МИНИСТЕРСТВО ОБРАЗОВАНИЯ РОССИЙСКОЙ ФЕДЕРАЦИИ
ФЕДЕРАЛЬНОЕ АГЕНТСТВО ПО ОБРАЗОВАНИЮ
БАЛАШОВСКИЙ ФИЛИАЛ
САРАТОВСКОГО ГОСУДАРСТВЕННОГО УНИВЕРСИТЕТА
ИМ. Н.Г. ЧЕРНЫШЕВСКОГО
КАФЕДРА БИОЛОГИИ И ЭКОЛОГИИ
КОНТРОЛЬНАЯ РАБОТА ПО ГЕНЕТИКЕ
Выполнила: студентка IV курса заочного отделения ФЭБ
Демидова Светлана Григорьевна
Проверила: Сулига Е.М.
БАЛАШОВ 2007
Содержание
1. Хромосомы, их строение, видовая специфичность, кариотип
2. Мейоз как цитологическая основа образования и развития половых клеток
3. Сцепленное с полом наследование
4. Транскрипция ДНК
1. Хромосомы, их строение, видовая специфичность, кариотип
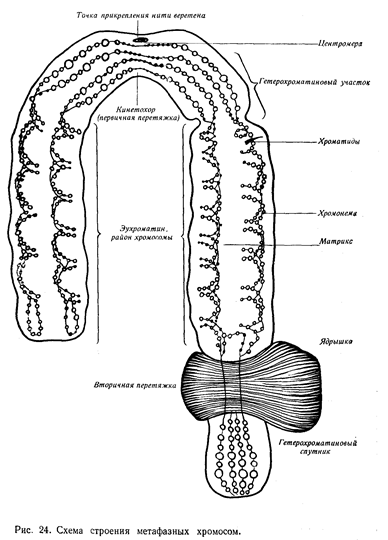
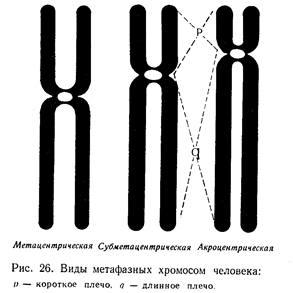
Хромосомы - наиболее важные компоненты ядра. Они играют ведущую роль в явлениях наследственности. Хромосомы хорошо видны под микроскопом в момент деления клетки. Хромосомы ядра неделящейся клетки не видны, поскольку они деконденсации хромосом, тем активнее протекают метаболические процессы в самом ядре. Морфологические хромосомы растений чаще всего имеют нитевидную или палочкообразную форму. Большинство хромосом разделено первичной перетяжкой на два плеча. Под микроскопом первичная перетяжка представлена светлой (неокрашенной) зоной, получившее название центромеры, которые играют основную роль в перемещении хромосом строго определении ядра. Центромера занимает на каждой из хромосом строго определенного место. По положению центромеры хромосомы делят на метацентрические (приблизительно равноплечие), субметацентрические (неравноплечие) и акроцентрические (головчатые), у которых центромера сдвинута к одному из концов. У некоторых хромосом имеется и вторичная перетяжка. Она, как правило, располагается у дистального конца хромосомы и отделяет небольшой ее участок, носящий название спутника. Вторичная перетяжка не участвует в движении хромосом при деление ядра. Она получила название ядрышкового организатора, поскольку в месте ее локализация происходит образование ядрышка. Концевые участки хромосомы называют теломерными. Они препятствуют ее соединению с другими хромосомами.
![]()
1 2 3 4 5
Формы хромосом на стадии метафазы (схема):
1,5 - равноплечие; 2,3 - неравноплечие; 4 – головчатые
Каждому из населяющих нашу планету видов растений и животных свойственно строгое число хромосом, обозначаемое 2n (диплоидный набор). В половых клетках число хромосом в два раза меньше и равно n (гаплоидный набор). В соматических клетках организма каждая хромосома имеет пара, идентичную как морфологически, так и генетически (гомологичные хромосомы). Исключение из этого правила составляют половые хромосомы у гетерогаметных особей. Специфический для определенного вида по числу и структуре набор хромосом получил название кариотипа.
