Навигация
Строение и функции органелл клетки. Законы Г. Менделя
82818
знаков
0
таблиц
0
изображений
Федеральное агентство по здравоохранению и социальному развитию ГОУ ВПО Челябинская Государственная Медицинская Академия
Фармацевтический факультет
Контрольная работа по медицинской биологии и генетике
Вариант №2
Выполнила: студентка
1 курса 198 группы
Заочного отделения фармацевтического
Факультета Думанова Н. К.
Проверил(а): доцент
Г.Г. Собенина
Челябинск -2010
Клетка элементарная генетическая и структурно – функциональная единица многоклеточных организмов. Строение и функции органелл клетки
Организм человека состоит из нескольких сотен типов клеток, каждый из которых представлен триллионами индивидуумов. Это нервные, мышечные клетки, клетки желез, крови и другие; каждый тип имеет ряд различных подтипов. Как указывают названия, клетки определенных типов группируются в органы или системы органов, выполняющие специфические функции. Способ их группировки часто играет решающую роль в проявлении функций клеток. Например, поперечнополосатые мышечные клетки похожи друг на друга и обладают способностью к сокращению. Порядок их объединения определяет различия между сотнями различных мышц в теле человека. Типы связи клеток достигают вершины сложности в центральной нервной системе, состоящей из десятков миллиардов клеток, каждая из которых связана с десятками тысяч таких же клеток. Подобные связи устанавливаются между клетками, которые каким-то образом выявляют и распознают друг друга, а затем объединяются вместе. Их ассоциации стабилизируются и поддерживаются рядом внеклеточных структурных элементов, ответственных главным образом за формирование определенной характерной структуры каждого типа ткани.
Наши ближайшие сородичи, другие млекопитающие, по строению и организации похожие на нас, имеют такие же типы клеток. Сходные клеточные типы имеются и у более далеких видов животных. Типичные мышечные или нервные клетки обнаруживаются у рыб, насекомых, моллюсков, червей, но их организация по мере понижения уровня упрощается. Еще ниже, на уровне низших беспозвоночных, характер клеток сам по себе начинает меняться — от многоклеточных организмов к колониям полуавтономных клеток. На самой нижней ступени этой лестницы находятся полностью независимые простейшие, например амебы, состоящие из одной клетки.
Растения также состоят из различных типов клеток, объединенных структурными элементами. Но организация растений отличается от животных. Их структура в основном зависит от запасания солнечной энергии, которая утилизируется в специальных фабриках световой энергии, зеленых хлоропластах. Если их удалить, то останется нечто, напоминающее животную клетку. Как и у животных, у растений имеются различные уровни организации — от наиболее сложных цветковых растений я деревьев до наиболее просто устроенных одноклеточных водорослей. У их не-фотосинтезирующих сородичей, грибов, также имеются ранги сложности, нисходящие от высших грибов до плесени и дрожжей.
Все эти клетки, составляющие животное и растительное царства, построены по одному общему плану. В частности, их тела имеют в своем составе объемную вентральную структуру определенной формы, называемую ядром, и разделены на многочисленные четко выраженные отделения мембранными перегородками. Такие клетки называются эукариотическими (греч. еu — хорошо, полностью и karyon— ядро).
Наряду с эукариотами существует более простая форма живых клеток, называемых прокариотами из-за того, что их ядpo имеет примитивную структуру. К прокариотам относятся бактерии. Их размеры намного меньше, чем у эукариотов, они живут отдельными особями или объединены в примитивные колонии и имеют очень простую внутриклеточную организацию.
Тем не менее, бактерии обитают в колоссальном числе видов животных, они умудрились заселить наиболее негостеприимные участки окружающей среды, в том числе насыщенные парами водоемы, в которых бьют горячие источники, и насыщенную солью воду высыхающих морей. Бактерии имеются повсюду, они выполняют немало весьма важных функций, благодаря которым остатки мертвых организмов превращаются в вещества, которые вновь включаются в круговорот жизни. Без бактерий жизнь эукариотов вскоре бы замерла. Однако некоторые бактерии болезнетворны — они способны внедряться в высшие организмы и вызывать заболевания.
Клетка представляет собой обособленную, наименьшую по размерам структуру, которой присуща вся совокупность свойств жизни и которая может в подходящих условиях окружающей среды поддерживать эти свойства в самой себе, а также передавать их в ряду поколений. Клетка, таким образом, несет полную характеристику жизни. Вне клетки не существует настоящей жизнедеятельности. Поэтому в природе планеты ей принадлежит роль элементарной структурной, функциональной и генетической единицы.
Это означает, что клетка составляет основу строения, жизнедеятельности и развития всех живых форм — одноклеточных, многоклеточных и даже неклеточных. Благодаря заложенным в ней механизмам клетка обеспечивает обмен веществ, использование биологической информации, размножение, свойства наследственности и изменчивости, обусловливая тем самым присущие органическому миру качества единства и разнообразия.
Занимая в мире живых существ положение элементарной единицы, клетка отличается сложным строением. При этом определенные черты обнаруживаются во всех без исключения клетках, характеризуя наиболее важные стороны клеточной организации как таковой.
Клетки многоклеточных организмов, как животных, так и растительных, обособлены от своего окружения оболочкой. Клеточная оболочка, или плазмалемма, животных клеток образована мембраной, покрытой снаружи слоем гликокаликса толщиной 10—20 нм. Основными составляющими гликокаликса служат комплексы полисахаридов с белками (гликопротеины) и жирами (гликолипиды). Изнутри к мембране примыкает кортикальный (корковый) слой цитоплазмы толщиной 0,1—0,5 мкм, в котором не встречаются рибосомы и пузырьки, но в значительном количестве находятся микротрубочки и микрофиламенты, имеющие в своем составе сократимые белки.
Плазмалемма выполняет отграничивающую, барьерную, транспортную и рецепторную функции. Благодаря свойству избирательной проницаемости она регулирует химический состав внутренней среды клетки. В плазмалемме размещены молекулы рецепторов, которые избирательно распознают определенные биологически активные вещества (гормоны). В удержании (заякоривании) этих веществ на клеточной поверхности участвуют белки кортикального слоя. Наличие в оболочке рецепторов дает клеткам возможность воспринимать сигналы извне, чтобы целесообразно реагировать на изменения в окружающей их среде или состоянии организма. В пластах и слоях соседние клетки удерживаются благодаря наличию разного вида контактов, которые представлены участками плазмалеммы, имеющими особое строение.
В клетке выделяют ядро и цитоплазму. Клеточное ядро состоит из оболочки, ядерного сока, ядрышка и хроматина. Функциональная роль ядерной оболочки заключается в обособлении генетического материала (хромосом) эукариотической клетки от цитоплазмы с присущими ей многочисленными метаболическими реакциями, а также регуляции двусторонних взаимодействий ядра и цитоплазмы. Ядерная оболочка состоит из двух мембран, разделенных околоядерным (перинуклеарным) пространством. Последнее может сообщаться с канальцами цитоплазматической сети.
Ядерная оболочка пронизана порами диаметром 80—90 нм. Область поры или поровый комплекс с диаметром около 120 нм имеет определенное строение, что указывает на сложный механизм регуляции ядерно-цитоплазматических перемещений веществ и структур. Количество пор зависит от функционального состояния клетки. Чем выше синтетическая активность в клетке, тем больше их число.
В области порового комплекса начинается так называемая плотная пластинка- белковый слой, подстилающий на всем протяжении внутреннюю мембрану ядерной оболочки. Эта структура выполняет прежде всего опорную функцию, так как при ее наличии форма ядра сохраняется даже в случае разрушения обеих мембран ядерной оболочки. Предполагают также, что закономерная связь с веществом плотной пластинки способствует упорядоченному расположению хромосом в интерфазном ядре.
Основу ядерного сока, или матрикса, составляют белки. Ядерный сок образует внутреннюю среду ядра, в связи с чем он играет важную роль в обеспечении нормального функционирования генетического материала. В составе ядерного сока присутствуют нитчатые, или фибриллярные, белки, с которыми связано выполнение опорной функции: в матриксе находятся также первичные продукты транскрипции генетической информации - гетероядерные РНК (гя-РНК), которые здесь же подвергаются процессингу, превращаясь в м-РНК.
Ядрышко представляет собой структуру, в которой происходит образование и созревание рибосомалъных РНК (рРНК). Гены рРНК занимают определенные участки (в зависимости от вида животного) одной или нескольких хромосом (у человека 13-15 и 21-22 пары)- ядрышковые организаторы, в области которых и образуются ядрышки. Такие участки в метафазных хромосомах выглядят как сужения и называются вторичными перетяжками. С помощью электронного микроскопа в ядрышке выявляют нитчатый и зернистый компоненты. Нитчатый (фибриллярный) компонент представлен комплексами белка и гигантских молекул РНК-предшественниц, из которых затем образуются более мелкие молекулы зрелых рРНК. В процессе созревания фибриллы преобразуются в рибонуклеопротеиновые зерна (гранулы), которыми представлен зернистый компонент.
Хроматиновые структуры в виде глыбок, рассеянных в нуклеоплазме, являются интерфазной формой существования хромосом клетки.
В цитоплазме различают основное вещество (матрикс, гиалоплазма), включения и органеллы. Основное вещество цитоплазмы заполняет пространство между плазмалеммой, ядерной оболочкой и другими внутриклеточными структурами. Обычный электронный микроскоп не выявляет в нем какой-либо внутренней организации. Белковый состав гиалоплазмы разнообразен. Важнейшие из белков представлены ферментами гликолиза, обмена Сахаров, азотистых оснований, аминокислот и липидов. Ряд белков гиалоплазмы служит субъединицами, из которых происходит сборка таких структур, как микротрубочки.
