Транспорт через мембрану клетки
Содержание
Введение
Натрий-калиевый обменный насос
Биохимические свойства натрий-калиевой АТФазы
Кальциевые насосы
АТФазы зндоплазматического и саркоплазматического ретикулумов
АТФазы плазматической мембраны
Натрий-кальциевый обменник
Транспортные системы натрий-кальциевого обмена
Реверсия направления работы NCX
Натрий-кальциевый обменник в палочках сетчатки
Литература
Введение
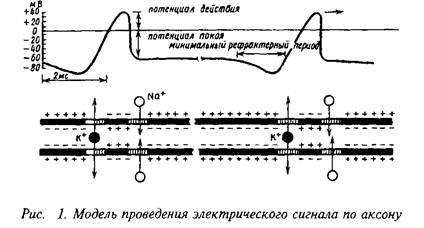
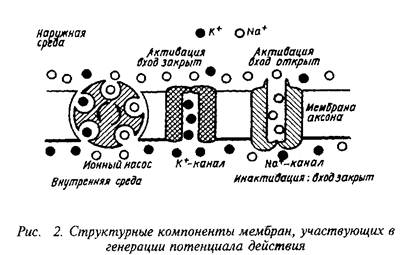
Вход и выход ионов через каналы в мембране нейрона имеет пассивный характер и происходит благодаря наличию электрических и химических градиентов. Для компенсации результатов передвижения ионов клетка использует активные транспортные механизмы, которые затрачивают энергию на перемещение ионов в направлении, противоположном их электрохимическим потенциалам. Таким образом, концентрации ионов в цитоплазме поддерживаются на постоянном уровне, что позволяет сохранить неизменным потенциал покоя, а также генерировать электрические сигналы.
Первичный активный транспорт осуществляется за счет энергии гидролиза АТФ. Наиболее распространенный пример такого транспорта - натрий-калиевый обменник, или насос. Специальная молекула, называемая натрий-калиевой АТФазой, осуществляет за счет энергии расщепления одной молекулы АТФ перенос трех ионов натрия наружу и двух ионов калия внутрь клетки. Поскольку в результате каждого транспортного цикла происходит изменение суммарного трансмембранного заряда на единицу, натрий-калиевый насос является электрогенным, то есть производит электричество.
Другой пример активного ионного транспорта - АТФазы, выводящие кальций из цитоплазмы: кальциевые АТФазы плазматической мембраны выкачивают кальций за пределы клетки, а АТФазы эндоплазматического и саркоплазматического ретикулумов закачивают кальций из цитоплазмы во внутриклеточные структуры.
Вторичный активный транспорт основан на энергии передвижения ионов натрия в направлении их электрохимического градиента. При этом другие ионы переносятся за счет движения ионов натрия либо в том же (ко-транспорт), либо в обратном направлении (ионообмен). Примером такого механизма является натрий-кальциевый обменник, выводящий один ион кальция за счет входа в клетку трех ионов натрия. Как и все системы активного транспорта, этот обменник обратим и может работать как в прямом, так и в обратном направлении, в зависимости от соотношения электрических и химических градиентов для обоих ионов. Вторая система натрий-кальциевого обмена встречается в клетках сетчатки и осуществляет перенос одного иона кальция и одного иона калия наружу, в обмен на четыре входящих иона натрия. Энергия входа натрия в клетку используется также для переноса ионов хлора и бикарбоната через клеточную мембрану. Все вышеперечисленные механизмы основаны на передвижении натрия в направлении его электрохимического градиента и, следовательно, зависят от эффективности работы натрий-калиевого насоса, обеспечивающего поддержание этого градиента.
Транспорт медиаторов необходим для функционирования нейронов. Накопление молекул медиатора в синаптических пузырьках (везикулах) в цитоплазме пресинаптического окончания невозможно без такого транспорта, основанного на перемещении ионов (ионно-сопряженный транспорт). Подобный же механизм используется для обратной закачки медиатора после его выброса в синаптическую щель.
