Навигация
2. ХРОМОСОМЫ ЭУКАРИОТ.
В 80-х годах прошлого столетия в ядрах эукариотических клеток были открыты нитевидные структуры (В. Флемминг, Э. Страсбургер, Э. Ван Бенеден), названные В. Вальдейером (1888 г.) хромосомами (от греч. chroma – цвет, окраска, soma – тело). Этим термином было подчеркнуто сильное сходство хромосом по сравнению с другими клеточными органеллами к основным красителям. В течение последующих 10 – 15 лет большинством биологов было подтверждено, что именно хромосомы служат материальным носителем наследственности.
Таким образом, митотический цикл включает митоз и промежуток между митозами – интерфазу. Интерфаза состоит из трех периодов: центрального – фазы синтеза ДНК (S), когда генетический материал удваивается, а также предсинтетического (G1) и постсинтетического (G2), после которого клетка вступает в митоз (М). После фазы синтеза ДНК в G2-периоде и в митозе, вплоть до анафазы, в хромосоме обнаруживаются две нити, называемые сестринскими хроматидами.
Основной химический компонент хромосом – молекулы ДНК. Содержание ее в ядрах соматических клеток в два раза больше, чем в ядрах зрелых половых клеток. Эти два типа клеток отличаются друг от друга и по числу хромосом. Число хромосом – п в соматических клетках и количество ДНК – с (от англ. content – содержание) в них обозначают как диплоидное (2п хромосом, 2с ДНК), а в зрелых половых клетках как гаплоидное (п хромосом, с ДНК). После фазы синтеза ДНК в соматических клетках число хромосом не изменяется (2п), однако каждая из них содержит две сестринские хроматиды, т.е. идентичные молекулы ДНК, поэтому содержание ДНК в ядрах G2-фазы 4
2.1. Митоз.
Митоз, или непрямое деление, - основной способ размножения эукариотических клеток, обусловливающий, в частности, возможность увеличения их биомассы, рост и регенерацию. Митоз состоит из четырех фаз:
Первая – профаза – характеризуется началом цикла компактизации хромосом, который продолжается в течение всей этой фазы. К концу профазы исчезают ядрышко и ядерная мембрана.
Вторая –метафаза. Хромосомы выстраиваются по экватору клетки. Хроматиды соединены между собой между собой в центромере, называемой также первичной перетяжкой.
Третья – анафаза – начинается с разрыва ценромеры, в результате чего сестринские хроматиды расходятся к разным полюсам клетки. С этого момента каждая пара сестринских хроматид получает название дочерних хромосом.
Четвертая – телофаза. Хромосомы достигают полюсов клетки, появляются ядерная мембрана, ядрышко. Заканчивается митоз делением цитоплазмы и в типичных случаях – восстановлением исходной биомассы дочерних клеток.
Биологическая роль митоза состоит в обеспечении идентичной генетической информацией двух дочерних клеток. Это достижимо только благодаря циклу компактизации – декомпактизации, который и позволяет распределить наследственные молекулы в минимальном объеме митотических хромосом. В противном случае, учитывая размеры клетки (десятки или сотни кубических микрометров) и длину декомпактизованной хромосомы (сантиметры), каждое клеточное деление сопровождалось бы хаотичным переплетением хромосомного материала.
В эволюции эукариотических клеток, видимо, это обстоятельство и послужило причиной становления столь сложного генетического процесса, как митоз.
2.2 Мейоз.
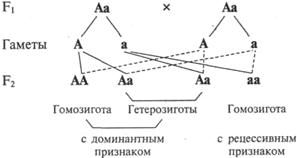
Термином «мейоз» обозначают два следующих друг за другом деления, в результате которых из диплоидных клеток образуются гаплоидные половые клетки – гаметы (рис. 4) Если бы оплодотворение происходило диплоидными гаметами, то плоидность потомков в каждом следующем поколении должна была бы возрастать в геометрической прогрессии. В то же время благодаря мейозу зрелые гаметы всегда гаплоидны, что позволяет сохранять диплоидность соматических клеток вида. Возможность существования подобного мейозу деления при созревании гамет животных и растений была предсказана А. Вейсманом еще в 1887 г. Мейотические деления не эквивалентны митозу. Обоим мейотическим делениям предшествует только одна фаза синтеза ДНК. Продолжительность ее, как и профазы I деления мейоза, во много раз превосходит соответствующие показатели митотического цикла любых соматических клеток данного вида. Главные события мейоза развертываются в профазе I деления. Она состоит из пяти стадий.