Основное вещество цитоплазмы образует истинную внутреннюю среду клетки, которая объединяет все внутриклеточные структуры и обеспечивает взаимодействие их друг с другом. Выполнение матриксом объединяющей, а также каркасной функции может быть связано с выявляемой с помощью сверхмощного электронного микроскопа микротрабекулярной сети, образованной тонкими фибриллами толщиной 2-3 нм и пронизывающей всю цитоплазму.
Включениями называют относительно непостоянные компоненты цитоплазмы, которые служат запасными питательными веществами (жир, гликоген), продуктами, подлежащими выведению из клетки (гранулы секрета), балластными веществами (некоторые пигменты).
Органеллы - это постоянные структуры цитоплазмы, выполняющие в клетке жизненно важные функции.
Выделяют органеллы общего значения и специальные. К ним относят, например, микроворсинки всасывающей поверхности эпителиальной клетки кишечника, реснички эпителия трахеи и бронхов, синаптические пузырьки, транспортирующие вещества - переносчики нервного возбуждения с одной нервной клетки на другую или клетку рабочего органа, миофибриллы, от которых зависит сокращение мышцы.
К органеллам общего значения относят элементы канальцевой и вакуолярной системы в виде шероховатой и гладкой цитоплазматической сети, пластинчатый комплекс, митохондрии, рибосомы и полисомы, лизосомы, пероксисомы, микрофибриллы и микротрубочки, центриоли клеточного центра. В растительных клетках выделяют также хлоропласты, в которых происходит фотосинтез.
Канальцевая и вакуолярная системы образованы сообщающимися или отдельными трубчатыми или уплощенными (цистерна) полостями, ограниченными мембранами и распространяющимися по всей цитоплазме клетки. Нередко цистерны имеют пузыревидные расширения. В названной системе выделяют шероховатую и гладкую цитоплазматическую сети. Особенность строения шероховатой сети состоит в прикреплении к ее мембранам полисом. В силу этого она выполняет функцию синтеза определенной категории белков, преимущественно удаляемых из клетки, например секретируемых клетками желез. В области шероховатой сети происходит образование белков и липидов цитоплазматических мембран, а также их сборка. Плотно упакованные в слоистую структуру цистерны шероховатой сети являются участками наиболее активного белкового синтеза и называются эргастоплазмой.
Мембраны гладкой цитоплазматической сети лишены полисом. Функционально эта сеть связана с обменом углеводов, жиров и других веществ небелковой природы, например стероидных гормонов (в половых железах, корковом слое надпочечников). По канальцам и цистернам происходит перемещение веществ, в частности секретируемого железистой клеткой материала, от места синтеза в зону упаковки в гранулы. В участках печеночных клеток, богатых структурами гладкой сети, разрушаются и обезвреживаются вредные токсические вещества, некоторые лекарства (барбитураты). В пузырьках и канальцах гладкой сети поперечно-полосатой мускулатуры сохраняются (депонируются) ионы кальция, играющие важную роль в процессе сокращения.
Рибосома - это округлая рибонуклеопротеиновая частица диаметром 20—30 нм. Она состоит из малой и большой субъединиц, объединение которых происходит в присутствии матричной (информационной) РНК (мРНК). Одна молекула мРНК обычно объединяет несколько рибосом наподобие нитки бус. Такую структуру называют полисомой. Полисомы свободно располагаются в основном веществе цитоплазмы или прикреплены к мембранам шероховатой цитоплазматической сети. В обоих случаях они служат местом активного синтеза белка. Сравнение соотношения количества свободных и прикрепленных к мембранам полисом в эмбриональных недифференцированных и опухолевых клетках, с одной стороны, и в специализированных клетках взрослого организма—с другой, привело к заключению, что на полисомах гиалоплазмы образуются белки для собственных нужд (для «домашнего» пользования) данной клетки, тогда как на полисомах гранулярной сети синтезируются белки, выводимые из клетки и используемые на нужды организма (например, пищеварительные ферменты, белки грудного молока).
Пластинчатый комплекс Гольджи образован совокупностью диктиосом числом от нескольких десятков (обычно около 20) до нескольких сотен и даже тысяч на клетку.
Диктиосома представлена стопкой из 3-12 уплощенных дискообразных цистерн, от краев которых отшнуровываются пузырьки (везикулы). Ограниченные определенным участком (локальные) расширения цистерн дают более крупные пузырьки (вакуоли). В дифференцированных клетках позвоночных животных и человека диктиосомы обычно собраны в околоядерной зоне цитоплазмы. В пластинчатом комплексе образуются секреторные пузырьки или вакуоли, содержимое которых составляют белки и другие соединения, подлежащие выводу из клетки. При этом предшественник секрета просекрет), поступающий в диктиосому из зоны синтеза, подвергается в ней некоторым химическим преобразованиям. Он также обособляется (сегрегируется) в виде «порций», которые здесь же одеваются мембранной оболочкой. В пластинчатом комплексе образуются лизосомы. В диктиосомах синтезируются полисахариды, а также их комплексы с белками (гликопротеины) и жирами (гликолипиды), которые затем можно обнаружить в гликокаликсе клеточной оболочки.
Митохондрии - это структуры округлой или палочковидной, нередко ветвящейся формы толщиной 0,5 мкм и длиной обычно до 5—10 мкм.
Оболочка митохондрии состоит из двух мембран, различающихся по химическому составу, набору ферментов и функциям. Внутренняя мембрана образует впячивания листовидной (кристы) или трубчатой (тубулы) формы. Пространство, ограниченное внутренней мембраной, составляет матрикс органеллы.
Главная функция митохондрии состоит в ферментативном извлечении из определенных химических веществ энергии (путем их окисления) и накоплении энергии в биологически используемой форме (путем синтеза молекул аденозинтрифосфата — АТФ). В целом этот процесс называется окислительным фосфорилированием. В энергетической функции митохондрии активно участвуют компоненты матрикса и внутренняя мембрана. Именно с этой мембраной связаны цепь переноса электронов (окисление) и АТФ-синтетаза, катализирующая сопряженное с окислением фосфорилирование АДФ в АТФ. Среди побочных функций митохондрии можно назвать участие в синтезе стероидных гормонов и некоторых аминокислот (глутаминовая).
Лизосомы представляют собой пузырьки диаметром обычно 0,2—0,4 мкм, которые содержат набор ферментов кислых гидролаз, катализирующих при низких значениях рН гидролитическое (в водной среде) расщепление нуклеиновых кислот, белков, жиров, полисахаридов. Их оболочка образована одинарной мембраной, покрытой иногда снаружи волокнистым белковым слоем (на электронограммах «окаймленные» пузырьки). Функция лизосом-внутриклеточное переваривание различных химических соединений и структур.
Первичными лизосомами (диаметр 100 нм) называют неактивные органеллы, вторичными - органеллы, в которых происходит процесс переваривания. Вторичные лизосомы образуются из первичных. Они подразделяются на гетеролизосомы (фаголизосомы) и аутолизосомы (цитолизосомы). В первых переваривается материал, поступающий в клетку извне путем пиноцитоза и фагоцитоза, во вторых разрушаются собственные структуры клетки, завершившие свою функцию. Вторичные лизосомы, в которых процесс переваривания завершен, называют остаточными тельцами (телолизосомы). В них отсутствуют гидролазы и содержится непереваренный материал.
Микротельца составляют сборную группу органелл. Это ограниченные одной мембраной пузырьки диаметром 0,1 —1,5 мкм с мелкозернистым матриксом и нередко кристаллоидными или аморфными белковыми включениями. К этой группе относят, в частности, пероксисомы. Они содержат ферменты оксидазы, катализирующие образование пероксида водорода, который, будучи токсичным, разрушается затем под действием фермента пероксидазы. Эти реакции включены в различные метаболические циклы, например в обмен мочевой кислоты в клетках печени и почек. В печеночной клетке число пероксисом достигает 70-100.
К органеллам общего значения относят также некоторые постоянные структуры цитоплазмы, лишенные мембран. Микротрубочки - трубчатые образования различной длины с внешним диаметром 24 нм, шириной просвета 15 нм и толщиной стенки около 5 нм. Встречаются в свободном состоянии в цитоплазме клеток или как структурные элементы жгутиков, ресничек, митотического веретена, центриолей. Свободные микротрубочки и микротрубочки ресничек, жгутиков и центриолей имеют разную устойчивость к разрушающим воздействиям, например химическим (колхицин). Микротрубочки строятся из стереотипных субъединиц белковой природы путем их полимеризации. В живой клетке процессы полимеризации протекают одновременно с процессами деполимеризации. Соотношением этих процессов определяется количество микротрубочек. В свободном состоянии микротрубочки выполняют опорную функцию, определяя форму клеток, а также являются факторами направленного перемещения внутриклеточных компонентов.
Микрофиламентами называют длинные, тонкие образования, иногда образующие пучки и обнаруживаемые по всей цитоплазме. Существует несколько разных типов микрофиламентов. Актиновые микрофиламенты благодаря присутствию в них сократимых белков (актин) рассматривают в качестве структур, обеспечивающих клеточные формы движения, например амебоидные. Им приписывают также каркасную роль и участие в организации внутриклеточных перемещений органелл и участков гиалоплазмы.
По периферии клеток под плазмалеммой, а также в околоядерной зоне обнаруживаются пучки микрофиламентов толщиной 10 нм - промежуточные филаменты. В эпителиальных, нервных, глиальных, мышечных клетках, фибробластах они построены из разных белков. Промежуточные филаменты выполняют, по-видимому, механическую, каркасную функцию.