Натрий-калиевый обменный насос
Большинство возбудимых клеток имеют потенциал покоя от - 90 до - 60 мВ. Равновесный потенциал для ионов натрия (ENa) - обычно порядка +50 мВ. Таким образом, существует большой электрохимический потенциал, стремящийся перенести ионы натрия внутрь клетки. Такой перенос осуществляется при помощи многочисленных механизмов. Кроме того, равновесный потенциал для калия более отрицателен, чем потенциал покоя, вследствие чего ионы калия постоянно выводятся из клетки. Для поддержания жизнеспособности клетки необходимо, чтобы ионы натрия непрерывно переносились наружу, а ионы калия внутрь клетки, т.е. против их электрохимических градиентов. Для этой цели в мембране клетки существует натрий-калиевый обменный насос, который при каждом своем цикле переносит три иона натрия наружу и два иона калия внутрь клетки.
Уже ранние эксперименты Ходжкина и Кейнеса на гигантском аксоне кальмара убедительно показали, что источником энергии для этого процесса является расщепление АТФ. В то же время Скау продемонстрировал, что АТФаза, изолированная из нерва краба, обладает многими из биохимических характеристик, которые должен иметь натрий--калиевый насос. Действительно, как для работы насоса, так и для активации АТФазы необходимо присутствие ионов натрия и калия. Кроме того, уабаин (ouabain) одинаково блокирует активность насоса и АТФазы.
На основе этих наблюдений был сделан вывод о том, что натрий-калиевая АТФаза и есть натрий-калиевый насос. Она принадлежит к семейству АТФаз Р-типа (P-type), получившему свое название потому, что в процессе работы эти АТФазы образуют фосфорилированную форму. Перенос ионов осуществляется за счет энергии расщепления АТФ. К этому же семейству относятся кальциевые АТФазы, выводящие ионы кальция из цитоплазмы клетки, и протон-калиевые АТФазы, наиболее примечательная функция которых заключается в секреции большого количества кислоты в полость желудка.
Биохимические свойства натрий-калиевой АТФазы
Биохимические свойства натрий-калиевой АТФазы хорошо известны уже на протяжении многих лет. Стехиометрическое соотношение связываемых катионов совпадает со свойствами транспортного процесса: в среднем, на каждую расщепляемую молекулу АТФ переносится три иона натрия и два иона калия. Избирательность к ионам калия весьма высока: это единственный субстрат, переносимый насосом наружу клетки, и единственный моновалентный катион, не принимаемый насосом для переноса внутрь. Например, литий, аммоний, рубидий, цезий и таллий могут заменить калий во внеклеточном растворе, но не натрий во внутриклеточном. В отсутствии калия насос переносит натрий гораздо менее эффективно (около 10% мощности).
Специфическим блокатором натрий-калиевой транспортной системы являются вещества, используемые при лечении сердечнойнедостаточности (digitalis glycosides), в особенности уабаин и строфантидин. Блокируя активный транспорт натрия и калия, эти вещества не оказывают воздействия на пассивное перемещение ионов через ионные каналы в мембране.
Кальциевые насосыИзменение концентрации ионов кальция внутри клетки играет важнейшую роль во многих процессах жизнедеятельности нейронов, таких как высвобождение медиатора в синаптическую щель, активация ионных каналов в клеточной мембране, а также регуляция целого ряда цитоплазматических ферментов.
В мышечных клетках кальций играет ключевую роль в запуске процесса сокращения мышечного волокна. Все эти функции связаны с кратковременным повышением концентрации кальция в цитоплазме, поэтому важной задачей для клетки является поддержание неизменного уровня кальция в покое. В противном случае различные кальций - зависимые механизмы будут активироваться не в ответ на специфическое раздражение, а постоянно.