Графическое изображение кариотипа, показывающие его структурные особенности, называется идиограммой. В последние годы получил распространение метод дифференциального окрашивание хромосом. При этом на каждого из хромосом прокрашиваются специфические, характерные для нее полосы (бэнды), что значительно облегчает идентификацию отдельных хромосом кариотипа. Хромосомы, определяющие пол особи, называют половыми хромосомами, а все остальные - аутосомами. Внутренне строение хромосом чрезвычайно сложно. По химическому составу они на 40% состоят из ДНК и на 60% из белков, в среднем около 60% из которых приходится гистоны. Строение метафазной хромосомы при исследовании с помощью светового микроскопа представляет следующим образом. Каждая хромосома состоит из двух хроматид, спирально закрученными и располагающихся параллельно оси хромосомы. Для прокрашивающихся в интерфазном яде участков хромосом используют термин "хромонема" - красящая нить. Утолщения на хромонемах получили название хромомер. Особенность вышеописанного строения хромосом зависит от уровня меняется при переходе от интерфазного состояния хромосом к метафазному
Первый, получивший название нуклеосомного, определяет скручивание ДНК по поверхности гистоновой сердцевины. Второй - объединение нескольких нуклеосом (до 10) в бусину - называется нуклеомерный. Третий уровень - объединение скрепками из негистоновых белков фибрилл дезоксирибонуклеопротеида в петлевой домен, называемый хромомером. Четвертый - образование хромонем. Далее, по-видимому, хромонема укладывается в виде спирали в хроматиде, хотя весьма вероятно, что это еще один уровень - "петлистых структур".
2. Мейоз как цитологическая основа образования и развития половых клетокМейоз, или редукционное деление, - особый тип деления клеток, характерный только для спорогенных тканей. При этом число хромосом в дочерних клетках уменьшается вдвое, т.е. происходит редукция числа хромосом. Мейоз предшествует интерфаза, которая аналогична таковой при митозе. В S-период интерфазы происходит редупликация хромосом, поэтому хромосомы, вступающие в процесс мейотического деления, состоят из двух хроматид. Мейоз состоит двух ядерных делений, которое следует одно за другим. При первом делении (мейоз II) происходит редукция числа хромосом, т.е. число хромосом в клетке уменьшается в два раза. Второе деление (мейоз II) протекает по типу митоза. Как и митоз, первое и второе деление мейоз подразделяют на следующие фазы: профаза, метафаза, анафаза и телофаза. Соответственно эти фазы обозначают: метафаза I, метафаза II, анафаза I и т.д.
Мейоз I начинается с профазы I. Это наиболее продолжительная фаза мейоза, которая, в свою очередь, подразделяются на стадии лептотена, зиготена, пахитена, диплотена и диакинез.
На стадии лептотены в ядре появляются слабоспирализованные хромосомы. Постепенно они приобретают нитевидную форму.
Зиготена начинается с постепенно попарного соединения (конъюгации, синапсиса) по длине параллельно уложенных гомологичных хромосом. Соединение попарно хромосомы образуют биваленты. В связи с тем, что перед началом мейоза произошла редупликация хромосом, каждый бивалент состоит из четырех хроматид. Функцию синапсиса выполняет синаптонемный комплекс (СК) - белковое образование, входящие в состав бивалента и имеющие вид трехслойной ленты, располагающейся между конъюгирующими хромосомами. СК формируются постепенно по принципу застежки - молнии на протяжении всей стадии зиготены. Образование бивалентов создает предпосылки для возможности обмена гомологичными участками между гомологичными хромосомами (кроссинговера), что представляет важное генетическое событие. В то же время продолжается процесс конденсации хромосом.
Пахитена - это стадия, на которой СК сформирован по всей длине хромосом (стадия стабильного синапсиса). Она характеризуется продолжающимся утолщением хромосом в результате непрерывной конденсации хроматина. На этой стадии происходит обмен гомологичным участками хроматид (кроссинговер) и, как следствие, рекомбинация сцепленных генов.
На следующей за пахитеной стадии, получившей название диплотены, продолжается конденсация хромосом, но при этом начинается процесс расхождение гомологичных хромосом, которые удерживаются в точках обмена участками, возникшими при кроссинговере. Они получили название хиазм.
