В 1786 году профессор анатомии Болонского университета Луиджи Гальвани провел ряд опытов, положивших начало целенаправленным исследованиям в области биоэлектрических явлений. В первом опыте он подвешивал препарат обнаженных лапок лягушки с помощью медного крючка на железной решетке, и обнаружил, что при каждом касании мышцами решетки, они сокращались. Гальвани предположил, что сокращения мышц вообще – следствие воздействия на них «животного электричества», источником которого являются нервы и мышцы. Однако, по мнению Вольта, причиной сокращения был электрический ток, возникший в области контакта разнородных металлов. Гальвани поставил второй опыт, в котором источником тока, действовавшего на мышцу, точно был нерв: мышца опять сокращалась. Таким образом, было получено точное доказательство существования «животного электричества».
Все клетки имеют свой электрический заряд, который формируется в результате неодинаковой проницаемости мембраны для различных ионов. Клетки возбудимых тканей (нервная, мышечная, железистая) отличаются тем, что они под действием раздражителя меняют проницаемость своей мембраны для ионов, в результате чего ионы очень быстро транспортируются согласно электрохимическому градиенту. Это и есть процесс возбуждения. Его основой является потенциал покоя.
Потенциал покоя
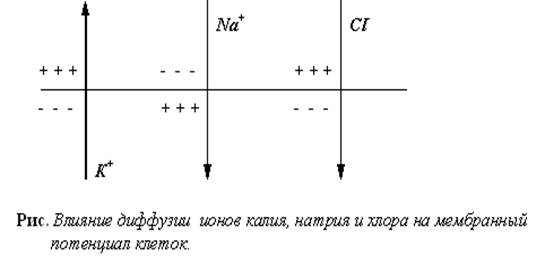
Потенциал покоя – относительно стабильная разность электрических потенциалов между наружной и внутренней сторонами клеточной мембраны. Его величина обычно варьирует в пределах от -30 до -90 мВ. Внутренняя сторона мембраны в покое заряжена отрицательно, а наружная – положительно из-за неодинаковых концентраций катионов и анионов внутри и вне клетки.
Внутри- и внеклеточные концентрации ионов (ммоль/л) в мышечных клетках теплокровных животных
| Внутриклеточная концентрация | Внеклеточная концентрация | |
| Na+ | 12 | 145 |
| K+ | 155 | 4 |
| Ca2+ | 10-8 – 10-7 | 2 |
| Cl- | 4 | 120 – 130 |
| HCO3- | 8 | 27 |
| Высокомолекулярные внутриклеточные анионы | 155 | _ |
| Прочие катионы | Мало | 5 |
В нервных клетках похожая картина. Таким образом, видно, что основную роль в создании отрицательного заряда внутри клетки играют ионы K+ и высокомолекулярные внутриклеточные анионы, главным образом они представлены белковыми молекулами с отрицательно заряженными аминокислотами (глутамат, аспартат) и органическими фосфатами. Эти анионы, как правило, не могут транспортироваться через мембрану, создавая постоянный отрицательный внутриклеточный заряд. Во всех точках клетки отрицательный заряд практически одинаков. Заряд внутри клетки является отрицательным как абсолютно (в цитоплазме анионов больше, чем катионов), так и относительно наружной поверхности клеточной мембраны. Абсолютная разность невелика, однако этого достаточно для создания электрического градиента.
Главным ионом, обеспечивающим формирование потенциала покоя (ПП), является K+. В покоящейся клетке устанавливается динамическое равновесие между числом входящих и выходящих ионовK+. Это равновесие устанавливается тогда, когда электрический градиент уравновесит концентрационный. Согласно концентрационному градиенту, создаваемому ионными насосами, K+ стремится выйти из клетки, однако отрицательный заряд внутри клетки и положительный заряд наружной поверхности клеточной мембраны препятствуют этому (электрический градиент). В случае равновесия на клеточной мембране устанавливается равновесный калиевый потенциал.
Равновесный потенциал для каждого иона можно рассчитать по формуле Нернста:
Eion=RT/ZF·ln([ion]o/[ion]i),
где Eion - потенциал, создаваемый данным ионом;
R – универсальная газовая постоянная;
Т – абсолютная температура (273+37°С);
Z – валентность иона;
F – постоянная Фарадея (9,65·104);
[ion]o – концентрация иона во внешней среде;
[ion]i - концентрация иона внутри клетки.