Изменение концентрации кальция в цитоплазме может происходить по двум причинам: кальций может входить или выходить через клеточную мембрану, либо переходить из цитоплазмы во внутриклеточные органеллы и обратно, в первую очередь в эндоплазматический ретикулум (в мышце - саркоплазматический ретикулум) и митохондрии. Для измерения внутриклеточной концентрации кальция в клетку вводятся специальные вещества, такие как экворин (aequorin) или фура-2 (fura2), которые излучают или поглощают свет при связывании ионизированного кальция. Другой способ отслеживания изменений уровня кальция - трансфекция особых белковых комплексов, созданных при помощи генной инженерии таким образом, что их флуоресцентные свойства изменяются в зависимости от концентрации ионизированного кальция. В обоих случаях изменения в поглощении или излучении, пропорциональные изменениям уровня кальция, измеряются с помощью высокочувствительных оптических методов. Средняя концентрация кальция в покое для большинства нейронов составляет от 10 до 100 нМ. Уровень кальция в межклеточном пространстве позвоночных составляет от 2 до 5 ммоль.
Для поддержания низкой внутриклеточной концентрации кальция необходим механизм, осуществляющий непрерывный вывод кальция из клетки вопреки наличию значительного концентрационного градиента. Кроме того, системы кальциевого транспорта через внутриклеточные мембраны поддерживают высокую концентрацию кальция в органеллах. Так, уровень кальция в эндоплазматическом ретикулуме может достигать 400 мкмоль, а в саркоплазматическом ретикулуме мышцы поднимается до 10 ммоль. Молекула, ответственная за транспорт кальция через плазматическую и цитоплазматическую мембраны, называется кальциевая АТФаза. Еще один механизм транспорта кальция будет обсуждаться ниже в этой главе.
АТФазы зндоплазматического и саркоплазматического ретикулумов
Одно из семейств АТФаз расположено в мембране эндоплазматического ретикулума нейронов, а также в саркоплазматической мембране скелетной мышцы. Эти АТФазы переносят ионы кальция из цитоплазмы во внутриклеточные органеллы. Сокращение мышечного волокна происходит при освобождении кальция из саркоплазматического ретикулума в миоплазму. Быстрое устранение ионов кальция из миоплазмы, необходимое для релаксации мышцы, обеспечивается за счет высокой концентрации АТФаз в мембране саркоплазматического ретикулума.
Кальциевый транспортный цикл в принципе аналогичен циклу работы натрий-калиевого насоса. Он начинается с присоединения двух ионов кальция к местам связывания, расположенным в цитоплазме и обладающим высоким сродством к кальцию (Km (Ca) ~ 100 нМ) Затем происходит фосфорилирование фермента и изменение его конформации, в результате чего ионы кальция переносятся внутрь ретикулума. После освобождения кальция молекула АТФазы дефосфорилируется и возвращается в свое первоначальное состояние.
АТФазы плазматической мембраныКальциевые АТФазы встречаются также в плазматической мембране любой клетки. За исключением некоторых деталей, строение и функция этих АТФаз не отличается от кальциевых АТФаз эндоплазматического и саркоплазматического ретикулумов. Внутриклеточное место связывания обладает высоким сродством к кальцию, однако во время транспортного цикла происходит связывание всего одного иона кальция. Концентрация АТФаз в плазматической мембране нейронов и мышечных клеток довольно низка, поэтому эффективность этой транспортной системы не слишком высока. Тем не менее, с задачей устранения входящего в клетку кальция она справляется.
Натрий-кальциевый обменникВо многих механизмах ионного транспорта используется совершенно иной принцип переноса ионов через мембрану против электрохимического градиента. Вместо энергии расщепленной молекулы АТФ эти механизмы используют энергию уже существующего перемещения ионов натрия в направлении их концентрационного градиента, то есть внутрь клетки.
Один из примеров - натрий-протоновый обменник, переносящий ионы в соотношении 1: 1 и участвующий в поддержании внутриклеточного рН. Протоны переносятся из клетки наружу вопреки электрохимическому градиенту в обмен на ионы натрия, перемещаемые внутрь клетки.
Подобным же образом переносятся кальций, калий, бикарбонат и хлор. В результате работы этих вторичных транспортных механизмов, в состоянии покоя внутрь клетки попадает значительное количество натрия. Тем более важна роль натрий-калиевых обменников, выводящих натрий обратно во внеклеточную среду.
В некоторых случаях механизмами вторичного активного транспорта используется энергия перемещения ионов калия в направлении их электрохимического градиента.