Диакинез - последняя стадия профазы I. Она характеризуется максимальной конденсации хромосом. Исчезает ядрышко, а биваленты располагаются по периферии ядра. При этом гомологичные хромосомы удерживаются в составе бивалентов благодаря хиазмам.
Далее следует метафаза I. Ее началу соответствует распад оболочки ядра и формирование веретена деления. Биваленты располагаются в экваториальной плоскости.
Анафаза I - стадия, на которой гомологичные хромосомы расходится к полюсам. В результате число хромосом во вновь образующейся клетке (n) будет в два раза меньше, чем в родительской (2n). В этом отличие анафаза I мейоза от анафазы митоза.
Окончательное расхождение хромосом к полюсам свидетельствует о том, что началась телофаза I.
За ней у ряда видов следует очень короткий интеркинез, во время которого синтез ДНК и репликация хромосом не происходят, и начинается второе деление мейоза (мейоз II). В этом случае хромосомы не деконденсируются. Однако у некоторых видов растений интерфаза между первым и вторым делением мейоза продолжается довольно долго. В этом случае хромосомы деконденсируются, образуя два вида ядра, разделенные клеточной перегородкой. Второе деление мейоза протекает довольно быстро по типу обычного митоза, но уже в клетках с гаплоидным числом хромосом. В тех случаях, когда интерфаза короткая, профаза II выпадает и второе деление начинается с метафазы II, во время которой происходит образование веретена деления и хромосомы располагаются в экваториальной плоскости. В анафазе II центромеры делятся и начинается расхождение хроматид к полюсам, которое заканчивается на стадии телофазы II. На этой стадии происходит полная деконденсация хроматина, образуются ядра и клеточные перегородки. В конечном итоге в результате мейоза образуется 4 клетки, каждая из которых содержит в ядре гаплоидное (n) число хромосом.
3. Сцепленное с полом наследованиеВ опытах Менделя и других исследователей по изучению закономерностей наследования было установлено, что ход наследования многих признаков не зависит от того, материнским или отцовским организмом вносит тот или другой аллель, т.е. реципрокные скрещивания дают одинаковый результат.
Однако при анализе наследования ряда признаков у раздельнополых организмов оказалось, что некоторые из них передаются своеобразно и явно зависит от пола.
В этих случаях реципрокные скрещивания давали разные результаты. Было высказано предложение о том, что определяющие такие признаки гены находится в половых хромосомах, в то время как гены, определяющие признаки, наследующиеся в соответствии с классическими схемами, локализованы в хромосомы, одинаковых у обоих полов, т.е. в аутосомах.
Этот вывод и его доказательство были получены еще в 1909г.Т. Морганом с сотрудниками. Изучая наследование признаков, он установил у дрозофилы наличие связи определенных генов с половыми хромосомами и таким образом заложил фундамент хромосомной теории наследственности. Кроме генов, определяющих пол, половые хромосомы содержат гены, влияющие на разные признаки, не имеющие отношения к дифференциации пола. При передачи таких генов и наблюдается явление так называемого наследования, сцепленного с полом.
В своих первых опытах Морган использовал мутацию белых глаз. Дикие дрозофилы имеют красные глаза. Красный цвет глаз W доминирует над белыми w (white).
При скрещивании гомозиготной красноглазой самки с белоглазым самцом в F1 глаза у всех мух красные, а в F2 происходило расщепление в равном соотношении на красноглазых и белоглазых мух только среди самцов, а все самки F2 были красноглазые. Соотношение по полу было 1: 1:
P ♀ красноглазая х ♂ белоглазый
F1 ♀ красноглазые, ♂ красноглазые
F2 ♀ красноглазые, Ѕ ♂ красноглазые и Ѕ ♂ белоглазые
В реципрокном скрещивание результаты отличались тем, что уже в F1 надобилось расщепление по цвету глаз, причем все самки были красноглазыми, т.е. похоже на отцов, а все самцы - белоглазыми, т.е. похоже на матерей. Такое наследование называют крисс - кросс, или крест - накрест. В F2 и среди самок, и среди самцов половина особей имела красные глаза, половина - белые:
P ♀ белоглазая х ♂ красноглазый
F1 ♀ красноглазые, ♂ белоглазые
F2 Ѕ ♀ красноглазые и Ѕ ♀ белоглазые
Ѕ ♂ красноглазый и Ѕ ♂ белоглазые
Такое расщепление становится понятным, если допустить, что гены, определяющие окраску глаз, находится в X-хромосоме, а в Y-хромосоме их нет.
