Доклад по биологии
« Эволюционное учение »
02.06.2009г
Кепдип Нжог Аший Флориян
Гр. 8323 б
МИКРОЭВОЛЮЦИЯ
Микроэволюция: это совокупность пусковых эволюционных процессов, протекающих внутри вида, в пределах отдельных или смежных популяций. При этом популяции рассматриваются как элементарные эволюционные структуры; мутации, лежащие в основе наследственной изменчивости, — как элементарный эволюционный материал, а мутационный процесс, волны жизни, разные формы изоляции и естественный отбор — как элементарные эволюционные факторы. Под давлением этих факторов происходит изменение генотипического состава популяции — ведущий пусковой механизм эволюционного процесса. Ранее термин «Микроэволюция» употреблялся некоторыми эволюционистами для обозначения изменчивости и формообразования внутри вида и противопоставлялся макроэволюции. Современное учение о Микроэволюция развилось после синтеза генетики с классическим дарвинизмом, начало чему было положено работами советского генетика С. С. Четверикова (1926) и английского генетика Р. А. Фишера (1930). По современным воззрениям (иногда называемым «синтетической теорией эволюции»), все основные пусковые механизмы эволюции (на всех её уровнях) протекают внутри видов, т. е. на микроэволюционном уровне. Микроэволюция завершается видообразованием, т. е. возникновением видов, репродуктивно изолированных от исходных и других близких видов. Поэтому нет принципиальных различий между Микроэволюция и макроэволюцией, различающихся лишь временными и пространственными масштабами. Для успеха исследований на микроэволюционном уровне необходим синтез популяционно-генетических опытов, количественных описаний процессов популяционной динамики и экологии, изучения этологических явлений, аналитического применения теоретических положений генетики и, наконец, построения математических моделей внутрипопуляционных и межпопуляционных процессов.
Основные формы организации жизни:
Определение жизни
Вопросы о происхождении жизни, закономерностях исторического развития в различные геологические эпохи всегда интересовали человечество. Понятие жизнь охватывает совокупность всех живых организмов на Земле и условия их существования.
Сущность жизни заключается в том, что живые организмы оставляют после себя потомство. Наследственная информация передается из поколения в поколение, организмы саморегулируются и восстанавливаются при воспроизводстве потомства. Жизнь — это особая качественная, наивысшая форма материи, способная, оставляя потомство, к самовоспроизведению.
Понятию жизнь в разных исторических периодах давались различные определения. Первое научно правильное определение дал Ф. Энгельс: "Жизнь есть способ существования белковых тел, и этот способ существования состоит по своему существу в постоянном самообновлении химических составных частей этих тел". При прекращении процесса обмена веществ между живыми организмами и окружающей средой белки распадаются, и жизнь исчезает.
Опираясь на современные достижения биологической науки, русский ученый М. В. Волькенштейн дал новое определение понятию жизнь: "Живые тела, существующие на Земле, представляют собой открытые, саморегулирующиеся и самовоспроизводящиеся системы, построенные из биополимеров — белков и нуклеиновых кислот". Это определение не отрицает наличие жизни и на других планетах космического пространства. Жизнь называется открытой системой, на что указывает непрерывный процесс обмена веществ и энергии с окружающей средой.
На основании последних научных достижений современной биологической науки дано следующее определение жизни: "Жизнь — это открытые саморегулирующиеся и самовоспроизводящиеся системы совокупностей живых организмов, построенные из сложных биологических полимеров — белков и нуклеиновых кислот".
Основой всего живого считаются нуклеиновые кислоты и белки, так как они функционируют в клетке, образовывают сложные соединения, которые входят в структуру всех живых организмов.
Основные свойства живых организмов
Живые организмы отличаются от неживой природы присущими им свойствами. К характерным свойствам живых организмов относятся: единство химического состава, обмен веществ и энергии, сходство уровней организации. Для живых организмов характерны также размножение, наследственность, изменчивость, рост и развитие, раздражимость, дискретность, саморегуляция, ритмичность и др.
Изменчивость. Мутационный процесс
Мутация — это внезапное наследственное изменение, вызванное резким структурным и функциональным изменением генетического материала. Генетический материал организован в иерархию структурно-функциональных единиц — от молекулярных сайтов внутри гена до целых хромосом и геномов. Соответственно существуют разные типы мутаций — от генных до геномных. Эта глава посвящена в основном генным мутациям.
Внезапные наследственные изменения фенотипа могут быть вызваны не только структурными изменениями генов, но и другими генетическими процессами. Мутации могут быть истинными или ложными. Фенотипические изменения сами по себе не дают представления о тех генетических процессах, которые их вызывают. На основании одних лишь прямых наблюдений трудно различать разные типы истинных и ложных мутаций. Существует также, как мы увидим в дальнейшем, внезапное изменение генетического материала, не вызывающее фенотипического эффекта.