Транспортные системы натрий-кальциевого обмена
Существуют по крайней мере два механизма натрий-кальциевого обмена. Наиболее часто встречаются обменники типа NCX (Na-Са exchange), которые были впервые описаны в сердечной мышце, нерве краба и аксоне кальмара. При переносе транспортной молекулой одного иона кальция наружу происходит перенос трех ионов натрия внутрь клетки. Несмотря на то, что сродство NCX к кальцию ниже, чем у кальциевой АТФазы, общая мощность этого транспортного механизма выше приблизительно в 50 раз, поскольку плотность таких молекул в мембране значительно выше. NCX играют важную роль в условиях повышенного входа кальция в клетку, вызванного электрической активностью и превышающего возможности АТФаз по устранению избыточного кальция из клетки.
Для измерения внутриклеточной концентрации кальция использовали флуоресцентную молекулу экворин (aequorin). В состоянии покоя вход кальция в направлении электрохимического градиента уравновешивается за счет переноса ионов из клетки ионным обменником. В начале опыта внутриклеточная концентрация кальция высока потому, что уровень кальция снаружи аксона повышен (112 ммоль).
При снижении внеклеточной концентрации снижается и пассивный вход кальция в аксон. В результате снижается внутриклеточная концентрация кальция, и движущая сила для этого иона возрастает до тех пор, пока скорость пассивного тока вновь не сравняется со скоростью выброса кальция.
С другой стороны, снижение внеклеточной концентрации натрия приводит к увеличению внутриклеточной концентрации кальция, поскольку обменник медленнее выводит кальций в условиях сниженной движущей силы для натрия.
Поэтому происходит повышение внутриклеточного уровня кальция, снижающее скорость входа кальция в клетку. Замена ионов кальция ионами лития, не способными участвовать в работе обменника, приводит к дальнейшему повышению внутриклеточной концентрации кальция.
Реверсия направления работы NCXИзменение градиентов для одного или нескольких ионов, участвующих в работе обменника, может привести к перемене направления его работы. Интересно, что в случае NCX такая смена направления может произойти в физиологических условиях. При этом кальций будет переноситься внутрь клетки, а натрий - выводиться из нее. Направление работы NCX определяется разницей между энергией, выделяемой при перемещении трех ионов натрия внутрь клетки, и энергией, необходимой для переноса одного иона кальция наружу. Одним из факторов, определяющих этот энергетический баланс, является мембранный потенциал. Влияние потенциала мембраны обусловлено тем, что процесс ионообмена не является электрически нейтральным. Вследствие каждого прямого цикла работы обменника, через мембрану внутрь клетки переносится один положительный заряд. Следовательно, гиперполяризация мембраны облегчает прямой цикл обменника, в то время как деполяризация его затрудняет и может привести к изменению направления, то есть к работе обменника в режиме обратного цикла. Следует подчеркнуть, что, несмотря на отсутствие электрической нейтральности, обменники не являются электрогенными. В отличие от насосов, они работают за счет электрохимических градиентов, а не производят их.
Энергия, выделяемая за счет входа ионов натрия в клетку (или энергия, необходимая для переноса этих ионов наружу), равна про изведению заряда на разность между равновесным потенциалом для натрия (ENa) и мембранным потенциалом (Vm). Для трех ионов натрия эта энергия будет равна 3 (. ENa - Vm). Соответственно, для одного двухвалентного иона кальция энергия переноса будет равна 2 (ЕСа-Vm). При определенном значении мембранного потенциала уровни энергии сравняются и перенос прекратится. Обозначив это значение через потенциал реверсии Vr, получим
![]() или
или ![]()
При значениях потенциала более отрицательных, чем потенциал реверсии, натрий перемещается внутрь клетки, а кальций - наружу. При более положительных значениях направление движения ионов сменяется на обратное.