В первом случае скрещивание гомозиготной красноглазой самки WW с белоглазым самцом wY приводит к образованию красноглазых гетерозиготных по генам окраски самок (Ww) и красноглазых самцов (WY). В F1 самки образуют два типа гамет: с Х-хромосомой с геном W и с Y-хромосомой, не несущей гена окраски. В соответствии с этим в F2 все самки будут красноглазыми: Ѕ гомозиготы WW и Ѕ гетерозиготы Ww, а самцы Ѕ красноглазые WY и Ѕ белоглазые wY:
P ♀ WW х ♂ wY
Красноглазая Белоглазый
F1 ♀ Ww ♂ WY
Красноглазые Красноглазые
F2 ♀ WW ♀ Ww ♂ WY ♂ wY
Красноглазые Красноглазые Красноглазые Белоглазые
В реципрокном скрещивании результаты получаются другие, так как гомозиготная белоглазая самка ww образует один тип гамет - с Х-хромосомой с геном w, красноглазый самец (WY) - два типа гамет: с Х-хромосомой, несущий ген W, и Y-хромосомой, не несущей окраски. В F1 все самки Ww будут красноглазые, а самцы wY - белоглазые. В F2 появится красноглазые гетерозиготные по гену окраски самки Ww и гомозиготные белоглазые ww; Ѕ самцов получают Х-хромосомы, несущие ген красных глаз, и Ѕ - ген белых глаз wY:
P ♀ ww х ♂ WY
Белоглазая Красноглазый
F1 ♀ Ww ♂ wY
Красноглазые Белоглазые
F2 ♀ Ww ♀ ww ♂ WY ♂ wY
Красноглазые Белоглазые Красноглазые Белоглазые
Из результатов скрещивания следует, что самки могут быть гетерозиготными (Ww) или гомозиготными (WW, ww) по генам окраски глаз. У самцов ген окраски локализован только в Х-хромосоме. Y-хромосому называют в этом случае генетически инертной, т.е. проявляется одна доза гена. Такое состояние называют гемизиготными, т.е. WY - красноглазый самец, wY - белоглазый.
Аналогичным образом наследуется все признаки, определяемые генами, локализованными в Х-хромосомах, и у других организмов, у которых гетерогаметен мужской пол. Так, у человека около 60 генов наследуются сцеплено с Х-хромосомой, в том числе гены, обусловливающие такие заболевания, как гемофилия, цветовая слепота, мускульная дистрофия и др.
Однако установлено, что Y-хромосомы не во всех случаях генетически инертны и их функции не сводят только к роли синаптических партнеров при конъюгации с Х-хромосомы во время мейоза. Известно небольшое число примеров, когда в Y-хромосоме локализованы гены, не имеющие аллелей в Х-хромосоме. Например, у живородящей рыбки лебистуса (гуппи) один из признаков - темное пятно спиной плавнике - обусловлено геном, локализованными в Y-хромосоме, и потому передается только от отца к сыну.
Такие признаки называются голандрическими, т.е. наследуемыми исключительно по мужской линии. У человека, таким образом, наследуется локализованный в Y-хромосоме ген SPY, ответственный за развитие мужской потенции, а также гены, контролирующий размер зубов, развитие кожи перепонки между пальцами ног, волосатость мочек ушей (ихтиоз) и др.
Кроме генов, аллели которых локализованы только либо в Х-, либо в Y-хромосоме, имеются гены, общие для обеих половых хромосом. Такие гены у одного и того же вида наследуется как сцепленные то с Х-, то с Y-хромосомой и проявляются в зависимости от того, в какой из них находится доминантный аллель, а какой - рецессивный.
У разных организмов количество таких общих для Х - и Y-хромосом генов неодинаково, а следовательно, различаются и размеры гомологичных участников половых хромосом.
Специфическая часть Y-хромосомы, не имеющая гомологии с Х-хромосомой, у всех изученных организмов генетически инертна, т.е. содержит очень мало генов.