Актиновые микрофибриллы и промежуточные филаменты, как и микротрубочки, построены из субъединиц. В силу этого их количество зависит от соотношения процессов полимеризации и деполимеризации.
Для животных клеток, части клеток растений, грибов и водорослей характерен клеточный центр, в состав которого входят центриоли. Ее стенка образована 27 микротрубочками, сгруппированными в 9 триплетов. В функцию центриолей входит образование нитей митотического веретена, которые также образованы микротрубочками. Центриоли поляризуют процесс деления клетки, обеспечивая расхождение сестринских хроматид (хромосом) в анафазе митоза.
генетика клетка онтогенез гибрид
Законы Г. Менделя
В своих опытах по скрещиванию Мендель применял гибридологический метод. Используя этот метод, он изучал наследование по отдельным признакам, а не по всему комплексу, проводил точный количественный учет наследования каждого признака в ряду поколений, изучал характер потомства каждого гибрида в отдельности.
Первый закон Менделя - закон единообразия гибридов первого поколения.
При скрещивании гомозиготных особей, отличающихся по одной паре альтернативных (взаимоисключающих) признаков, все потомство в первом поколении единообразно как по фенотипу, так по генотипу.
Мендель проводил моногибридное скрещивание чистых линий гороха, отличающихся по одной паре альтернативных признаков, например, по цвету горошин (желтые и зеленые).
В качестве материнского растения использовали горох с желтыми семенами (доминантный признак), а отцовского - горох с зелеными семенами (рецессивный признак).
В результате мейоза каждое растение давало один сорт гамет. При мейозе из каждой гомологичной пары хромосом в гаметы отходило по одной хромосоме с одним из аллельных генов (А или а).
В результате оплодотворения парность гомологичных хромосомах восстановилась и образовались гибриды. Все растения имели семена только желтого цвета (по фенотипу) и были гетерозиготными по генотипу.
Гибрид 1-го поколения Аа имел один ген - А от одного родителя, а второй ген - а от другого родителя и проявлял доминантный признак, скрывая рецессивный.
По генотипу весь горох гетерозиготен.
Первое поколение единообразно и проявило признак одного из родителей.
Для записи скрещиваний применяют специальную таблиц предложенную английским генетиком Пеннетом и называемую решеткой Пеннета. По горизонтали выписывают гаметы отцовской особи, по вертикали - материнской. В местах пересечений - неровные генотипы потомков. В таблице число клеток зависит от числа типов гамет, образуемых скрещиваемыми особями.
Решеткой Пеннета удобно пользоваться при ди- и полигибридном скрещивании.
Далее Мендель скрестил гибриды между собой. Второй закон Менделя - закон расщепления гибридов.
При скрещивании гибридов 1-го поколения между собой во втором поколении появляются особи как с доминантными, так и с рецессивными признаками, и происходит расщепление по генотипу отношении 3:1 и 1:2:1 по генотипу.
В результате скрещивания гибридов между собой получит» особи, как с доминантными признаками, так и с рецессивными.
Такое расщепление возможно при полном доминировании.
Основы генетики пола
Фенотипические различия между особями разного пола обусловлены генотипом. Есть правила индивидуальности, постоянства и парности хромосом. Гены находятся в хромосомах. Диплоидный набор хромосом называют кариотипом. В женском и мужском кариотипе 23 пары (46) хромосом. 22 пары хромосом одинаковы. Их называют аутосомами. 23-я пара хромосом - половые хромосомы. В женском кариотипе одинаковые ХХ-половые хромосомы. В мужском организме XY-половые хромосомы. Y-хромосома очень мала и содержит мало генов.
Пол наследуется как менделирующий признак (по законам Менделя).
Сочетание половых хромосом в зиготе определяет пол будущего организма.
При созревании половых клеток в результате мейоза гаметы получают гаплоидный набор хромосом. В каждой яйцеклетке есть 22 аутосомы + Х-хромосома. Пол, который образуют гаметы, одинаковые по половой хромосоме, называют гомогаметным.
Сперматозоиды дают гаметы двух видов: половина содержит 22 аутосомы + Х-половую хромосому, и половина содержит 22 аутосомы + Y-половую хромосому. Пол, образующий разные гаметы, называют гетерогаметным. Пол будущего ребенка определяется в момент оплодотворения и зависит от того, каким сперматозоидом будет оплодотворена данная яйцеклетка. Если яйцеклетка оплодотворена сперматозоидом, имеющим Х-хромосому, развивается женский организм, если Y-хромосому – мужской.
Теоретически вероятность рождения мальчика и девочки равна 1:1 или 50%: 50%. Однако, рождается больше мальчиков, но т.к. мужской организм имеет всего одну Х-хромосому, доминантные и рецессивные) проявляют свое действие, то мужской организм менее жизнеспособен.
Такое определение пола характерно для человека и млекопитающих.
У некоторых насекомых (кузнечики, тараканы) нет Y-хромосомы. Самец имеет одну Х-хромосому, а самка две XX. У пчел самки имеют 2n набор хромосом (32 хромосомы), а самцы - n (16) хромосом. Самки развиваются из оплодотворенных яиц, а самцы из неоплодотворенных. У птиц и бабочек самки гетерогаметны и имеют ZW половые хромосомы, а самцы гомогаметны и имеют ZZ половые хромосомы).
У женщин в соматических клетках, кроме аутосом, присутствуют две половые ХХ-хромосомы. Одна из них выявляется, образуя глыбку хроматина, заметную в интерфазных ядрах при обработке красителями. Это Х-хроматин или тельце Барра. Эта хромосома спирализована и неактивна. Вторая хромосома сохраняет свою активность. В клетках мужского и женского организмов содержится по одной активной Х-хромосоме.
У мужчин тельце Барра не выявляется. Если при мейозе произойдет нерасхождение хромосом, то в одну яйцеклетку попадут две ХХ-хромосомы. При оплодотворении такой яйцеклетки сперматозоидом, зигота будет иметь большее число хромосом.
Клетки, содержащие больше двух Х-хромосом, имеют большее число телец Барра, потому что активна всегда только одна Х-хромосома.
Например, XXX (трисомия по Х-хромосоме) по фенотипу девочка. У нее в ядрах соматических клеток выявляются два тельца Барра. При этом синдроме недоразвиты половые железы, снижен интеллект.
XXY - синдром Клайнфельтера - по фенотипу мальчик. У него выявляется тельце Барра, недоразвиты семенники, наблюдается умственная отсталость.
ХО - моносомия по Х-хромосоме - синдром Шерешевского - Тернера. Это девочка. У нее недоразвиты половые железы, малый рост, тельце Барра отсутствует.
YO - не жизнеспособен.
Признаки, гены которых находятся в половых хромосомах, наследуются сцепленно с полом. Наследование признаков, гены которых находятся в X и Y-хромосомах, называют наследованием, сцепленным с полом. Распределение генов в потомстве должно соответствовать распределению половых хромосом в мейозе и их сочетанию при оплодотворении .
В Y-хромосоме есть ген, определяющий развитие мужского пола, необходимый для дифференцировки семенников. В X-хромосоме таких генов нет, но есть много других генов. Y-xpoмосома очень мала и не содержит многих генов, которые есть в X—хромосоме.
У гетерогаметного пола (мужского) большинство генов, локализованных в Х-хромосоме, находится в гемизиготном состоянии, т.е. не имеют аллельной пары. В мужских организмах любой рецессивный ген, локализованный в одном из негомологичных участков Х-хромосомы, проявляется в фенотипе.
Y-хромосома содержит некоторое количество генов, гомологичных генам Х-хромосомы, например, гены геморрагического диатеза, общей цветной слепоты и др.
У человека известны рецессивные сцепленные с полом признаки, такие как гемофилия, дальтонизм, мышечная дистрофия и др. В X—хромосоме есть ген, определяющий признак свертывания крови. Рецессивный ген вызывает развитие гемофилии.
У женщин две ХХ-хромосомы. Рецессивный признак (гемофилия) проявляется в том случае, если гены, отвечающие за него, находятся в двух Х-хромосомах: XhXn. Если организм гетерозиготен по этим генам, то гемофилия не проявится. В мужском организме одна Х-хромосома. Если в ней ген Н или h, то эти гены обязательно проявят свое действие, потому что Y-хромосома не несет данных генов. Все девочки здоровы. Половина мальчиков больна гемофилией.
Женщина может быть гомозиготна или гетерозиготна по генам, локализованным в Х-хромосоме, но рецессивные гены проявляются только в гомозиготном состоянии.
Если гены находятся в Y-хромосоме (голандрическое наследование), то признаки, ими обусловленные, передаются от отца к сыну. Например, так наследуется волосатость ушей. Y-хромосома у человека контролирует дифференцировку семенников. У мужчин одна Х-хромосома. Все гены, находящиеся в ней, в том числе и рецессивные, проявляются в фенотипе. В этом заключается одна из причин повышенной смертности мужских особей по сравнению с женскими.
Признаки, проявление которых различно у представителей разных полов, или проявляющиеся у одного пола, относятся к признакам, ограниченным полом.
Эти признаки могут определяться генами, расположенными как в аутосомах, так и половых хромосомах. Возможность развития лризнака зависит от пола организма. Например, тембры голоса баритон и бас характерны только для мужчин.
Проявление генов, ограниченных полом, связано с реализацией генотипа в условиях среды целостного организма. Помимо генов, ответственных за развитие вторичных половых признаков, которые в норме работают только у одного из полов, у другого они присутствуют, но "молчат". Функциональную активность целого ряда других генов определяет гормональная деятельность организма. Например, у быков есть гены, контролирующие продукцию молока и его качественные особенности (жирность, содержание белка я др.), но у быков они "молчат", а функционируют только у коров. Потенциальная способность быка давать высокомолочное потомство делает его ценным производителем молочного стада.