Предположим, что концентрации натрия и кальция в клетке принимают значения 15 ммоль и 100 нМ соответственно, а во внеклеточной среде находится 150 ммоль натрия и 2 ммоль кальция. Такие значения вполне вероятны для клеток млекопитающих. По уравнению Нернста, равновесный потенциал для натрия будет +58 мВ, а для кальция +124 мВ. Потенциал реверсии, при котором движение ионов через обменник прекратится, будет - 74 мВ. Это значение близко к потенциалу покоя многих клеток млекопитающих. Следовательно, в отдельно взятой клетке направление ионообмена может быть как прямым, так и обратным, в зависимости от текущего значения мембранного потенциала и от того, происходило ли накопление ионов натрия или кальция в этой клетке. В клетках сердечной мышцы вход кальция через NCX в ходе потенциала действия способен вызвать сокращение мышцы, а его последующий вывод из клетки посредством того же NCX способствует релаксации.
Натрий-кальциевый обменник в палочках сетчаткиДля клеток с низкими значениями потенциала покоя NCX не представляется надежной системой вывода кальция из цитоплазмы. В таких клетках кальций сначала будет накапливаться до некоторого, достаточно высокого, уровня. Примером такой клетки может служить палочка сетчатки млекопитающих, мембранный потенциал покоя которой обычно находится на уровне - 40 мВ. В мембране таких клеток встречается другой тип натрий-кальциевого обменника, RetX. По сравнению с NCX дополнительная энергия для переноса кальция получается за счет двух отличий в стехиометрии RetX. Во-первых, вместо трех ионов натрия через RetX проходят четыре. Во-вторых, в работе обменника участвует ион калия, также перемещающийся в направлении своего электрохимического градиента и, следовательно, выделяющий при этом дополнительную энергию. Уравнение потенциала реверсии натрий--калий-кальциевого обменника имеет следующий вид:
![]()
Исходя из подсчитанных выше значений ЕNa и ECa и принимая значение ЕK за - 90 мВ, получим из уравнения Нернста Vr. = +74 мВ. Очевидно, что вероятность реверсии ионообмена через RetX весьма мала.
Для лучшего понимания работы RetX представляется целесообразным задать следующий вопрос: какое значение должен принять равновесный потенциал ЕCa для того, чтобы Vr сравнялся с потенциалом покоя клетки, т.е. - 40 мВ. С помощью того же уравнения получим 181 мВ, что при внеклеточной концентрации кальция 2 ммоль дает значение внутриклеточной концентрации 1 нМ. Другими словами, при мембранном потенциале - 40 мВ обменник RetX обладает достаточной энергией для того, чтобы снизить уровень кальция внутри клетки до 1 нМ. Интересно отметить, что при тех же условиях NCX способен понизить внутриклеточную концентрацию кальция только до 383 нМ.
Литература
1. Баранцев Р.Г., Андрианов И.В., Маневич Л.И. Асимптотология - путь к целостной простоте.
2. Пригожин И., Николис Г. Познание сложного. Введение.
3. Пригожин И., Гленсдорф П. Термодинамическая теория структуры, устойчивости и флуктуации
Похожие работы
... Формирование бислоя является особым свойством молекул липидов и реализуется даже вне клетки. Важнейшие свойства бислоя: - способность к самосборке - текучесть - ассиметричность. 1.2. Хотя основные свойства биологических мембран определяются свойствами липидного бислоя, но большинство спецефических функций обеспечивается мембранными белками. Большинство из них пронизывают бислой в виде одиночной ...