Генные мутацииГенная, или точковая, мутация представляет собой изменение последовательности нуклеотидов в пределах одного гена, приводящее к изменению характера действия гена. Как правило (за одним исключением, которое будет описано в следующем разделе), это молекулярное изменение в гене, которое вызывает фенотипический эффект. Допустим, что какой-то ген содержит в некоторой своей точке кодов, или триплет, ЦТТ, кодирующий одну из аминокислот полипептидной цепи — глутаминовую кислоту. В результате замены всего лишь одного нуклеотида кодом ЦТТ может превратиться в кодон ГТТ, Этот новый кодон обусловливает синтез уже не глутаминовой кислоты, а глутамина, так что в полипептидной цепи, синтезируемой под действием измененного гена, на месте глутаминовой кислоты окажется глутамин. Первоначальная и мутантная молекулы белка отличаются одна от другой, и вполне возможно, что это влечет за собой другие, вторичные, фенотипические различия.
Стабильность генов на протяжении последовательных поколений клеток и особей, а следовательно, и консервативность наследственности обусловлены точностью процесса копирования при репликации гена, Однако процесс копирования несовершенен. Время от времени при копировании возникают ошибки. Генные мутации можно рассматривать как такие ошибки копирования.
Новый мутантный аллель точно реплицируется до тех пор, пока не произойдет следующее мутационное изменение. Таким образом, в результате генной мутации появляется пара или серия гомологичных аллелей. И наоборот, наличие аллельной изменчивости по любому гену в конечном счете означает, что этот ген в то или иное время претерпел мутацию.
Любой ген, входящий в состав генотипа, по-видимому, подвержен мутированию. Во всяком случае в генах, контролирующих весьма разнообразные признаки, наблюдаются мутации. Например, у Drosophila melanogaster известны мутанты со слегка сморщенными крыльями, сильно укороченными крыльями или вообще бескрылые; мутанты с белыми или пурпурными глазами; мутанты с разнообразными изменениями щетинок и т. п. Известен ряд мутантных разновидностей смородинного томата (Lycopersicon pimpinellifolium), различающихся по форме листьев. Биохимические мутации, затрагивающие различные звенья метаболических процессов, хорошо известны у микроорганизмов и имеются, хотя и гораздо менее изучены, у высших организмов.
По степени фенотипического проявления генные мутации варьируют в широком диапазоне — от мутаций со слабыми эффектами до мутаций, вызывающих значительные изменения фенотипа. Эти два экстрёмальных типа называют соответственно малыми мутациями и макромутациями. Хорошо заметные, но не обладающие сильным действием мутации типичны для средней части диапазона. Примерами малых мутаций служат мутанты Drosophila melanogaster со статистически незначительными отклонениями от нормальной жизнеспособности или от нормального числа щетинок. Примером макромутации служит мутант tetraptera у D. melanogaster с четырьмя крыльями вместо двух. Он представляет собой резкое отклонение от двукрылости, характерной для сем. Drosophilidae и для отряда Diptera.
У диплоидных животных и растений значительную долю новых мутаций составляют рецессивные мутации, а гены дикого типа доминируют. Важное следствие рецессивности многих мутантных аллелей заключается в том, что они не подвергаются действию отбора немедленно, но могут сохраняться в диплоидной популяции на протяжении многих поколений.
Типы точковых мутацийТочковые мутации можно разделить на несколько типов в зависимости от характера молекулярного изменения в гене. Здесь мы кратко опишем четыре типа таких мутаций (Wallace, 1981*)
1. Missense-мутация. К этому типу принадлежит мутация, описанная в предыдущем разделе. В одном из триплетов происходит замена одного основания (например, ЦТТ→ГТТ), в результате чего измененный триплет кодирует аминокислоту, отличную от той, которую кодировал прежний триплет.
2. Мутация со сдвигом рамки. Если в последовательность ДНК включается новое основание или пара оснований, то все лежащие за ними триплеты изменяются, что влечет за собой изменение синтезируемого полипептида. Возьмем, например, последовательность АТТ—ТАГ—ЦГА, перед которой включилось основание Т. В результате получится новая последовательность ТАТ—ТТА—ГЦГ—А… К такому же результату приведёт утрата одного из имеющихся оснований.
3. Nonsense-мутация. В результате замены одного основания возникает новый триплет, представляющий собой терминирующий кодон. В генетическом коде имеется три таких триплета. При такой замене синтез полипептидной цепи прекращается в новой (т. е. другой) точке, и соответственно эта цепь отличается своим свойствам от полипептида, который синтез прежде.
4. Синонимическая missence-мутация. Генетический код обладает значительной избыточностью: два или несколько его триплетов кодируют одну и ту же аминокислоту. Поэтому можно ожидать, что в некоторых случаях при замене оснований один триплет заменяется другим — синонимическим, кодирующим ту же аминокислоту. В этом случае, вследствие избыточности кода мы имеем дело с молекулярным изменением в пределах данного гена, которое не вызывает фенотипического эффекта. Такие синонимические мутации, вероятно, довольно обычны.
