Навигация
Стабильная температурная регуляция у рептилий
47229
знаков
0
таблиц
0
изображений
2.4 Стабильная температурная регуляция у рептилий
В сравнении с амфибиями рептилии преимущественно являются жителями суши. У всех видов рептилий существуют сходные формы размножения, в том числе и у вторичных водных жителей, преимущественно живущих в воде. Яйцо покрыто скорлупой и располагается на суше под солнцем, где и высиживается - у некоторых видов рептилий также за счет дополнительного тепла тела. Во время эмбрионального развития возникает оболочка яйца, которая функционирует как орган дыхания и обмена и обеспечивает процесс размножения при относительно сильных колебаниях условий окружающей среды.
Первые рептилии появились от 310 до 250 миллионов лет назад в карбоновом и в пермском периоде. Высший пункт их развития проявился могущественным разнообразием форм и, как следствие этого, они заполнили все жизненные пространства - воздух, сушу и воду.
Из 16 отрядов рептилий, которые известны по ископаемым находкам, до современности сохранились только пять. Сейчас известно около 3000 видов ящериц, 2750 видов змей. 220 видов черепах, 21 вид крокодилов, а также отряд клювоголовых. Все современные рептилии располагают эффективными и хорошо функционирующими системами температурной регуляции.
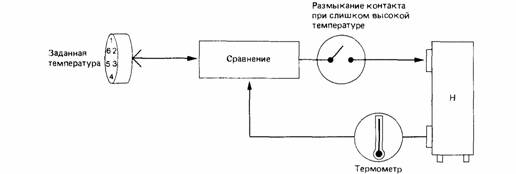
Терморецепторы у рептилий располагаются в коже, во внутренних органах и в мозге. Раздражение их естественными или экспериментальными стимулами вызывает координированные терморегуляторные реакции. В 1967 году впервые удалось имплантировать в ствол мозга полосатого сцинка (Eumeces fasciatus) - ящерицы с длиной тела более 60 см - термоды и микроэлектроды. Если через термоды, по которым могла протекать холодная или теплая вода, терморегуляторную область мозга охлаждали, то животные, у которых была возможность выбора между теплой и холодной частями камеры, быстро искали более теплое пространство. Животные, которые находились в теплой части камеры, при охлаждении их ствола мозга, покидали теплое пространство и перебирались в более прохладную часть камеры.С помощью микроэлектродов удалось зарегистрировать активные потенциалы отдельных нервных клеток ствола головного мозга. Установлено, что нагревание головного мозга увеличивает активности тепловых нейронов. Охлаждение увеличивает активность Холодовых нейронов.
У некоторых видов змей, таких как боа (Constrictor sonstrictor) и гремучая змея, между носовым отверстием и глазом существует парный орган - орган ямки, в котором располагаются высокочувствительные терморецепторы. Орган состоит из углубления, которое с помощью мембраны толщиной от 10 до 15 мкм, делится на верхнюю и нижнюю половины. Через узкое отверстие верхней камеры инфракрасные тепловые лучи могут поступать в полость ямки и нагревать мембрану. Теплоемкость мембраны чрезвычайно мала. Поэтому минимальный приход тепла ведет к температурным изменениям, что активирует терморецепторы мембраны. Это происходит тогда, когда порог температурных изменений составляет около 0,003°С при продолжительности раздражения в пределах 0,06 секунды.
Падение тепловых лучей отвесно через отверстие, максимально раздражает температурные рецепторы. Каждое отклонение от перпендикуляра изменяет силу раздражения, так что по направлению тепловых лучей можно определить их источник.Высокая восприимчивость температурных рецепторов и способность к определению направления и источника инфракрасных лучей позволяют змеям и днем и ночью находить добычу, температура тела которой выше, чем температура окружающей среды.
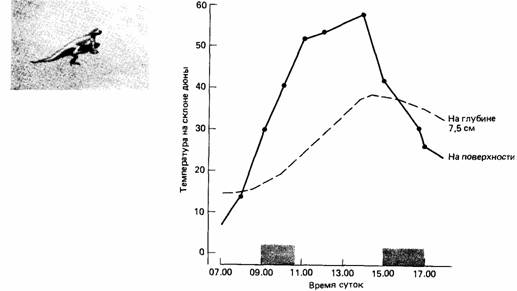
За счет терморегуляторного поведения многие рептилии во время бодрствования сохраняют стабильную температуру тела. Особо следует выделить терморегуляторное поведение крокодилов. В зависимости от окружающей температуры они принимают в воде различные положения. При низкой температуре воздуха, но высокой температуре воды они полностью погружаются в воду. Высовывая из воды голову или спину, крокодилы способны с помощью терморецепторов кожи спины измерить температуру воздуха и интенсивность солнечного излучения (инсоляцию). Если температура воздуха выше температуры воды, то животные ищут освещенное солнцем место на берегу. За счет выбора более или менее влажного грунта происходит изменение теплоотдачи. В условиях интенсивной инсоляции температура тела возрастает выше оптимального уровня, что заставляет животных искать тенистое место или опускать хвост в воду, а при увеличении тепловой нагрузки и другие части тела, что позволяет животным поддерживать постоянной температуру тела, так как известно, что тепловое содержание слагается из теплоотдачи и теплопродукции.