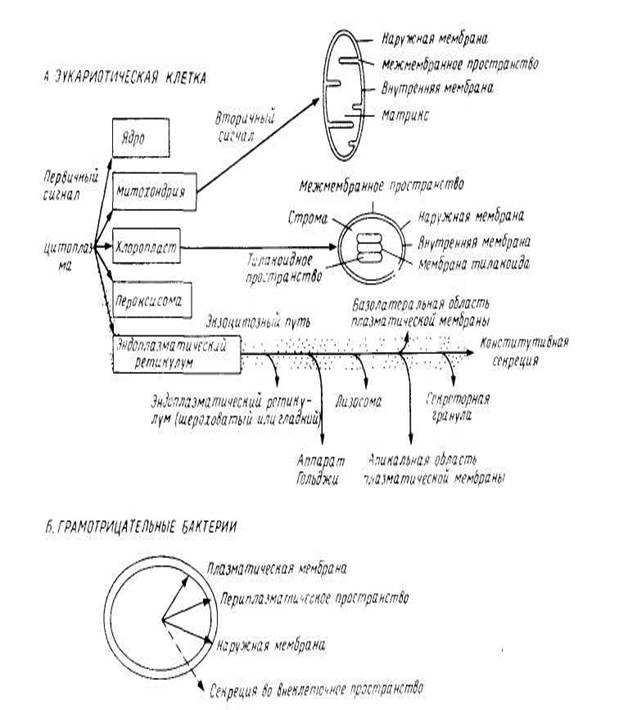
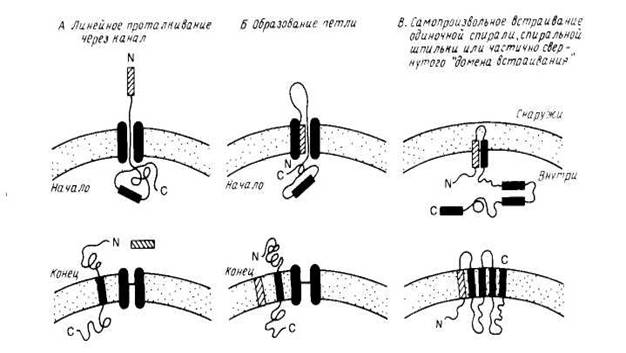
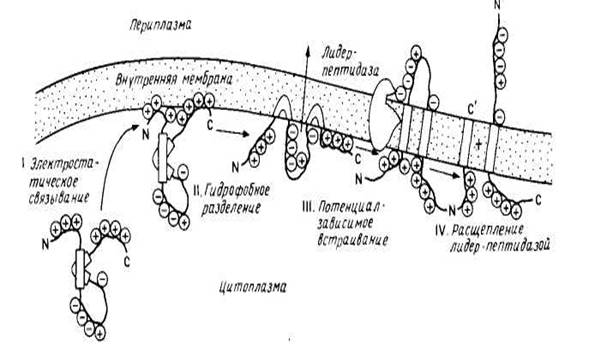
... рассматриваться как вторичные сигналы. В нескольких случаях их удалось идентифицировать как сигнальные последовательности, физически отделенные от первичных, хотя, возможно, так бывает не всегда. рис.1 Сортировка мембранных белков биогенез мембрана белок липид Особый интерес представляет процесс сборки мембранных белков, который целесообразно рассмотреть в связи с их сортировкой. На рис. 2 ...
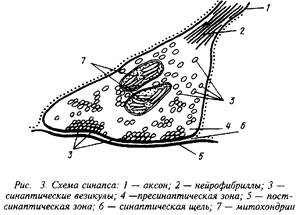
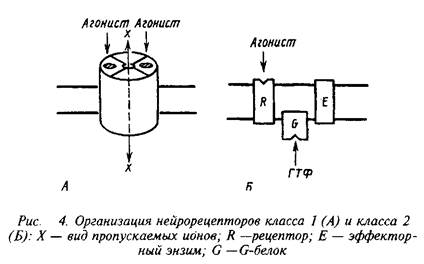
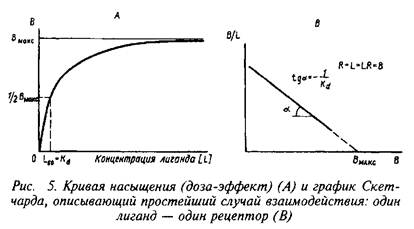
... роль нейрорецепторов сводится к созданию специфических информационных входов, организующих единый функциональный ансамбль нейронов. Именно совокупность рецепторов определяет лицо клетки и ее реакции на поступление разнообразных химических сигналов. Молекулярные механизмы, лежащие в основе модуляции эффективности синаптической передачи, в которых важную роль играют рецепторные процессы, имеют ...
... – это период повышенной возбудимости. Он соответствует следовой деполяризации. В нейронах ЦНС возможна частичная деполяризация вслед за гиперполяризацией. Повышенная возбудимость обусловлена пониженным мембранным потенциалом и повышенной проницаемостью мембраны для Na+. Скорость протекания фазовых изменений возбудимости клетки определяет ее лабильность, или функциональная подвижность. Мерой ...
0 комментариев