При температуре 37°С равновесный потенциал для K+ равен -97мВ. Однако реальный ПП меньше – около -90 мВ. Это объясняется тем, что в формирование ПП свой вклад вносят и другие ионы. В целом ПП – это алгебраическая сумма равновесных потенциалов всех ионов, находящихся внутри и вне клетки, включающий также значения поверхностных зарядов самой клеточной мембраны.
Вклад Na+ и Cl- в создание ПП невелик, но, тем не менее, он имеет место. В покое вход Na+ в клетку низкий (намного ниже, чем K+), но он уменьшает мембранный потенциал. Влияние Cl- противоположно, так как это анион. Отрицательный внутриклеточный заряд не позволяет большому количеству Cl- проникнуть в клетку, поэтому Cl- это в основном внеклеточный анион. Как внутри клетки, так и вне ееNa+ и Cl- нейтрализуют друг друга, вследствие чего их совместное поступление в клетку не оказывает существенного влияния на величину ПП.
Наружная и внутренняя стороны мембраны несут на себе собственные электрические заряды, преимущественно с отрицательным знаком. Это полярные составляющие мембранных молекул – гликолипидов, фосфолипидов, гликопротеинов. Ca2+, как внеклеточный катион, взаимодействует с наружными фиксированными отрицательными зарядами, а также с отрицательными карбоксильными группами интерстиция, нейтрализуя их, что приводит к увеличению и стабилизации ПП.
Для создания и поддержания электрохимических градиентов необходима постоянная работа ионных насосов. Ионный насос – это транспортная система, обеспечивающая перенос иона вопреки электрохимическому градиенту, с непосредственными затратами энергии. Градиенты Na+ и K+ поддерживаются с помощью Na/K – насоса. Сопряженность транспорта Na+ и K+ примерно в 2 раза уменьшает энергозатраты. В целом же траты энергии на активный транспорт огромны: лишь Na/K – насос потребляет около 1/3 всей энергии, расходуемой организмом в покое. 1АТФ обеспечивает один цикл работы – перенос 3Na+ из клетки, и 2 K+ в клетку. Асимметричный перенос ионов способствует заодно формированию и электрического градиента (примерно 5 – 10мВ).
Нормальная величина ПП является необходимым условием возникновения возбуждения клетки, т.е. распространения потенциала действия, инициирующего специфическую деятельность клетки.
Потенциал действия (ПД)
ПД – это электрофизиологический процесс, выражающийся в быстром колебании мембранного потенциала, вследствие специфического перемещения ионов и способный распространяться без декремента на большие расстояния. Амплитуда ПД колеблется в пределах 80 – 130 мВ, длительность пика ПД в нервном волокне – 0,5 – 1 мс. Амплитуда потенциала действия не зависит от силы раздражителя. ПД либо совсем не возникает, если раздражение подпороговое, либо достигает максимальной величины, если раздражение пороговое или сверхпороговое. Главным в возникновении ПД является быстрый транспорт Na+ внутрь клетки, что способствует вначале снижению мембранного потенциала, а затем – изменению отрицательного заряда внутри клетки на положительный.
В составе ПД различают 3 фазы: деполяризацию, инверсию, и реполяризацию.
1. Фаза деполяризации. При действии на клетку деполяризующего раздражителя начальная частичная деполяризация происходит без изменения ее проницаемости для ионов (не происходит движение Na+ внутрь клетки, т. к. закрыты быстрые потенциалчувствительные каналы для Na+). Na+ - каналы обладают регулируемым воротным механизмом, который расположен на внутренней и внешней сторонах мембраны. Имеются активационные ворота (m – ворота) и инактивационные (h – ворота). В покое m – ворота закрыты, а h – ворота открыты. В мембране также имеются K+ - каналы, имеющие только одни ворота (активационные), закрытые в покое.
Когда деполяризация клетки достигает критической величины (Екр – критический уровень деполяризации, КУД), которая обычно равна 50мВ, проницаемость для Na+ резко возрастает – открывается большое количество потенциалзависимых m – ворот Na+ - каналов. За 1 мс через 1 открытый Na+ - канал в клетку попадает до 6000 ионов. Развивающаяся деполяризация мембраны вызывает дополнительное увеличение ее проницаемости для Na+, открываются все новые и новые m - ворота Na+ - каналы, так что ток Na+ имеет характер регенеративного процесса (сам себя усиливает). Как только ПП становится равным нулю, фаза деполяризации заканчивается.