Есть признаки, зависимые от пола. Гены, степень проявления которых определяется уровнем половых гормонов, называются генами, зависимыми от пола. Эти гены могут находиться не только в половых хромосомах, но и в любых аутосомах.
Например, ген определяющий облысение, типичное для мужчин, локализован в аутосоме, и его проявление зависит от мужских половых гормонов. У мужчин этот ген действует как доминантный. а у женщин как рецессивный. Если у женщин этот ген в гетерозиготном состоянии, то признак не проявляется. Даже в гомозиготном состоянии у женщин этот признак слабее выражен, чем у мужчин.
Понятие об основных этапах эмбрионального развития (дробления, гаструляция, образование тканей и организмов)
Существуют следующие основные типы онтогенеза: прямой и непрямой. Прямое развитие встречается в двух формах — неличиночного и внутриутробного, непрямое — в виде личиночного.
Личиночный тип развития характеризуется тем, что в развитии организма имеется одна или несколько личиночных стадий. Личинки ведут, активный образ жизни, сами добывают пищу. Для осуществления жизненных функций у них имеется ряд провизорных (временных) органов, отсутствующих во взрослом состоянии. Этот тип развития сопровождается превращением (метаморфозом; см. ниже).
Неличиночный тип развития имеет место у рыб, пресмыкающихся, птиц а также беспозвоночных, яйцеклетки которых богаты питательным материалом (желтком), достаточным для завершения онтогенеза. Для питания, дыхания и выделения у этих зародышей также развиваются провизорные органы.
Внутриутробный тип развития характерен для высших млекопитающих и человека. Яйцеклетки почти не содержат питательного материала. В жизненные функции зародыша осуществляются через материнский организм. В связи с этим из тканей матери и зародыша образуются сложные провизорные органы, в первую очередь плацента. Этот наиболее поздний в филогенетическом отношении тип онтогенеза наилучшим образом обеспечивает выживание зародыша.
Эмбриональный период, или эмбриогенез (греч. embryon - зародыш), начинается образования зиготы. Окончание этого периода при разных типах онтогенеза связано с различными моментами развития: при личиночном типе - выходом из яйцевых оболочек, при неличиночном - с выходом из зародышевых оболочек, при внутриутробном - с моментом рождения.
Эмбриональный период делится на стадии зиготы, дробления, бластулы, образования зародышевых листков, гисто- и органогенеза. Зародыши млекопитающих и человека до зачатков органов принято называть эмбрионом, а в дальнейшем - плодом.
Зигота, образующаяся в результате слияния женской и мужской гамет, представляет собой одноклеточную стадию развития нового организма. Участки цитоплазмы яйца, содержащие зерна желтка, митохондрии, пигменты, видны на живых объектах, поэтому в зиготе удалось проследить значительные перемещения цитоплазмы.
В неоплодотворенных яйцах морского ежа Paracentrotus lividus в корковой области равномерно расположены зерна красного пигмента. После оплодотворения они перемещаются и образуют красный пояс ниже экватора, в то время как анимальный и вегетативный полюса обесцвечиваются. Таким разом, создаются три зоны цитоплазмы: в анимальной части яйца – непигментированная, ниже экватора - пигментированная, на вегетативном полюсе — бесцветная. В дальнейшем из цитоплазматического материала в ней бесцветной зоны формируется эктодермы пигментированной зоны - энтодерма, из нижней — элементы мезодермы.
В зиготах асцидий, земноводных и других животных также обнаружены значительные перемещения цитоплазмы, в результате чего определяются участки, из которых в дальнейшем развиваются те или иные органы и ткани. В это же время появляется ярко выраженная двусторонняя симметрия. Установлено, что и в зиготе млекопитающих и человека также до начала дробления происходит дифференциация цитоплазмы и появляется билатеральная симметрия яйца.
В зиготе осуществляется интенсивный синтез белка, матрицей для которого на начальных стадиях развития служит и РНК, синтезированная время овогенеза, но одновременно синтезируется и новая и РНК.
Начальный этап развития оплодотворенного яйца (зиготы) носит название дробления. Характер дробления обусловлен типом яйцеклетки. В изолецитальном, бедном желтком оплодотворенном яйце ланцетника первая борозда дробления в виде щели начинается на анимальном полюсе и постепенно распространяется в продольном меридиональном правлении к вегетативному, разделяя яйцо на две клетки – 2 бластомера. Вторая борозда проходит перпендикулярно первой — образуются 4 бластомера. Третья борозда проходит экваториально: возникает 8 бластомеров. В результате последующих дроблений в меридиональных и экваториальных плоскостях образуется 16, 32, 64 и т. д. бластомеров. При каждом последующем дроблении клетки становятся мельче, так как бластомеры, не достигнув размеров первоначального, вновь делятся. Однако клетки, расположенные на вегетативном полюсе, несколько крупнее, чем на анимальном.
В процессе дробления увеличивается число бластомеров, однако бластомеры не вырастают до размеров исходной клетки, а с каждым дроблением становятся мельче. К концу периода дробления весь зародыш лишь ненамного крупнее зиготы. Это объясняется тем, что митотические циклы дробящейся зиготы не имеют типичной интерфазы. Во время дробления митотические деления следуют быстро друг за другом.
Дробление яйца заканчивается образованием бластулы (греч. Blastos- росток). Отметим, что в зиготе и бластомерах ядерно-плазменное отношение нарушено в пользу цитоплазмы. В клетках бластулы устанавливается типичное для каждого вида животных ядерно-плазменное отношение. Начиная с бластулы клетки зародыша принято называть не бластомерами, а э мбриональными клетками. У ланцетника бластула обратится по достижении зародышем 128 клеток. В силу накопления продуктов, жизнедеятельности бластомеров между ними появляется полость (бластоцель, или первичная полость). При полном равномерном дроблении у ланцетника) бластула имеет форму пузырька со стенкой в один слой клеток, который назван бластодермой. Стадию бластулы проходят зародыши всех типов животных.
У всех многоклеточных животных следующим за бластулой этапом развития является гаструляция (греч. gastre - чрево сосуда, gaster - желудок), которая представляет собой сложный процесс перемещения эмбрионального материала с образованием двух или трех слоев тела зародыша, называемых зародышевыми листками. В процессе гаструляции следует различать два этапа: 1) образование экто- и энтодермы (двухслойный зародыш); 2) образование мезодермы (трехслойный зародыш). У животных с изолецитальным типом яиц гаструляция идет путем инвагинации, т.е. впячивания. Вегетативный полюс бластулы впячивается внутрь наподобие стенки продырявленного резинового мяча. Противоположные полюса бластодермы почти смыкаются, так бластоцель либо исчезает полностью, либо остается в виде незначительной полости, а из шара возникает двухслойный зародыш.
Внешний слой клеток носит название наружного листка, или э к т дермы (греч. ectos — снаружи, derma — кожа), внутренний слой внутреннего листка, или энтодермы (греч. entoc — внутри). Полость называется гастроцелем, или первичной кишкой, а вход в кишку получил наименование бластопора, или первичного рта. Края его сближаются, образуя верхнюю и нижнюю губы.
Гаструляция происходит не только путем инвагинации. Другими ее способами являются деламинация (расслоение), эпиболия (обрастание) и иммиграция (проникновение внутрь).
Образование гаструлы путем иммиграции характерно для кишечнополостных. Этот способ заключается в массовом активном перемещении клеток бластодермы в бластоцель. Эпиболия встречается, у животных, имеющих телолецитальные яйца. При этом способе гаструляции мелкие клетки анимального полюса, размножаясь, обрастают и покрывают снаружи крупные, богатые желтком клетки вегетативного полюса, которые становятся внутренним слоем. При деламинации клетки зародыша делятся параллельно его поверхности, образуя наружный и внутренний зародышевые листки.
Чаще всего имеет место смешанный тип гаструляции, когда одновременно происходят и впячивание, и обрастание, и миграция. Так протекает, например, гаструляция у земноводных.
На стадии двух зародышевых листков заканчивается развитие губок и кишечнополостных. У всех организмов, относящихся к типам, стоящим на более высоких ступенях эволюции, развиваются три зародышевых листка.
Третий, или средний, зародышевый листок называется мезодермой (греч. mesos — средний), так как он образуется между наружным и внутренним листком.
Различают два основных способа образования мезодермы – телобластический и энтероцельный. Телобластический способ встречается у многих беспозвоночных. Заключается он в том, что вблизи бластопопора с двух сторон первичной кишки во время гаструляции образуется по одной крупной клетке — телобласту. В результате размножения телобластов, от которых отделяются мелкие клетки, формируется мезодерма. Энтероцельный способ характерен для хордовых. В этом случае с двух сторон от первичной кишки образуются выпячивания - карманы (целомические мешки). Внутри карманов находится полость, представляющая собой продолжение первичной кишки (гастроцеля). Целомические мешки полностью отшнуровываются от первичной кишки и разрастаются между экто- и эндодермой. Клеточный материал этих участков дает начало среднему зародышевому листку - мезодерме. Дорсальный отдел мезодермы, лежащий по бокам от нервной трубки и хорды, расчленен на сегменты - сомиты. Вентральный ее отдел образует сплошную боковую пластинку, находящуюся по бокам кишечной трубки. Сомиты дифференцируются на три отдела — медиальный (склеротом), центральный (миотом) и латеральный (дерматом). В вентральной части мезодермальной закладки принято различать нефрогонотом (ножка сомита) и спланхнотом. Закладка спланхнотома разделяется на два листка, между которыми образуется полость. В отличие от бластоцеля она получила название вторичной полости, или целома. Один из листков (висцеральный) граничит с энтодермальной кишечной трубкой, а другой (париентальный) подлежит непосредственно эктодерме.