В первой стадии – лептотене, следующей непосредственно за окончанием предмейотического синтеза ДНК, выявляются тонкие длинные хромосомы.
Во второй стадии профазы I деления – зиготене – происходит тесное сближение по всей длине (конъюгация) гомологичных хромосом. Гомологичными называются хромосомы, имеющие одинаковую форму и размер, но одна из них получена от матери, другая – от отца. Гаплоидный набор равен числу пар гомологов.
Третья стадия профазы I деления – пахитена – у большинства видов самая длительная. В результате в каждом гомологе смешиваются отцовский и материнский наследственный материал. По мере приближения к метафазе первого деления число хиазм уменьшается.
В метафазе I деления мейоза район центромеры каждой хромосомы соединен (в отличие от метафазы митоза) нитью веретена только с одним полюсом клетки, причем центромеры разошедшихся гомологов всегда связаны с противоположными полюсами. Анафазе I деления мейоза не предшествует расщепление центромеры, как при митозе, и поэтому к полюсам отходят не хроматиды, а целые хромосомы, состоящие из двух хроматид.
Второе деление мейоза, следующее после краткого промежутка – интеркинеза, приводит в соответствие число хромосом и содержание ДНК. Формально оно напоминает митоз. В начале анафазы происходит разделение центромеры, сестринские хроматиды становятся дочерними хромосомами и расходятся к полюсам.
Итак, главное отличие мейоза от митоза – конъюгация гомологичных хромосом с последующим расхождением их в разные гаметы. Точность расхождения обусловлена точностью конъюгации, а последняя – идентичностью молекулярной структуры ДНК гомологов.
В заключение отметим, что цитологами доказано независимое расхождение негомологичных хромосом в профазе I деления мейоза. Это означает, что любая отцовская хромосома может попасть в гамету с любой, в крайнем варианте – со всеми материнскими негомологичными хромосомами. Однако если речь идет о дочерних хромосомах (во II делении мейоза), образовавшихся из перекрещенных, т.е. претерпевших кроссинговер, или кроссоверных хроматид (рис.4), то их, строго говоря, нельзя рассматривать ни как чисто отцовские, ни как чисто материнские.
Похожие работы
... за данную аномалию, локализованы в половых хромосомах. Поскольку особи разного пола несут разные половые хромосомы (XX у женщин и XY у мужчин), это приводит к определенным отклонениям от менделевских закономерностей наследования. Цветовая слепота вызывается рецессивным аллелем с на X-хромосоме. В результате того, что мужчины получают свою единственную X-хромосому от матери, даже одного аллеля, ...
... индивидуальных хромосом; - накапливать сведения об изменениях хромосом в опухолевых клетках, у больных с наследственными заболеваниями крови и т.д. Главный недостаток методов, основанных на использовании низших организмов, заключается в невозможности экстраполировать полученные результаты на человека в связи с отсутствием процессов метаболической активации и детоксикации, характерных для всех ...
... ─ и конкордантные, и дискордантные (как и в семье из двух детей). Наследственные болезни подразделяются на хромосомные и генные болезни (наследственные заболевания обмена веществ, наследственные нарушения иммунитета, болезни с преимущественным поражением эндокринной системы и др.) Хромосомные болезни. Характеризуются изменением структуры или числа хромосом. Они встречаются примерно у 1-2 ...