2.Фаза инверсии. Вход Na+ в клетку продолжается, т. к. m - ворота Na+ - каналы еще открыты, поэтому внутри клетки заряд становится положительным, а снаружи – отрицательным. Теперь электрический градиент препятствует входу Na+ в клетку, однако, из-за того, что концентрационный градиент сильнее электрического, Na+ все же проходит в клетку. В тот момент, когда ПД достигает максимального значения, происходит закрытие h – ворот Na+ - каналов (эти ворота чувствительны к величине положительного заряда в клетке) и поступление Na+ в клетку прекращается. Одновременно открываются ворота K+ - каналов. K+ транспортируется из клетки согласно химическому градиенту (на нисходящей фазе инверсии – еще и по электрическому градиенту). Выход положительных зарядов из клетки приводит к уменьшению ее заряда. K+ с небольшой скоростью может выходить из клетки также через неуправляемые K+ - каналы, которые всегда открыты. Все рассмотренные процессы являются регенеративными. Амплитуда ПД складывается из величины ПП и величины фазы инверсии. Фаза инверсии заканчивается, когда электрический потенциал снова становится равным нулю.
3.Фаза реполяризации. Связана с тем, что проницаемость мембраны для K+ еще высока, и он выходит из клетки по градиенту концентрации, несмотря на противодействие электрического градиента (клетка внутри снова имеет отрицательный заряд). Выходом K+ обусловлена вся нисходящая часть пика ПД. Нередко в конце ПД наблюдается замедление реполяризации, кто связано с закрытием значительной части ворот K+ - каналов, а также – с возрастанием противоположно направленного электрического градиента.
Если из внеклеточной среды убрать Na+ или заблокировать Na+ - каналы, то ПД не возникает. Так с помощью местных анестетиков расстраивается механизм управления у ворот Na+ - каналов.
Следовые явления в процессе возбуждения клетки
В конце ПД нередко наблюдается замедление реполяризации, что называют отрицательным следовым потенциалом. Затем может быть зарегистрирована гиперполяризация мембраны (характерно для нервных клеток) – положительный следовый потенциал. Вслед за ним может возникнуть частичная деполяризация клеточной мембраны – также отрицательный следовый потенциал.
Следовая гиперполяризация обычно является результатом еще сохраняющейся повышенной проницаемости для К+. Характерна для нейронов. Активационные ворота K+ - каналов еще не полностью закрыты, K+ продолжает выходить из клетки согласно концентрационному градиенту. Na/K – насос непосредственно за фазы ПД не отвечает, хотя он работает непрерывно в покое и продолжает работать во время развития ПД. Возможно, Na/K – насос способствует развитию следовой гиперполяризации. Длительная гиперполяризация хорошо выражена в тонких немиелинизированных нервных волокнах (болевых афферентах).
Следовая деполяризация также характерна для нейронов. Возможно, она связана с кратковременным повышением проницаемости мембраны для Na+ и входом его в клетку по градиентам.
Исследования ионных токов
Наиболее распространенный метод исследования ионных каналов – это метод фиксации напряжения (voltage - clamp). Мембранный потенциал с помощью подачи электрического напряжения изменяют и фиксируют на определенном уровне. Затем мембрану градуально деполяризуют, что ведет к открытию ионных каналов и возникновению ионных токов, которые могли бы деполяризовать клетку. Однако при этом пропускается электрический ток, равный по величине, но противоположный по знаку, поэтому трансмембранная разность потенциалов не изменяется. Это дает возможность получить величину ионного тока через мембрану.
Количественное соотношение между ионными токами по отдельным каналам в покое и во время ПД можно выяснить с помощью метода локальной фиксации потенциала (patch clamp). К мембране подводят микроэлектрод – присоску (внутри него создается разрежение) и, если на этом участке оказывается канал, исследуют ионный ток через него. В остальном методика подобна предыдущей. Таким методом было установлено, что через Na+ - каналы может проходить и К+, но его ток в 10 – 12 раз меньше. Na+ также может проходить через К+ - каналы в 100 раз менее интенсивно.