Дифференцированный на три эмбриональные закладки зародышевый материал дает начало всем тканям и органам развивающегося зародыша. Расположение главнейших из них, так называемых осевых органов, намечается уже в процессе гаструляции.
Каждый зародышевый листок дает начало только определенным органам. Так, из эктодермы развиваются ткани нервной системы (очень рано обособляющиеся в виде нервной пластинки). Нервная система у хордовых закладывается дорсально, т. е. на спинной стороне зародыша. Нервная пластинка в составе эктодермы растет интенсивнее остальных участков и затем прогибается, образуя желобок. Размножение клеток продолжается, края лобка смыкаются, образуется нервная трубка, которая тянется вдоль тела от переднего конца к заднему. На переднем конце нервной трубки путем дальнейшего роста и дифференцировки формируется головной мозг. Отростки нервных клеток центральных отделов нервной системы образуют периферические нервы. Кроме того, из эктодермы развиваются наружный покров кожи - эпидермис и его производные (ногти, волосы, сальные и потовые железы, эмаль зубов, воспринимающие клетки органов зрения, слуха, обоняния и т. п.).
Из энтодермы развивается эпителиальная ткань, выстилающая органы пищеварительной, дыхательной и частично мочеполовой систем, органы желудочно-кишечного тракта, в том числе печень и поджелудочная железа.
Миотом дает начало скелетной мускулатуре, нефрогонотом — органа выделения и половым железам (гонады). Клетки, образующие висцеральные и париетальные листки спланхнотома, являются источником эпителиальная выстилки вторичной полости тела - целома. За счет элементов склеротома развивается хрящевая, костная и соединительная ткань, образующая вокруг хорды осевой скелет. Дерматом дает начало соединительной ткани кожи. спланхнотом - соединительной ткани внутренних органов, кровеносным сосудам, гладкой мускулатуре кишечника, дыхательных и мочеполовых путей. В образовании сердца принимает участие также висцеральный листок спланхнотома. Железы внутренней секреции имеют различное происхождение: одни из них (эпифиз, части гипофиза) развиваются из закладок нервной системы, другие - из эктодермы. Надпочечники и половые железы являются производными мезодермы.
Органогенез завершается в основном к концу эмбрионального период развития. Однако дифференцировка и усложнение органов продолжаются и постэмбриональном онтогенезе. Описанные процессы связаны не только с активным клеточным размножением первичных эмбриональных закладок, но и с их значительным перемещением, изменением формы тела зародыша, образованием отверстий и полостей, а также формированием ряда временных зародышевых (провизорных) органов.
В зависимости от места развития дефинитивного (окончательного) ротового отверстия все типы животного мира принято делить на первичноротых и вторичноротых.
К первичноротым относят организмы, у которых в процессе эмбрионального развития дефинитивное ротовое отверстие образуется на месте первичного рта - бластопора. Это все черви, моллюски и членистоногие. У вторичноротых организмов (иглокожие, погонофоры и хордовые) дефинитивный рот формируется на конце, противоположном бластопору.
Малярийный плазмодий. Систематика, морфология, цикл развития, видовые различия. Борьба с малярией. Задача противомалярийной службы на современном этапе
КЛАСС СПОРОВИКИ (SPOROZOA). ОТРЯД КРОВЯНЫЕ СПОРОВИКИ
К классу Споровиков относят около 3600 видов, ведущих паразитический образ жизни. Они не имеют органелл передвижения; пищеварительных, сократительных вакуолей, питание и выделение происходит осмотически. Жизненный цикл сложный и связан со сменой хозяина.
Бесполое размножение обычно осуществляется в форме шизогонии или множественного деления. При этом ядро делится сразу на несколько частей, затем на столько же частей делится цитоплазма и образуется соответствующее число дочерних особей. Бесполое размножение происходит в теле позвоночных животных и человека, а половое в организме безпозвоночных (малярийного комара).
Паразиты человека относятся к отряду кровяных споровиков. Наибольший интерес представляет малярийный плазмодий, вызывающий заболевание человека - малярию. Описал малярийного плазмодия французский врач А.Лаверан в 1878 г. У человека вызывают заболевание 4 вида малярийных плазмодиев: возбудитель трехдневной малярии {Plasmodium vivax), возбудитель четырехдневной малярии (Plasmodium malariae), возбудитель тропической малярии (Plasmodium falciparum) и возбудитель малярии типа трехдневной (Plasmodium ovale).
Заболевание малярией выражается в повторяющихся с определенной частотой приступах лихорадки, сопровождающихся подъемом температуры,
Малярия распространена в странах с жарким и влажным климатом, расположенных в тропическом и субтропическом поясе, и там, где обитает малярийный комар. Это антропоноз, так как резервуаром может быть только человек. Малярия - облигатно-трансмиссивное заболевание, передается с помощью кровососущих насекомых, природно-эндемичная инфекция.
Заражение человека происходит при укусе самками малярийного комара, содержащими малярийный плазмодий на стадии спорозоита. Спорозоиты имеют размеры 5-8 мкм. Током крови они разносятся по телу и внедряются в клетки печени, где превращаются в шизонтов, размножающихся бесполым путем (шизогонией). В результате этого образуются мерозоиты, которые внедряются в эритроциты, где снова растут и делятся. Цикл развития мерозоитов в эритроцитах повторяется несколько раз.
Правильное чередование приступов при малярии связано с периодичностью шизогонии в эритроцитах. Начало приступа (озноб) совпадает с распадом эритроцитов и поступлением в плазму крови мерозоитов и продуктов их жизнедеятельности, вызывающих интоксикацию организма.
После нескольких циклов бесполого размножения (шизогонии) начинается подготовка к половому процессу. При этом мерозоиты, находящиеся в эритроцитах, дают начало гамонтам (подготовительные стадии образования гамет).
Макрогамонты дадут впоследствии женские половые клетки - макрогаметы, микрогамонты - мужские микрогаметы. В крови человека образования половых клеток не происходит, так как человек - промежуточный хозяин в цикле развития малярийного плазмодия. Дальнейшее развитие плазмодия осуществляется, если гамонты с кровью попадают в желудок малярийного комара при сосании им крови больного человека. Малярийные комары - окончательные хозяева в цикле развития малярийного плазмодия. В желудке комара гамонты превращаются в крупные неподвижные макрогаметы и в мелкие подвижные микрогаметы. Происходит ко-пуляция гамет, в результате чего образуется подвижная зигота (оокинета), которая внедряется в стенку желудка комара и инцистируются на стороне, обращенной в полость тела, превращаясь в ооцисту. Ядро зиготы многократно делится. Ооциста распадается на большое количество (до 10000) спорозоитов. Оболочка спороцисты разрушается и спорозоиты попадают в полость тела комара, а затем в слюнные железы насекомого. Малярийный комар снова может заражать человека. Ни на одной стадии паразит не находится во внешней среде.
Инкубационный период длится от 2 недель до 6 месяцев.
Диагноз ставят при нахождении малярийных плазмодий в крови. Кровь берут непосредственно после приступа малярии.
При выезде в районы, опасные в отношении малярии, в качестве личной профилактики рекомендуется предохраняться от укусов комаров (спать под пологом, сеткой, смазывать кожу отпугивающими комаров средствами). Кроме того, следует принимать внутрь лекарственные противомалярийные препараты, оказывающие профилактическое действие. В нашей стране все больные малярией состоят на специальном учете. Лечение их проводится в обязательном порядке. Противомалярийные препараты выдаются бесплатно. Осуществляются многочисленные мероприятия по борьбе с комарами. Блестящие результаты дало применение ядов, к которым чувствительны насекомые,- инсектицидов, синтезированных химиками.
Профилактика связана с уничтожением мест обитания малярийного комара.
Общие закономерности филогенеза кровеносной системы позвоночных животных. Филогенез систем органов хордовых
Филогенез систем органов хордовых рассмотрен в соответствии с прогрессивным направлением эволюции этого типа животных от подтипа Бесчерепные до класса Млекопитающие. Организация систем органов класса Птицы не описана в связи с тем, что птицы произошли от пресмыкающихся значительно позже млекопитающих и являются боковой ветвью эволюции хордовых.
Необходимым условием существования высокоорганизованных крупных многоклеточных организмов является наличие жидкой подвижной внутренней среды, которая обеспечивает интеграцию организма в целостную систему, выполняя транспортные функции. Эти функции являются основными для кровеносной системы. Конкретная функция кровеносной системы зависит от того, что она транспортирует: питательные вещества, кислород, углекислый газ, другие продукты диссимиляции или гормоны. Кровеносная система всех хордовых замкнутая и состоит из двух основных артериальных сосудов: брюшной и спинной аорт. По брюшной аорте венозная кровь продвигается кпереди, обогащается кислородом в органах дыхания, а по спинной - кзади. Из спинной аорты кровь через систему капилляров возвращается по венам в брюшную аорту. Брюшная аорта или ее часть, периодически сокращаясь, проталкивает кровь по сосудам.
Эволюция общего плана строения кровеносной системы хордовых
У ланцетника (кровеносная система наиболее проста. Круг кровообращения один. По брюшной аорте венозная кровь поступает в приносящие жаберные артерии, которые по количеству соответствуют числу межжаберных перегородок (до 150 пар), где и обогащается кислородом.
По выносящим жаберным артериям кровь поступает в корни спинной аорты, расположенные симметрично с двух сторон тела. Они продолжаются как вперед, неся артериальную кровь к головному мозгу, так и назад. Передние ветви этих двух сосудов являются сонными артериями. На уровне заднего конца глотки задние ветви образуют спинную аорту, которая разветвляется на многочисленные артерии, направляющиеся к органам и распадающиеся на капилляры.
