоноидов люпина желтого. — Механизм действия гербицидов и синтетических регуляторов роста растений и их судьба в биосфере. Х Международный симпозиум. Пущино. Издательство АН СССР, 1975, ч. 1, с. 117—120.
'23. Либерман Е. А., Топалы В. П., 3 и л ь'б ер ш т е й.н А. Я., О х л о б ы с т и н О. Ю. Подвижные переносчики ионов и отрицательное сопротивление мембран. 1. Разобщители окислительного фосфорелирова-ния — переносчики протонов. — Биофизика, 1971, т. ;16, № 4, с. 615—625.
124. Либерман Е. А., Цофина Л. М., Арчаков А. И., Д е в и-че некий В. М., Карузина И. И., Карякин А. В. Липофильные анионы — новый класс ингибиторов реакций гидроксилирования. — Доклады АН СССР, 1974, т. 218, № 5, с. 1239—1241.
25. Л и х о л а т Т. В., Поспелов В. А., Морозова Т. М., С а л -га ни к Р. И. О механизме действия ауксина на клетки разного возра-.ста. — Механизм действия гербицидов и синтетических регуляторов роста растений и их судьба в биосфере. Х Международный симпозиум. Пущино. Издательство АН СССР, 1975, ч. 1, с. 112—116.
26. М i х н о А. М. НуклеТнов! кислоти та азотов! сполуки в суцв1ттях 'кукурудзи при х1м1чн1й стерил1зац11. — Науков! пращ УСГА, 1971, вып. 40, с. 54—56.
27. Палилова А. Н. Цитоплаэматическая мужская стерильность у растений. Минск. «Наука и техника», 1969, 209 с.
'28. Поддубная-Арнольди В. П. Цитоэмбриология покрытосеменных растений. М., «Наука», 1976, с. 68—142.
29. Ракитин Ю.В., В л а д им ир ц е в а -С. В., Николаева Л. А. Действие гидразида малеиновой кислоты на обмен РНК и белка в почках картофеля. — Механизм действия гербицидов и синтетических регуляторов роста растений и их судьба в биосфере. Х Международный симп'ози-ум. Пущино. Издательство АН СССР, 1975, ч. 1, с. 7—9.
0 комментариев