Резерв в клетке ионов, обеспечивающих возникновение возбуждения, огромен. Концентрационные градиенты ионов в результате одного цикла возбуждения практически не изменяются. Клетка может возбуждаться до 5·105 раз без подзарядки, т. е. без работы Na/K – насоса. Число импульсов, которое генерирует и проводит нервное волокно, зависит от его толщины, определяющей запас ионов.
Если сила раздражителя, действующего на нервную ткань мала, деполяризация не достигает критического уровня, импульс не возникает. В этом случае ответ ткани на раздражение будет носить форму локального потенциала. Величина такого потенциала вариабельна, она может достигать 10 – 40 мВ. Локальными являются также возбуждающий и тормозной постсинаптические потенциалы, рецепторный и генераторный потенциалы.
Сравнительная характеристика локального потенциала и ПД
| Свойство | Локальный потенциал | Потенциал действия |
| Распространение | На 1 – 2 мм с затуханием (декрементом) | Без декремента на большие расстояния по всей длине нервного волокна |
| Зависимость от величины стимула | Возрастает с увеличением силы раздражителя, т. е. подчиняется закону «силы» | Не зависит (подчиняется закону «все или ничего») |
| Явление суммации | Суммируется – возрастает при частых повторных подпороговых раздражениях | Не суммируется |
| Амплитуда | 10 – 40 мВ | 80 – 130 мВ |
| Возбудимость ткани при возникновении потенцала | Увеличивается | Уменьшается вплоть до абсолютной рефрактерности |
Повышение возбудимости клетки во время локального потенциала объясняется тем, что мембрана оказывается частично деполяризованной. Если КУД остается на постоянном уровне, то для его достижения требуется гораздо меньший раздражитель. Амплитуда ПД не зависит от силы раздражителя, потому что он возникает вследствие регенеративных процессов.
Возбудимость клетки во время ПД быстро и сильно изменяется. Различают несколько фаз изменения возбудимости:
1. Кратковременное повышение возбудимости в начале ПД. В зависимости от силы раздражителя может формироваться либо локальный потенциал, либо ПД. Возбудимость повышается потому, что клетка частично деполяризована и ПП приближается к критическому значению. Когда деполяризация достигает 50% от пороговой величины, начинают открываться быстрые потенциал – чувствительные Na+ - каналы.
2. Абсолютная рефрактерность – это полная невозбудимость клетки. Соответствует пику ПД и продолжается 1 – 2 мс. Невозбудимость на фазе деполяризации и восходящей стадии инверсии обусловлена тем, что запущен каскад регенеративных реакций, на который повлиять извне уже нельзя: m - ворота Na+ - каналов уже открыты, а еще закрытые открываются в ответ на уменьшение мембранного потенциала. В период нисходящей стадии инверсии мембрана невозбудима, т. к. закрываются инактивационные ворота, состояние которых не может изменить даже сильное раздражение. Абсолютная рефрактерность продолжается и в период реполяризации до достижения величины Екр ± 10 мВ.
Абсолютная рефрактерная фаза ограничивает максимальную частоту генерации ПД. Если абсолютная рефрактерность завершается через 2 мс после начала ПД, клетка может возбуждаться с частотой максимум 500 имп/с. Нейроны ретикулярной формации и толстые миелиновые нервные волокна могут генерировать ПД с частотой 1000 имп/с.
3. Относительная рефрактерная фаза – период восстановления возбудимости, когда сильное раздражение может вызвать новое возбуждение. Соответствует конечной стадии реполяризации и следовой гиперполяризации. Пониженная возбудимость связана с повышенным транспортом К+ из клетки. Поэтому для вызова возбуждения необходимо более сильное раздражение. Кроме того во время гиперполяризации потенциал больше и, следовательно, дальше отстоит от КУД. У нервных волокон относительная рефрактерность длится несколько мс.
4. Фаза экзальтации – это период повышенной возбудимости. Он соответствует следовой деполяризации. В нейронах ЦНС возможна частичная деполяризация вслед за гиперполяризацией. Повышенная возбудимость обусловлена пониженным мембранным потенциалом и повышенной проницаемостью мембраны для Na+.
Скорость протекания фазовых изменений возбудимости клетки определяет ее лабильность, или функциональная подвижность. Мерой лабильности является максимальное число ПД, которое может ткань воспроизвести в 1 с. Лабильность нерва равна 500 – 1000, нервно – мышечного синапса около 100 имп/с. При постепенном увеличении частоты ритмического раздражителя лабильность ткани повышается.