После тканевого газообмена кровь поступает в парные передние или задние кардинальные вены, расположенные симметрично. Передняя и задняя кардинальные вены с каждой стороны впадают в кювьеров проток. Оба кювьеровых протока впадают с двух сторон в брюшную аорту. От стенок пищеварительной системы венозная кровь оттекает по воротной вене печени в печеночный вырост, где формируется система капилляров. Затем капилляры вновь собираются в венозный сосуд - печеночную вену, по которой кровь поступает в брюшную аорту.
Таким образом, несмотря на простоту кровеносной системы в целом, уже у ланцетника имеются основные магистральные артерии, характерные для позвоночных, в том числе для человека: это брюшная аорта, преобразующаяся позже в сердце, восходящую часть дуги аорты и корень легочной артерии; спинная аорта, становящаяся позже собственно аортой, и сонные артерии. Основные вены, имеющиеся у ланцетника, также сохраняются у более высокоорганизованных животных. Так, передние кардинальные вены станут позже яремными венами, правый кювьеров проток преобразуется в верхнюю полую вену, а левый, сильно редуцировавшись, - в коронарный синус сердца. Для того чтобы понять, как это происходит, необходимо сопоставить кровеносные системы всех классов позвоночных животных.
Более активный образ жизни рыб предполагает более интенсивный метаболизм. В связи с этим на фоне олигомеризации их артериальных жаберных дуг в конечном счете до четырех пар в них отмечается высокая степень дифференцировки: жаберные сосуды распадаются на капилляры, пронизывающие жаберные лепестки. В процессе интенсификации сократительной функции брюшной аорты часть ее преобразовалась в двухкамерное сердце, состоящее из предсердия и желудочка и располагающееся под нижней челюстью, рядом с жаберным аппаратом. В остальном кровеносная система рыб соответствует строению ее уланцетника.
В связи с выходом земноводных на сушу и появлением легочного дыхания у них возникает два круга кровообращения. Соответственно этому в строении сердца и артерий появляются приспособления, направленные на разделение артериальной и венозной крови. Перемещение земноводных в основном за счет парных конечностей, а не хвоста обусловливает изменения в венозной системе задней части туловища.
Сердце амфибий расположено каудальнее, чем у рыб, рядом с легкими; оно трехкамерное, но, как и у рыб, от правой половины единственного желудочка начинается единственный сосуд - артериальный конус, разветвляющийся последовательно на три пары сосудов: кожно-легочные артерии, дуги аорты и сонные артерии. Как и у всех более высокоорганизованных классов, в правое предсердие впадают вены большого круга, несущие венозную кровь, в левое - малого с артериальной кровью. При сокращении предсердий в желудочек, внутренняя стенка которого снабжена большим количеством мышечных перекладин, одновременно попадают обе порции крови. Полного их смешения из-за своеобразного строения стенки желудочка не происходит, поэтому при его сокращении первая порция венозной крови поступает в артериальный конус и с помощью спирального клапана, находящегося там, направляется в кожно-легочные артерии. Кровь из середины желудочка, смешанная, поступает таким же образом в дуги аорты, а оставшееся небольшое количество артериальной крови, последней попадающей в артериальный конус, направляется в сонные артерии.
Две дуги аорты, несущие смешанную кровь, огибают сердце и пищевод сзади, образуя спинную аорту, снабжающую все тело, кроме головы, смешанной кровью. Задние кардинальные вены сильно редуцируются и собирают кровь только с боковых поверхностей туловища. Функционально их замещает возникшая заново задняя полая вена, собирающая кровь в основном из задних конечностей. Она располагается рядом со спинной аортой и, находясь позади печени, вбирает в себя печеночную вену, которая у рыб впадала непосредственно в венозный синус сердца. Передние кардинальные вены, обеспечивая отток крови от головы, называют теперь яремными венами, а кювьеровы потоки, в которые они впадают вместе с подключичными венами, - передними полыми венами.
В кровеносной системе пресмыкающихся возникают следующие прогрессивные изменения: в желудочке их сердца имеется неполная перегородка, затрудняющая смешение крови, поступающей из правого и левого предсердий; от сердца отходит не один, а три сосуда, образовавшихся в результате разделения артериального ствола. Из левой половины желудочка начинается правая дуга аорты, несущая артериальную кровь, а из правой -легочная артерия с венозной кровью. Из середины желудочка, в области неполной перегородки, начинается левая дуга аорты со смешанной кровью. Обе дуги аорты, как и у предков, срастаются позади сердца, трахеи и пищевода в спинную аорту, кровь в которой смешанная, но более богата кислородом, чем у земноводных, в связи с тем что до слияния сосудов только по левой дуге течет смешанная кровь. Кроме того, сонные и подключичные артерии с обеих сторон берут начало от правой дуги аорты, в результате чего артериальной кровью снабжается не только голова, но и передние конечности. В связи с появлением шеи сердце располагается еще более каудально, чем у земноводных. Венозная система пресмыкающихся принципиально не отличается от системы вен земноводных.
Прогрессивные изменения кровеносной системы млекопитающих сводятся к полному разделению венозного и артериального кровотоков. Это достигается, во-первых, завершенной четырехкамерностью сердца и, во-вторых, редукцией правой дуги аорты и сохранением только левой, начинающейся от левого желудочка. В результате все органы млекопитающих снабжаются артериальной кровью. В венах большого круга кровообращения также обнаруживаются прогрессивные изменения: возникла безымянная вена, объединяющая левые яремную и подключичную вены с правыми, в результате чего остается лишь одна передняя полая вена, располагающаяся справа. Левый кювьеров проток в виде рудиментарного сосуда sinus coronarius теперь собирает венозную кровь только от миокарда, а непарная и полунепарная вены - рудименты задних кардинальных вен, имеют существенное значение в основном в случаях формирования обходных путей венозного оттока через кава-кавальные анастомозы, формируемые ими.
В эмбриональном развитии млекопитающих и человека рекапитулируют закладки сердца и основных кровеносных сосудов предковых классов.
Сердце закладывается на первых этапах развития в виде недифференцированной брюшной аорты, которая за счет изгибания, появления в просвете перегородок и клапанов, становится последовательно двух-, трех- и четырехкамерным. Однако рекапитуляции здесь неполны в связи с тем, что межжелудочковая перегородка млекопитающих формируется иначе и из другого материала по сравнению с рептилиями. Поэтому можно считать, что четырехкамерное сердце млекопитающих формируется на базе трехкамерного сердца, а межжелудочковая перегородка является новообразованием, а не результатом доразвития перегородки пресмыкающихся. Таким образом, в филогенезе сердца позвоночных проявляется девиация: в процессе морфогенеза этого органа у млекопитающих рекапитулируют ранние филогенетические стадии, а затем развитие его идет в ином направлении, характерном лишь для этого класса.
Интересно, что место закладки и положение сердца в филогенетическом ряду позвоночных полностью рекапитулируют у млекопитающих и человека. Так, закладка сердца у человека осуществляется на 20-е сутки эмбриогенеза, как у всех позвоночных, позади головы. Позже за счет изменения пропорций тела, появления шейной области, смещения легких в грудную полость осуществляется и перемещение сердца в переднее средостение. Нарушения развития сердца могут выражаться как в возникновении аномалий строения, так и места его положения. Возможно сохранение к моменту рождения двухкамерного сердца. Этот порок совершенно не совместим с жизнью.
Чаще встречаются дефекты межпредсердной перегородки (1 случай на 1000 рождений), межжелудочковой перегородки (2,5—5 случаев на 1000 рождений), вплоть до трехкамерного сердца с одним общим желудочком. Известен и такой порок, как шейная эктопия сердца, при которой оно находится в шейной области. Этот порок связывают с задержкой сердца в области его первоначальной закладки. При этом ребенок обычно погибает сразу после рождения. Перечисленные пороки сердца наиболее часто встречаются не в изолированном виде, а в комплексе с другими аномалиями сердца, сосудов, а нередко и других органов. Это свидетельствует о том, что в морфогенезе сердца большое значение имеют онтогенетические корреляции. Состояние больных при таких пороках зависит от того, насколько сильно нарушается гемодинамика и осуществляется смешение крови в кровеносном русле.
Филогенез артериальных жаберных дуг
В связи с тем что основные артериальные сосуды у млекопитающих и человека формируются на базе закладок жаберных артерий, проследим их эволюцию в филогенетическом ряду позвоночных. В эмбриогенезе абсолютного большинства позвоночных закладывается шесть пар артериальных жаберных дуг, соответствующих шести парам висцеральных дуг черепа. В связи с тем что две первые пары висцеральных дуг включаются в состав лицевого черепа, две первые артериальные жаберные дуги быстро редуцируются. Оставшиеся четыре пары функционируют у рыб как жаберные артерии. У наземных позвоночных 3-я пара жаберных артерий теряет связь с корнями спинной аорты и несет кровь к голове, становясь сонными артериями. Сосуды 4-й пары достигают наибольшего развития и вместе с участком корня спинной аорты во взрослом состоянии становятся дугами аорты - основными сосудами большого круга кровообращения.
У земноводных и пресмыкающихся оба сосуда развиты и принимают участие в кровообращении. У млекопитающих также закладываются оба сосуда 4-й пары, а позже правая дуга аорты редуцируется таким образом, что от нее остается лишь небольшой рудимент - плечеголовной ствол. Пятая пара артериальных дуг в связи с тем, что она функционально дублирует четвертую, редуцируется у всех наземных позвоночных, кроме хвостатых амфибий. Шестая пара, которая снабжает венозной кровью кроме жабр еще и плавательный пузырь, у кистеперых рыб становится легочной артерией.