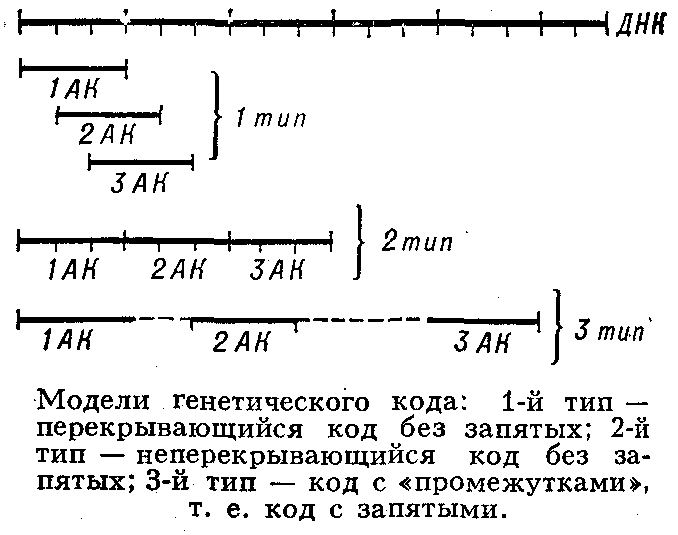
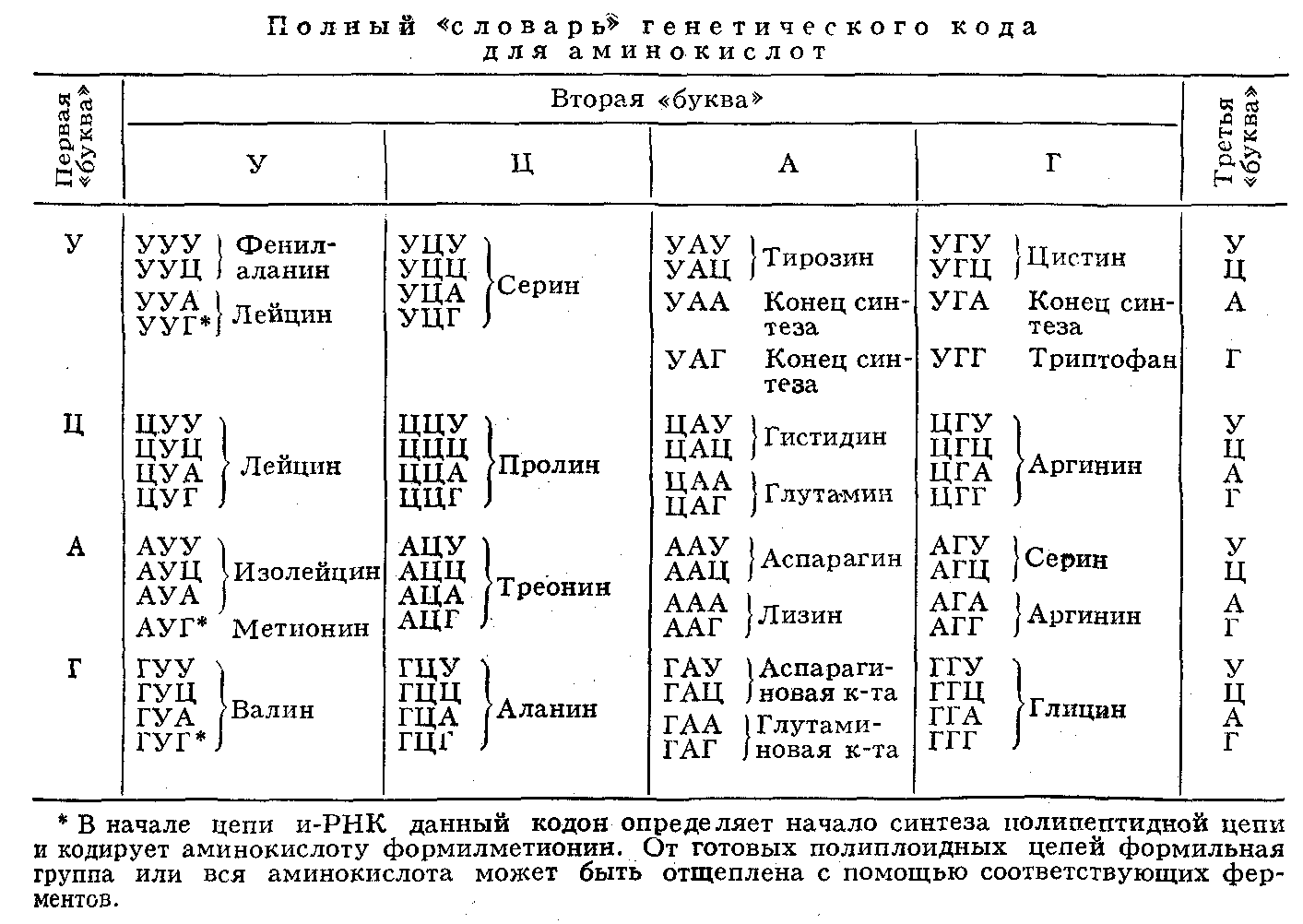
... живые организмы-и удивительное многообразие генов, кодирующих эти белки. В геноме каждого человека есть какие-то области, определяющие его индивидуальность. Некоторые гены человека отличаются от генов крысы всего несколько нуклеотидами-знаками генетического кода. Другие гены у них разные, но одинаковые у двух людей. Изменчивость, связанная с существованием генов , подобных генам группы крови у ...
0 комментариев