Показателями состояния возбудимости ткани являются пороговый потенциал, пороговая сила, пороговое время.
Пороговый потенциал (∆V) – это минимальная величина, на которую надо уменьшить ПП, чтобы вызвать возбуждение:
∆V = Е0 - Екр,
где Е0 – это потенциал покоя.
Пороговая сила – это наименьшая сила раздражителя, способная вызвать ПД при неограниченном во времени действии раздражителя. При использовании в качестве раздражителя электрический ток, его пороговая сила равна 1 реобазе. Если возбудимость ткани высока, пороговая сила раздражителя мала.
Аккомодация – это понижение возбудимости ткани и амплитуды ПД вплоть до полного их исчезновения при медленно нарастающем стимуле. Ее главной причиной является инактивация Na+ - каналов, возникающая при медленной деполяризации мембраны – в течение 1 с и более. Клетка теряет возбудимость, если закрывается около 50% инактивационных ворот Na+ - каналов.
Пороговое время – это минимальное время, в течение которого должен действовать на ткань раздражитель пороговой силы, чтобы вызвать ее возбуждение. Хронаксия – наименьшее время, в течение которого должен действовать ток в две реобазы, чтобы вызвать возбуждение.
Проведение нервных импульсов по нервным волокнам
Нервные волокна представляют собой отростки нейронов, с помощью которых осуществляется связь между нейронами, а также с эффекторами. В состав нервного волокна входит осевой цилиндр (нервный отросток) и глиальная оболочка. По взаимоотношению с глиальными клетками выделяют миелиновые и безмиелиновые нервные волокна. Оболочку безмиелиновых волокон образуют шванновские клетки (леммоциты). При этом осевые цилиндры прогибают клеточную оболочку леммоцитов и погружаются в них. Место, где имеется сдвоенная мембрана леммоцита, называется мезаксон. У миелиновых волокон мезаксон удлиняется и спирально закручивается вокруг осевого цилиндра, формируя электроизолирующую миелиновую оболочку. Миелиновая оболочка через равные участки (0,5 – 2 мм) прерывается, образуя, свободные от миелина участки, узловые перехваты Ранвье. Их протяженность находится в пределах 0,25 – 1,0 мкм, в волокнах ЦНС – до 14 мкм. В перехватах возможно формирование ПД, т. к. там есть потенциалзависимые Na+ - каналы. В межузловых сегментах таких каналов нет. В безмиелиновых нервных волокнах Na+ - каналы расположены равномерно по всей поверхности.
Распространение локальных потенциалов
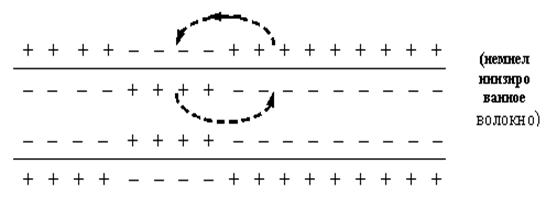
Локальный потенциал изменяет ПП в сторону деполяризации в результате входа в клетку Na+ согласно электрохимическому градиенту. В результате между деполяризованными и соседними участками волокна формируется градиент, вызывающий передвижение ионов Na+ в соседние участки волокна, а ионы на наружной поверхности волокна движутся в противоположном направлении. В итоге поляризация соседнего участка уменьшается.
Затухание локального потенциала связано с отсутствием потенциалзависимых Na+ - каналов или их неактивацией, с продольным сопротивлением цитоплазмы волокна и шунтированием тока во внеклеточную среду через каналы утечки. Деполяризация мембраны, не сопровождающаяся изменением проницаемости потенциалзависимых Na+ - и К+ - каналов, называется электротонической. Она характерна для участков, где такие каналы отсутствуют: большая часть мембраны дендритов, межперехватные промежутки миелиновых волокон. Если электротоническая деполяризация достигает участков с потенциалзависимыми каналами, но его амплитуда не достигает порогового значения, формируется препотенциал, а если достигает – ПД.