В эмбриогенезе человека рекапитуляции артериальных жаберных дуг происходят с особенностями: все шесть пар дуг никогда не существуют одновременно. В то время, когда две первые дуги закладываются, а затем перестраиваются, последние пары сосудов еще не начинают формироваться. Кроме того, пятая артериальная дуга уже закладывается в виде рудиментарного сосуда, присоединенного обычно к 4-й паре, и редуцируется очень быстро.
Из атавистических пороков развития сосудов, развивающихся из артериальных жаберных дуг, остановимся на следующих: с частотой 1 случай на 200 вскрытий детей, умерших от врожденных пороков сердца, встречается персистирование обеих дуг аорты 4-й пары. При этом обе дуги, так же как у земноводных или пресмыкающихся, срастаются позади пищевода и трахеи, образуя нисходящую часть спинной аорты. Порок проявляется нарушением глотания и удушьем. Несколько чаще (2,8 случая на 200 вскрытий) встречается нарушение редукции правой дуги аорты с редукцией левой. Эта аномалия часто клинически не проявляется.
Наиболее частый порок (0,5—1,2 случая на 1000 новорожденных) - персистирование артериального, или боталлова, протока, представляющего собой часть корня спинной аорты между 4-й и 6-й парами артерий слева. Проявляется сбросом артериальной крови из большого круга кровообращения в малый. Очень тяжелый порок развития - персистирование первичного эмбрионального ствола, в результате которого из сердца выходит только один сосуд, располагающийся обычно над дефектом в межжелудочковой перегородке. Он обычно заканчивается смертью ребенка. Нарушение дифференцировки первичного эмбрионального ствола может привести к такому пороку развития, как транспозиция сосудов - отхождение аорты от правого желудочка, а легочного ствола - от левого, что встречается в 1 случае на 2500 новорожденных. Этот порок обычно несовместим с жизнью.
Рекапитуляции проявляются и в эмбриональном развитии крупных вен человека. При этом возможно формирование атавистических пороков развития. Среди пороков развития венозного русла укажем на возможность персистирования двух верхних полых вен. Если обе они впадают в правое предсердие, аномалия клинически не проявляется. При впадении левой полой вены в левое предсердие происходит сброс венозной крови в большой круг кровообращения. Иногда обе полые вены впадают в левое предсердие. Такой порок несовместим с жизнью. Данные аномалии встречаются с частотой 1% от всех врожденных пороков сердечно-сосудистой системы.
Очень редкая врожденная аномалия - неразвитие нижней полой вены. Отток крови от нижней части туловища и ног осуществляется в этом случае через коллатерали непарной и полунепарной вен, являющихся рудиментами задних кардинальных вен.
Популяция. Ее экологические и генетические характеристики
В природных условиях организмы одного вида расселены неравномерно. Имеет место чередование участков повышенной и пониженной концентрации особей. В результате вид распадается на группировки или популяции, соответствующие зонам более плотного заселения. «Радиусы индивидуальной активности» отдельных особей ограничены. Так, виноградная улитка способна преодолеть расстояние в несколько десятков метров, ондатра - в несколько сотен метров, песец - в несколько сотен километров. Благодаря этому размножение (репродуктивные ареалы) в основном приурочено к территориям с повышенной плотностью организмов.
Вероятность случайных скрещиваний (панмиксии), обусловливающих из поколения в поколение эффективную перекомбинацию генов, внутри «сгущений» оказывается выше, чем в зонах между ними и для вида в целом. Таким образом, в репродуктивном процессе генофонд вида представлен генофондами популяций.
Популяцией называют минимальную самовоспроизводящуюся группу особей одного вида, населяющих определенную территорию (ареал) достаточно долго (в течение многих поколений). В популяции фактически осуществляется сравнительно высокий уровень панмиксии, и она в определенной степени отделена от других популяций той или иной формой изоляции.
Экологически популяция характеризуется величиной, оцениваемой по занимаемой территории (ареалу), численности особей, возрастному и половому составу. Размеры ареала зависят от радиусов индивидуальной активности организмов данного вида и особенностей природных условий на соответствующей территории. Численность особей в популяциях организмов разных видов различается. Так, количество стрекоз Leucorrhinia albifrons в популяции на одном из подмосковных озер достигало 30 000, тогда как численность земляной улитки Cepaea nemoralis оценивалась в 1000 экземпляров. Существуют минимальные значения численности, при которых популяция способна поддерживать себя во времени. Сокращение численности ниже этого минимума приводит к вымиранию популяции.
Величина популяции постоянно колеблется, что зависит от изменений экологической ситуации. Так, осенью благоприятного по кормовым условиям года популяция диких кроликов на одном из островов у юго-западного побережья Англии состояла из 10 000 особей. После холодной малокормной зимы число особей снизилось до 100.
Возрастная структура популяций организмов разных видов варьирует в зависимости от продолжительности жизни, интенсивности размножения, возраста достижения половой зрелости. В зависимости от вида организмов она может быть то более, то менее сложной. Так, у стадных млекопитающих, например дельфинов белух Delphinapterus leucas, в популяции одновременно находятся детеныши текущего года рождения, подросший молодняк прошлого года рождения, половозрелые, но, как правило, не размножающиеся животные в возрасте 2—3 лет, взрослые размножающиеся особи в возрасте 4—20 лет. С другой стороны, у землероек Sorex весной рождаются 1—2 приплода, вслед за чем взрослые особи вымирают, так что осенью вся популяция состоит из молодых неполовозрелых животных.
Половой состав популяций обусловливается эволюционно закрепленными механизмами формирования первичного (на момент зачатия), вторичного (на момент рождения) и третичного (во взрослом состоянии) соотношения полов. В качестве примера рассмотрим изменение полового состава популяции людей. На момент рождения оно составляет 106 мальчиков на 100 девочек, в возрасте 16—18 лет выравнивается, в возрасте 50 лет насчитывает 85 мужчин на 100 женщин, а в возрасте 80 лет — 50 мужчин на 100 женщин.
Генетические характеристики популяции
Генетически популяция характеризуется ее генофондом (аллело-фондом). Он представлен совокупностью аллелей, образующих генотипы организмов данной популяции. Генофонды природных популяций отличает наследственное разнообразие (генетическая гетерогенность, или полиморфизм), генетическое единство, динамическое равновесие доли особей с разными генотипами.
Наследственное разнообразие заключается в присутствии в генофонде одновременно различных аллелей отдельных генов. Первично оно создается мутационным процессом. Мутации, будучи обычно рецессивными и не влияя на фенотипы гетерозиготных организмов, сохраняются в генофондах популяций в скрытом от естественного отбора состоянии. Накапливаясь, они образуют резерв наследственной изменчивости. Благодаря комбинативной изменчивости этот резерв используется для создания в каждом поколении новых комбинаций аллелей. Объем такого резерва огромен. Так, при скрещивании организмов, различающихся по 1000 локусов, каждый из которых представлен десятью аллелями, количество вариантов генотипов достигает 101000, что превосходит число электронов во Вселенной.
Генетическое единство популяции обусловливается достаточным уровнем панмиксии. В условиях случайного подбора скрещивающихся особей источником аллелей для генотипов организмов последовательных поколений является весь генофонд популяции. Генетическое единство проявляется также в общей генотипической изменчивости популяции при изменении условий существования, что обусловливает как выживание вида, так и образование новых видов.
Естественный отбор, формы естественного отбора, его значение для видообразования
В природных популяциях организмов, размножающихся половым способом, существует большое разнообразие генотипов и, следовательно, фенотипов. Благодаря индивидуальной изменчивости в условиях конкретной среды обитания приспособленность разных генотипов (фенотипов) различна. В эволюционном контексте приспособленность определяют как произведение жизнеспособности в данной среде, обусловливающей большую или меньшую вероятность достижения репродуктивного возраста, на репродуктивную способность особи. Различия между организмами по приспособленности, оцениваемой передачей аллелей следующему поколению, выявляются в природе с помощью естественного отбора. Главный результат отбора заключается не просто в выживании более жизнеспособных, а в относительном вкладе таких особей в генофонд дочерней популяции.
Необходимой предпосылкой отбора является борьба за существование - конкуренция за пищу, жизненное пространство, партнера для спаривания. Естественный отбор происходит на всех стадиях онтогенеза организмов. На дорепродуктивных стадиях индивидуального развития, например в эмбриогенезе, преобладающим механизмом отбора служит дифференциальная (избирательная) смертность. В конечном итоге отбор обеспечивает дифференциальное (избирательное) воспроизведение (размножение) генотипов. Благодаря естественному отбору аллели (признаки), повышающие выживаемость и репродуктивную способность, накапливаются в ряду поколений, изменяя генетический состав популяций в биологически целесообразном направлении. В природных условиях естественный отбор осуществляется исключительно по фенотипу. Отбор генотипов происходит вторично через отбор фенотипов, которые отражают генетическую конституцию организмов.
Как элементарный эволюционный фактор естественный отбор действует в популяциях. Популяция является полем действия, отдельные особи - объектами действия, а конкретные признаки - точками приложения отбора.
Эффективность отбора по качественному и количественному изменению генофонда популяции зависит от величины давления и направления его действия. Величину давления отбора выражают коэффициентом отбора S, который характеризует интенсивность устранения из репродуктивного процесса или сохранения в нем соответственно менее или более приспособленных форм по сравнению с формой, принятой за стандарт приспособленности. Так, если некий локус представлен аллелями A1 и А2, то популяция по генотипам делится на три группы: A1A1; A1A2; А2А2. Обозначим приспособленность этих генотипов Wo, Wi, W2. Выберем в качестве стандарта первый генотип, относительная приспособленность которого максимальна и равна 1. Тогда приспособленности других генотипов составят доли этого стандарта:
A1A1A1A2A2A2
WoW1W2
или Wo/Wo = 1, W1/W0 = 1 – S1, W2/W0 = 1 - S2.