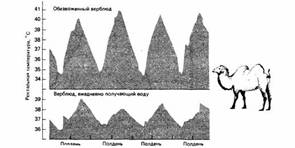
У рептилий наряду с терморегуляторным поведением имеют значение другие физиологические механизмы температурной регуляции. Большие рептилии обладают в три-пять раз меньшим энергообменом, чем маленькие рептилии при схожей температуре тела, и даже в 10 раз меньшим энергообменом, чем птицы или млекопитающие с аналогичной массой тела. За счет акклиматизации к высоким или низким температурам окружающей среды энергообмен рептилий может значительно измениться. Животные, акклиматизированные к 35°С, имеют по сравнению с ящерицами, у которых температура акклиматизации составляет 15°С, при наружных температурах в диапазоне от 15 до 40°С, крайне низкий уровень теплопродукции. Теплопродукция акклиматизированных к теплу животных при 40°С такая же, как и акклиматизированных к холоду ящериц при температуре 30°С; подобная теплоустойчивость существенно увеличивает шансы выживания рептилий при высоких температурах.
В принципе считается, что у рептилий, как и у всех эндодермных животных, энергообмен снижается при снижении температуры окружающей среды. Исключение из этого правила известно уже многие десятилетия. Питоны, которые путем перекатывания закапывают в песок свои яйца, за счет ритмических сокращений мускулатуры увеличивают свою теплопродукцию и повышают температуру тела в отличие от температуры окружающей среды даже в тени от 4 до 5°К (т.е., выше температуры воздуха). Например, питон, высиживающий яйца, имеет при температуре окружающей среды в 25,5°С, на 930% более высокий энергообмен, чем его собрат по виду, не высиживающий яйца. За счет сильных сокращений мускулатуры змеи становятся фактически эндотермными. Этот пример показывает непрерывную связь перехода эктотермии в эндотермию.
Многие рептилии ускоряют при высокой внешней температуре свою дыхательную деятельность (по аналогии с млекопитающими и птицами). За счет увеличенного дыхательного объема вода испаряется более интенсивно с поверхности широко открытого пространства пасти, отнимая тепло от слизистых оболочек полости рта. Особенно эффективен этот процесс у крокодилов, варанов и ящериц больших размеров. Подобным образом они снижают в жару температуру своего тела в течение нескольких часов ниже температуры окружающей среды.
Похожие работы
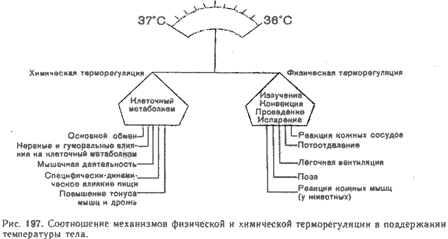
... образом, понижение окружающей температуры, а следовательно, изменение характера температурного воздействия на экстерорецепторы уменьшает температурную чувствительность гипоталамуса. Участие гипоталамуса в терморегуляции обеспечивает взаимодействие восприятия сигналов об изменении температуры окружающей и внутренней среды. Именно в гипоталамусе расположены основные центры терморегуляции, которые ...
... предположение о прямом слежении за этими веществами и их гомеостатическом контроле, но соответствующих данных недостаточно. 5. Мотивационные системы Требования гомеостаза, как мы видели, ставят определенные задачи перед поведением животных. В каждый данный момент животное должно оценивать свое внутреннее состояние, добавлять к этому свои знания о вероятных будущих нуждах и о тех новых ...
... волокон типа А (дельта) и С, локализованы в коже и внутренних органах. Существуют и центральные, локализованные в гипоталамусе, терморецепторы. Кожные терморецепторы реализуют передачу в центры терморегуляции сигналов об изменениях температуры среды, а также обеспечивают формирование температурных ощущений. Число холодовых рецепторов кожи во много раз превышает число тепловых рецепторов. Во ...
... функция дыхания. Интенсивность подъема температуры зависит от 2-х факторов патогенеза: 1. От количества эндогенных пирогенных веществ (интерлейкина 1, например) 2. От индивидуальной чувствительности центра терморегуляции (хорошо лихорадят кролики, плохо крысы - то пример видовой специфичности чувствительности центра терморегуляции). Вторая стадия лихорадка - после подъема температуры, ...
0 комментариев