Эффективность электротонического распространения зависит от сопротивления и емкости мембраны, сопротивления цитоплазмы (улучшается при увеличении диаметра волокна, т. е. с уменьшением сопротивления цитоплазмы, а также при миелинизации волокна, т. е. с увеличением сопротивления мембраны и уменьшением ее емкости). Эффективность электротонического распространения характеризуется постоянной длиной мембраны (λm). Это расстояние, на которое может электротонически распространиться биопотенциал, пока его амплитуда не уменьшится до 37% от исходной величины.
Проведение потенциала действия
В распространении ПД можно выделить два этапа:
- этап электротонического проведения – физический механизм;
- генерация ПД в новом участке на пути его движения, обусловленная реакцией ионных каналов.
В зависимости от расположения и концентрации ионных каналов в мембране возможно два типа проведения ПД: непрерывный и сальтаторный (скачкообразный).
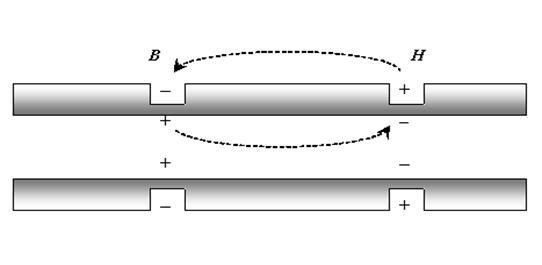
1. Непрерывное распространение ПД осуществляется в безмиелиновых волокнах, имеющих равномерное распределение по поверхности ионных каналов. На расстояние постоянной длины мембраны потенциал распространяется электротонически, а далее формируется новый ПД. Число электротонического распространения невелико из-за того, что равномерно распределенные по поверхности каналы, находятся в непосредственной близости, и все они обязательно возбуждаются при уменьшении мембранного потенциала. Таким образом, непрерывное распространение возбуждения идет через генерацию нервных импульсов по эстафете, когда каждый участок мембраны выступает сначала как раздражаемый, а затем как раздражающий.
2. Сальтаторный тип проведения ПД осуществляется в миелиновых волокнах, в которых потенциалзависимые ионные каналы сконцентрированы только в перехватах Ранвье, где их плотность составляет 12000 на 1 мкм2, что в 100 раз выше, чем в мембранах безмиелиновых волокон. В межузловых сегментах возбудимых каналов почти нет, и мембрана практически невозбудима. В этих участках ПД распространяется только электротонически, а по достижении следующего перехвата снова генерируется ПД. λm миелинового волокна равна 5 мм, поэтому в случае повреждения соседних перехватов ПД может электротонически возбудить 2-й – 4-й, и даже 5-й перехваты. Т. о., сальтаторное проведение возбуждения имеет два важных преимущества: высокая скорость проведения (электротонический транспорт в 107 быстрее непрерывного проведения возбуждения) и энергетически экономично, т. к. снижения концентрационных градиентов после проведения возбуждения меньше, чем в безмиелиновых нервных волокнах.
Синаптическая передача возбуждения
Синапс - специализированный сигнальный межклеточный контакт, обеспечивающий передачу возбуждающих или тормозных влияний между двумя возбудимыми клетками. Через синапс также осуществляется трофическое влияние, приводящее к изменению метаболизма иннервируемой клетки. Синапс включает в себя пресинаптическую и постсинаптическую мембраны, а также – синаптическую щель.
Пресинаптическое окончание – расширенное окончание аксона, в котором имеются синаптические пузырьки диаметром 40 нм, содержащие медиатор. В неактивном состоянии везикулы посредством белка синапсина связаны с цитоскелетом. Также в пресинаптическом окончании есть митохондрии, осуществляющие энергообеспечение синаптической передачи, цистерны гладкой ЭПС, в которых депонируется Ca2+, а также микротрубочки и микрофиламенты, участвующие во внутриклеточном перемещении везикул. Часть мембраны, ограничивающая синаптическую щель, называется пресинаптической мембраной. Через нее происходит экзоцитоз медиатора в синаптическую щель.
Синаптическая щель содержит межклеточную жидкость и мукополисахаридное плотное вещество в виде полосок, мостиков, которое обеспечивает связь между пре- и постсинаптическими мембранами и может содержать ферменты, участвующие в метаболизме медиатора.
Постсинаптическая мембрана – утощенная часть клеточной мембраны иннервируемой клетки, содержащая белковые рецепторы, способные связывать молекулы медиатора.
Механизм синаптической передачи
Передача осуществляется в два главных этапа.