Величины S1 и S2 означают пропорциональное снижение в очередном поколении воспроизводства генотипов А1А2 и А2А2 сравнительно с генотипом A1A1.
Отбор особенно эффективен в отношении доминантных аллелей при условии их полного фенотипического проявления и менее эффективен в отношении рецессивных аллелей, а также в условиях неполной пенетрантности. На результат отбора влияет исходная концентрация аллеля в генофонде. При низких и высоких концентрациях отбор происходит медленно. Изменение доли доминантного аллеля в сравнении с рецессивным при коэффициенте отбора 0,01 приведено ниже.
Возрастание, %Число поколений
0,1-1250
1-50500
50-985000
98-99,990000
В теории, упрощая ситуацию, допускают, что отбор через фенотипы действует на генотипы благодаря различиям в приспособительной ценности отдельных аллелей. В реальной жизни приспособительная ценность генотипов зависит от влияния на фенотип и взаимодействия всей совокупности генов. Оценка величины давления отбора по изменению концентрации отдельных аллелей технически часто невыполнима. Поэтому расчет проводят по изменению концентрации организмов определенного фенотипа.
Пусть в популяции присутствуют организмы двух фенотипических классов А и В в отношении СА/СВ = U1. Вследствие различий по приспособленности происходит естественный отбор (селекция), который изменяет соотношение особей с фенотипами А и В. В следующем поколении оно станет СА/СВ = U2 = U1 (1 + S), где S — коэффициент отбора. Отсюда S = U2/U1 — 1. При селективном преимуществе фенотипа A U2 > U1, a S > 0. При селективном преимуществе фенотипа В U2 < U1 и S < 0. Если приспособленность фенотипов А и В сопоставима и U2 = U1, a S=0. В рассмотренном примере при S > 0 отбор сохраняет в популяции в ряду поколений фенотипы А и устраняет фенотипы В, при S < 0 имеет место обратная тенденция. Отбор, сохраняющий определенные фенотипы, по своему направлению является положительным, тогда как отбор, устраняющий фенотипы из популяции,- отрицательным.
В зависимости от результата различают стабилизирующую, движущую и дизруптивную формы естественного отбора. Стабилизирующий отбор сохраняет в популяции средний вариант фенотипа или признака. Он устраняет из репродуктивного процесса фенотипы, уклоняющиеся от сложившейся адаптивной «нормы», приводит к преимущественному размножению типичных организмов. Так, сотрудник одного из университетов США подобрал после снегопада и сильного ветра 136 оглушенных воробьев Passer domesticus. Из них 72 выживших воробья имели крылья средней длины, тогда как 64 погибшие птицы были либо длиннокрылыми, либо короткокрылыми. Стабилизирующая форма соответствует консервативной роли естественного отбора. При относительном постоянстве условий среды благодаря этой форме сохраняются результаты предшествующих этапов эволюции.
Движущий (направленный) отбор обусловливает последовательное изменение фенотипа в определенном направлении, что проявляется в сдвиге средних значений отбираемых признаков в сторону их усиления или ослабления. При смене условий обитания благодаря этой форме отбора в популяции закрепляется фенотип, более соответствующий среде. После того как новое значение признака придет в оптимальное соответствие условиям среды, движущая форма отбора сменяется стабилизирующей. Примером такого отбора является замещение в популяции гавани Плимут (Англия) крабов Carcinus maenas с широким головогрудным щитком животными с узким щитком в связи с увеличением количества ила.
Направленный отбор составляет основу искусственного отбора. Так, в одном эксперименте на протяжении ряда поколений из популяции шестинедельных мышеи отбирали для скрещивания наиболее тяжелых и наиболее легких животных. Избирательное воспроизведение по признаку массы тела привело к образованию двух самостоятельных популяций, соответственно с возрастающей и убывающей массой тела. По окончании опыта, занявшего 11 поколений, ни одна из этих популяций не вернулась к первоначальной массе.
Дизруптивный (разрывающий) отбор сохраняет несколько разных фенотипов с равной приспособленностью. Он действует против особей со средним или промежуточным значением признаков. Так, в зависимости от преобладающего цвета почвы улитки Cepaea nemoralis имеют раковины коричневой, желтой, розовой окраски. Дизруптивная форма отбора «разрывает» популяцию по определенному признаку на несколько групп. Она поддерживает в популяции состояние генетического полиморфизма.
В зависимости от формы отбор сокращает масштабы изменчивости, создает новую или сохраняет прежнюю картину разнообразия. Как и другие элементарные эволюционные факторы, естественный отбор вызывает изменения в соотношении аллелей в генофондах популяций. Особенность его действия состоит в том, что эти изменения направлены. Отбор приводит генофонды в соответствие с критерием приспособленности. Он осуществляет обратную связь между изменениями генофонда и условиями обитания, накладывает на эти изменения печать биологической целесообразности (полезности). Естественный отбор действует совместно с другими эволюционными факторами. Поддерживая генотипическое разнообразие особей в ряду поколений, мутационный процесс, а также популяционные волны, комбинативная изменчивость создают для него необходимый материал. Естественный отбор нельзя рассматривать как «сито», сортирующее генотипы по приспособленности. В эволюции ему принадлежит творческая роль. Исключая из размножения генотипы с малой приспособительной ценностью, сохраняя благоприятные генные комбинации разного масштаба, он преобразует картину генотипической изменчивости, складывающуюся первоначально под действием случайных факторов, в биологически целесообразном направлении. Результатом творческой роли отбора является процесс органической эволюции, идущей в целом по линии прогрессивного усложнения морфофизио-логической организации (арогенез), а в отдельных ветвях - пути специализации (аллогенез).
Экологические факторы и их взаимодействие. Примеры
Живые организмы находятся в постоянном взаимодействии друг с другом и факторами неживой природы. Видовой состав данной местности определяется историческими и климатическими условиями, а взаимоотношение организмов друг с другом и с окружающей средой - характером питания.
Среда - это комплекс окружающих условий, которые действуют на особь, популяцию, сообщество (биоценоз) в месте их обитания.
Жизнь возникла в воде. Из водной среды организмы вышли на сушу и освоили почвенно-воздушную и почвенную среды. Специфической средой обитания для ряда организмов стали другие живые организмы. Так возникли паразитические формы, симбионты, комменсалы. Любой организм существует в соседстве с другими живыми существами, на которые он влияет и в то же время находится под их воздействием. Оказывают влияние на живые организмы такие факторы, как свет, температура, влажность, климат и др. Элементы среды обитания, которые способны оказывать прямое или косвенное влияние на живые организмы хотя бы на одной из стадий их индивидуального развития, называются экологическими факторами. Среди них различают три группы факторов.
Похожие работы
... . ЛИТЕРАТУРА: 1. Курс низших растений //Под ред. М. В. Горленко – М.: Высш. школа, 1981. 2. Жизнь растений, т. 1–3, - М.: Просвещение, 1974-77. 3. Н. Грин, У. Стаут, Д. Тейлор. Биология, т. 1-3. – М.: Мир, 1990. 4. Н.П.Горбунова , Е.С.Клюшникова, Н.А.Комарницкий. и др. Малый практикум по низшим растениям. – М.: Высш. школа, 1976. 5. Русанов В. А. Низшие растения. Методические указания ...

... Подводя итог, можно сказать, что именно выделение живой самостоятельной клетки из окружающей среды и стало толчком к началу эволюции жизни на земле и роль клетки в развитии всего живого является главенствующей. 4. Основные проблемы цитологии Перед современной цитологией стоит ряд серьёзных задач, важнейших для общества. Если так и не решенный вопрос о происхождении жизни и выделении живой ...
... . Однако эти модификации не наследуются, потому что гены, отвечающие за развитие растений, не меняются в ответ на изменения температуры, влажности, характера питания. Вывод, что признаки, приобретенные в течение жизни организмов, не наследуются, сделал крупный немецкий биолог А. Вейсман. Иногда модификационная изменчивость называется ненаследственной. Это верно в том смысле, что модификации ...
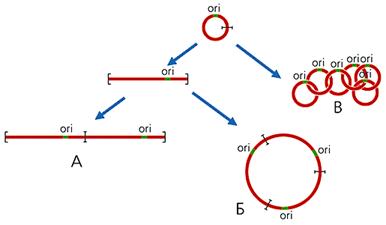
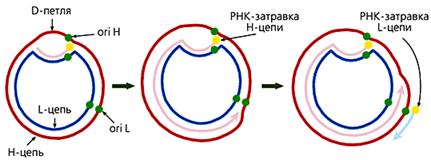
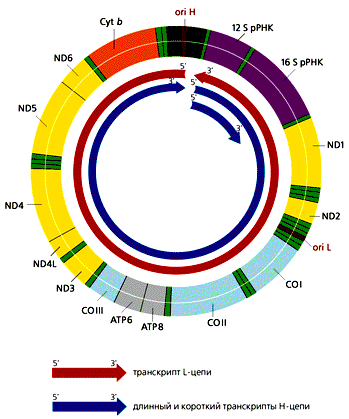
... многокомпонентной системы, необходимой становится ликвидация даного компонента с последующим анализом произошедших изменений. Так как темой даного реферата является указание роли материнского генома для развития потомка, логично было бы узнать о последствиях нарушений в составе митохондриального генома вызванных различными факторами. Инструментом для изучения вышеуказанной роли оказался ...
0 комментариев