4. Транскрипция ДНКЭто-перенос генетической информации, закодированной в последовательности пар нуклеотидов, с двуцепочечной молекулы ДНК на одноцепочечную молекулу РНК. При этой матрицей для синтеза РНК служит только одна цепь ДНК, называемая смысловой.
В транскрипции, как и в других матричных процессах, различают три стадии: инициацию, элонгацию и терминацию. Фермент, осуществляющий этот процесс, называют ДНК-зависимой РНК-полимеразой или просто РНК-полимеразой; при этом полимеризация полирибонуклеотида (РНК) происходит в направлении от 5 - к 3 - концу растущей цепи.
Синтез ферментов и других белков, необходимых для жизнедеятельности и развития организмов, происходит в основном на первой стадии интерфазы, до начала репликации ДНК.
В результате транскрипции наследственная информация, записанная в ДНК гена, точно транскрибируется (переписывается) в нуклеотидную последовательность мРНК. Синтез мРНК начинается с участка инициации транскрипции, называемого промотором. Промотор расположен перед геном и включает в себя около 80 пар нуклеотидов (у вирусов и бактерий этот участок соответствует примерно одному витку спирали ДНК и включает около 10 пар нуклеотидов). В нуклеотидных последовательностях промоторов часто встречаются пары АТ, поэтому их называют также ТАТА-последовательностями.
Транскрипция осуществляется с помощью ферментов РНК-полимераз. У эукариот известны три типа РНК-полимераз: I-ответственен за синтез рРНК, II-за синтез мРНК; III-за синтез тРНК и низкомолекулярной рРНК-5S РНК.
РНК-полимераза прочно связывается с промотором и разъединяет нуклеотиды комплементарных цепей. Затем этот фермент начинает двигаться вдоль гена (молекулы ДНК) и по мере разъединения цепей ведет на одну из них (смысловой) синтез мРНК, присоединяя согласно принципу комплементарности аденин к тимину, урацил к аденину, гуанин к цитозин к гуанину. Те участки ДНК, на которых полимераза образовала мРНК, вновь соединяются, а синтезируемая молекула мРНК постепенно отделяется от ДНК. Окончание синтеза мРНК определяется участком остановки транскрипции - терминатором. Нуклеотидные последовательности промотора и терминатора узнаются специальными белками, регулирующими активность РНК-полимеразы.
Перед выходом из ядра к начальной части мРНК (5-концу) присоединяется остаток метилированного гуанина, называемый "колпачком", а к концу мРНК (3-концу) - около 200 остатков адениловой кислоты. В таком виде зрелая мРНК проходит через ядерную мембрану в цитоплазму к рибосоме и соединяется с ней. Полагают, что у эукариот "колпачок" мРНК участвует в связывании её с малой субъединицей рибосомы.
Похожие работы
... число хромосом равно 48, а гаплоидное — 24. Усовершенствованные методы изучения хромосом позволили получить более точные сведения о количестве хромосом в клетках у человека, а также выявить аномалии нормального кариотипа, ответственные за некоторые уродства. Особенно плодотворным оказались два метода: 1. Обработка культуры клеток алкалоидом колхицином, который ведет к накоплению делящихся ...
... . Высказывается также предположение, что мезосомы не принимают активного участия в процессах клеточного метаболизма, но выполняют структурную функцию, обеспечивая компартментализацию прокариотной клетки, т. е. пространственное разграничение внутриклеточного содержимого на относительно обособленные отсеки, что создает более благоприятные условия для протекания определенных последовательностей ...
... веретена, которые также образованы микротрубочками. Центриоли поляризуют процесс деления клетки, обеспечивая расхождение сестринских хроматид (хромосом) в анафазе митоза. генетика клетка онтогенез гибрид Законы Г. Менделя В своих опытах по скрещиванию Мендель применял гибридологический метод. Используя этот метод, он изучал наследование по отдельным признакам, а не по всему комплексу, ...
... признаков в популяции Задачи генетики: 1. В области с/х - выведение новых сортов растений и новых пород животных, а также усовершенствование существующих 2. Медицинская генетика - разработка методов диагностики неследственных заболеваний, разработка их профилактики 3. Генная инженерия 43. Особенности наследования при моногибридном скрещивании, установленные Г.Менделем. ...
0 комментариев