1. Преобразование электрического сигнала в химический (электросекреторное сопряжение). ПД, поступивший в пресинаптическое окончание, вызывает его деполяризацию, открывающую потенциалзависимые Ca2+- каналы. Ионы Ca2+ входят в клетку согласно электрохимическому градиенту, что активирует фосфорилирование синапсина и последующее ослабление связи везикул с цитоскелетом. Везикула перемещается к мембране. При контакте с мембраной происходитферментативное «плавление»ее стенки, а также активация белка синаптопорина, формирующего канал, через который происходит выход медиатора в синаптическую щель путем экзоцитоза. Выделение молекул медиатора пропорционально количеству поступившего туда Ca2+ в 4-й степени, т. е. имеется усиление сигнала. Выделение медиатора происходит квантами, каждый из которых содержит до 10 тыс. молекул. После поступления в синаптическую щель молекулы медиатора диффундируют к постсинаптической мембране и вступают во взаимодействие с ее рецепторами. Скорость диффузии молекул медиатора позволяет им пройти расстояние синаптической щели в течение 0,1 – 0,2 мс. Длительность действия медиатора на рецепторы равна около 1 мс, что гораздо меньше его периода полураспада. Это значит, что медиатор удаляется из синаптической щели. Удаление происходит путем диффузии в окружающее межклеточное вещество и разрушения эстеразой.
2. Преобразование химического сигнала обратно в электрический. Действие молекул медиатора на рецепторы приводит к открытию ионных каналов. Открытое состояние сохраняется 1мс, в течение которого через него проходит около 500000 ионов. Ток Na+ через канал превосходит ток К+, т. к. транспорту К+ противостоит электрический градиент. Формируется деполяризация, называемая возбуждающим постсинаптическим потенциалом (ВПСП). Высокая возбудимость в синапсах может поддерживаться путем спонтанного выделения из пресинаптической мембраны 1 - 2 квантов медиатора во время между импульсами. Кроме того существует неквантовая утечка медиатора, которая, по предположениям, оказывает трофическое влияние.
В нейронах ЦНС возникновение ВПСП связано также с транспортом Ca2+ . Кроме быстрых Na+ - потенциалов существуют медленные кальциевые. В телах некоторых нейронов ПД создается преимущественно за счет Ca2+, а в аксоне – главным образом за счет Na+.
Таким образом видно, какое важное значение в организме играет неравномерное распределение ионов. Потенциальная энергия химических и электрических градиентов велика и используется организмом далеко не только для информационной связи между отдельными частями организма и внешней средой. Эта энергия может переводиться в энергию химических связей, как например в процессах фотосинтеза и внутриклеточного дыхания, может использоваться для транспорта через мембрану других веществ (как, например, при всасывании питательныъх веществ в кишечнике, реабсорбции веществ в канальцах нефрона), регуляции параметров внутренней среды и многих других процессах.
Похожие работы
... вдоль волокна без затухания (бездекрементное проведение). Это обусловлено тем, что локальные токи только деполяризуют мембрану до критического уровня, а потенциалы действия в каждом участке мембраны поддерживаются независимыми ионными потоками, перпендикулярными к направлению распространения возбуждения. Скорость уменьшения мембранного потенциала до критического уровня зависит от разности ...
... Формирование бислоя является особым свойством молекул липидов и реализуется даже вне клетки. Важнейшие свойства бислоя: - способность к самосборке - текучесть - ассиметричность. 1.2. Хотя основные свойства биологических мембран определяются свойствами липидного бислоя, но большинство спецефических функций обеспечивается мембранными белками. Большинство из них пронизывают бислой в виде одиночной ...
... эффективность этой транспортной системы не слишком высока. Тем не менее, с задачей устранения входящего в клетку кальция она справляется. Натрий-кальциевый обменник Во многих механизмах ионного транспорта используется совершенно иной принцип переноса ионов через мембрану против электрохимического градиента. Вместо энергии расщепленной молекулы АТФ эти механизмы используют энергию уже ...
... . И.П. Павлову, первому из русских ученых, 7 октября 1904 г. была присуждена Нобелевская премия в знак признания его работ по физиологии пищеварения. Потребность организма в пище проявляется в виде физиологической реакции голода. У человека голод приобретает выраженную субъективную окраску — от относительного безразличия к пище до яркой эмоциональной реакции. Физиологической основой голода ...
0 комментариев