Навигация
Минимальные последовательности, необходимые для начала правильной транскрипции (промотор) и для образования правильного 3`-конца зрелой РНК
27450
знаков
0
таблиц
0
изображений
2. Минимальные последовательности, необходимые для начала правильной транскрипции (промотор) и для образования правильного 3`-конца зрелой РНК
3. Последовательности, регулирующие частоту инициации транскрипции; к ним относятся последовательности, ответственные за индуцибельность и репрессию транскрипции, а также клеточную, тканевую и временную специфичность транскрипции. к их числу относятся энхансеры и сайленсеры – последовательности, которые оказывают дистанционное влияние на инициацию транскрипции независимо от своей ориентации относительно точки начала транскрипции.
Для инициации транскрипции с участием трех разных РНК-полимераз используются разные регуляторные последовательности, при этом последние располагаются каждая на определенном расстоянии от точки начала транскрипции. Кроме того, для РНК-полимеразы каждого типа требуются свои вспомогательные белки (факторы транскрипции), которые связываются с этими регуляторными последовательностями.
Для включения и выключения транскрипции эукариотических структурных генов используется множество разнообразных высокоспецифичных процессов. Многие из факторов транскрипции связываются непосредственно с нуклеотидной последовательностью длиной менее 10 п.н., называемые по-разному: боксом, модулем, элементом инициации, регуляторным элементом. В отличие от прокариот у эукариот опероны в большинстве своем отсутствуют, т.е. каждый эукариотический структурный ген имеет свой собственный набор регуляторных элементов. Существенную роль в регуляции транскрипции у эукариот играет также белок-белковое взаимодействие.Несмотря на индивидуальность набора регуляторных элементов у структурных генов эукариот, каждый из них имеет промоторный участок (ТАТА-бокс или бокс Хогнесса) из восьми нуклеотидов, включающий последовательность ТАТА; последовательность ЦЦААТ (ЦАТ-бокс); участок из повторяющихся динуклеотидов ГЦ (ГЦ-бокс). Эти элементы находятся на расстоянии 25, 75 и 90 п.н. от сайта инициации соответственно (прозрачка 21). Транскрипция структурного гена эукариот начинается со связывания с ТАТА-боксом фактора транскрипции IID (TFIID), который представляет собой комплекс из примерно 14 белков. Затем с TFIID и участками ДНК примыкающими к ТАТА-боксу связываются другие факторы транскрипции и, наконец, со всем этим транскрипционным комплексом связывается РНК-полимераза II. Затем при участии дополнительных факторов происходит инициация транскрипции в точке +1 (прозрачка 19). Ясно, что если последовательности ТАТА отсутствует, или сушественно изменена, то транскрипция структурного гена становится невозможной. Идентифицированы также факторы транскрипции специфичные для регуляторных элементов ЦЦААТ и ГЦ, но пока не ясно, как ДНК-белковые взаимодействия могут влиять в этом случае на эффективность транскрипции, если элементы расположены на расстоянии более 75 п.н. от сайта инициации. Кроме того, на расстоянии сотен и даже тысяч пар оснований от сайта инициации находится так называемая энхансерная последовательность, которая многократно повышает скорость транскрипции структурных генов. По-видимому, сближение удаленных регуляторных элементов и соответствующего структурного гена происходит при укладке хромосомной ДНК. Кроме того, факторы транскрипции, которые связываются с определенными энхансерами и регуляторными элементами, могут образовывать цепочку, соединяющую удаленные друг от друга сайты.
Некоторые репрессированные (не эксперессирующиеся) гены активируются каскадом событий, который запускается каким-либо специфическим внеклеточным сигналом, например, повышением температуры, или синтезом гормона. Гормон, поступив в кровоток, связывается с рецепторами специфических клеток, облегчающими его проникновение в клетку. Оказавшись в клетке, гормон вступает во взаимодействие с одним из клеточных белков и изменяет его конформацию. В таком измененном состоянии белок проникает в ядро и связывается со специфическим регуляторным элементом, который инициирует транскрипцию соответствующего гена.
Существуют также белки, которые, взаимодействуя с регуляторными элементами блокируют транскрипцию. Например, известен класс генов позвоночных (примерно 18), активно транскрибирующихся только в нервных клетках. Каждый их этих генов имеет регуляторный элемент из 24 п.н., находящийся «левее» сайта +1; он обозначается NRSE (от англ. Нейрон рестректив. Силенцер. элемент). Во всех клетках, кроме нейронов, синтезируется NRSF-фактор, который связывается с NRSE и блокирует транскрипцию соответствующих генов. В нейронах NRSF не синтезируется, и упомянутые гены активно транскрибируются.
Таким образом, структурный ген может иметь множество регуляторных элементов, которые активируются специфическими сигналами в клетках разного типа в разное время клеточного цикла. В синтезе РНК также выделяют стадии инициации, элонгации и терминации, но в этих процессах часто принимают участие другие ферменты и последовательности оснований, чем у прокариот, однако первыми основаниями, включаемыми в РНК при инициации, являются, как и у прокариот, А или Г.
Похожие работы
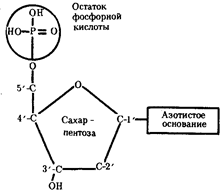
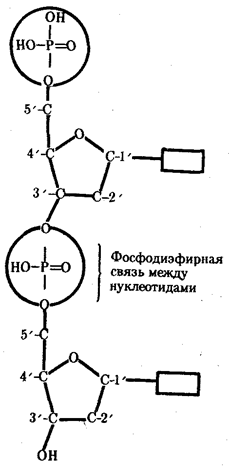
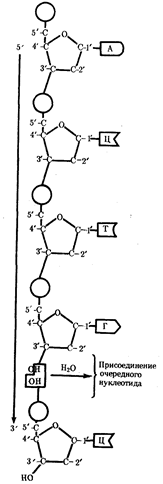
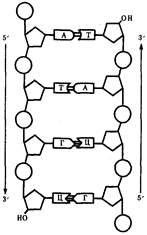
... : генный, хромосомный и геномный. На каждом из них проявляются основные свойства материала наследственности и изменчивости и определенные закономерности его передачи и функционирования. 4. Генный уровень организации генетического аппарата Элементарной функциональной единицей генетического аппарата, определяющей возможность развития отдельного признака клетки или организма данного вида, ...
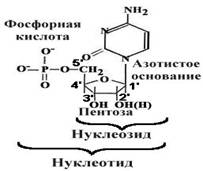
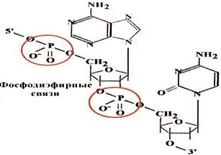
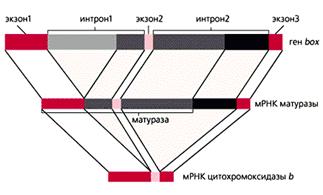

... гены резистентности к антибиотикам, перемещаясь из одной клетки в другую в процессе коньюгации, гены резистентности быстро распространяются в популяции бактерий). Клетки эукариот используют в качестве генетического материала лишь двуцепочечную ДНК. Структурные гены в них подразделяются на независимые гены (их транскрипция не связана с транскрипцией других генов, их активность регулируется, ...
0 комментариев