Частота возникновения мутаций (скорость мутирования)
Некоторые репрезентативные значения частоты возникновения спонтанных мутаций (мутаций в обычном смысле слова, т.е. вызывающих фенотипические эффекты) приведены в табл. 6.1. Следует отметить, во-первых, что частота мутаций в общем невелика, и, во-вторых, что разные гены у одного и того же вида часто сильно различаются по мутабильности. Например, у кукурузы ген окраски растения отличается высокой частотой мутаций, тогда как ген восковидно-крахмального эндосперма высоко стабилен. Наконец, в-третьих, частота возникновения мутаций у бактерий ниже, чем у многоклеточных организмов, причём различия колеблются от одного до нескольких порядков величин. Следует указать, что приведённые в таблице данные по нескольким бактериальным генам можно считать репрезентативными, поскольку они выбраны из гораздо большего количества данных. В общем, гены бактерий, по-видимому, более стабильны, чем гены эукариотических организмов.
Имеются основания считать, что по крайней мере некоторые оценки частоты мутаций у высших организмов завышены. Один из источников ошибок — трудность в различении истинных внутригенных мутаций и редких рекомбинаций очень тесно сцепленных генных единиц. Эти два явления могут приводить к одинаковому видимому результату, а именно к резкому фенотипическому изменению, передающемуся по наследству. Любой обширный набор мутаций какого-либо диплоидного организма, наблюдаемый на фенотипическом уровне, по всей вероятности, содержит помимо истинных генных мутаций некоторую долю необнаруженных редких рекомбинаций, что искажает оценку частоты мутаций в сторону повышения .
Другой возможный источник завышенной оценки частоты мутаций — невыявленное селективное преимущество гетерозиготной формы над соответствующими гомозиготами в диплоидных популяциях.
Если даже истинная частота мутаций у высших организмов на порядок ниже, чем показывают современные оценки, то всё же эта частота достаточна, чтобы поддерживать уровень мутационной изменчивости в популяциях. В популяции средней величины, продуцирующей до 100 млн. гамет, на каждый ген в среднем будет возникать по крайней мере по нескольку новых мутаций в каждом поколении.
Генотипический контроль
У Drosophila melanogaster известен ген, который вызывает высокую частоту мутаций других генов данного комплемента. Этот ген-мутатор обозначают символом Hi. У мух, гомозиготных по гену Hi, частота мутаций в 10 раз выше обычной; у гетерозигот по этому гену частота мутаций выше обычной в 2 — 7 раз. Ген Hi индуцирует как видимые, так и летальные мутации во многих генах. Он вызывает также инверсии — один из типов хромосомных мутаций (Ives, 1950; Hinton, Ives, Evans, 1952*).
Выше мы рассматривали генные мутации как случайные ошибки копирования, происходящие во время воспроизведения гена, и это, несомненно, само по себе верно. Однако, как показывают данные о действии генов-мутаторов, существует и другой аспект мутационного процесса. Возникновение новой мутационной изменчивости, которая имеет важное значение для долговременного успеха данного вида в эволюции, может быть не целиком предоставлено воле случая, а инициироваться генамимутаторами. Частота возникновения мутаций у данного вида частично может быть одним из генотипически контролируемых компонентов всей его генетической системы.
Близкородственные виды Drosophila willistoni и D. prosaltans, обитающие в тропических областях Америки, различаются по частоте возникновения мутаций. Представлены частоты летальных мутаций в хромосомах II и III у этих двух видов.
Вид D. willistoni, у которого частота мутаций выше, обычен, широко распространен и занимает разнообразные экологические ниши, тогда как D. prosaltans встречается редко и лишь в строго определённых экологических условиях. Было высказано весьма правдоподобное мнение, что высокая частота возникновения мутаций, поставляющих новые варианты, способствует повышению экологического разнообразия D. willistoni, а тем самым и обилию этого вида (Dobzhansky, Spassky, Spassky, 1952*).
Адаптивная ценностьБольшинство новых мутантов характеризуется более низкой жизнеспособностью, чем нормальный, или дикий, тип. Снижение жизнеспособности может быть выражено в разной степени — от чуть заметного субвитального состояния до полулетальности и летальности. При оценке жизнеспособности мутантов Drosophiia melanogaster, возникших в результате мутаций в Х-хромосоме, 90% оказались менее жизнеспособными, чем нормальные мухи, а 10% были супервитальными, т.е. обладали повышенной жизнеспособностью. Среди 90% мух с пониженной жизнеспособностью наблюдается весь диапазон изменений от слабой субвитальности (45%) через промежуточные стадии понижения жизнеспособности до полулетальности (6%) и летальности (14%) (Тимофеев-Ресовский, 1940*).
Вообще адаптивная ценность новых мутантов обычно бывает понижена. Адаптивная ценность слагается из плодовитости и функциональной полезности морфологических признаков, а также физиологической жизнеспособности. Многие мутанты неплодовиты независимо от того, обладают они нормальной жизнеспособностью или нет.
Морфологические макромутации обычно сопровождаются нарушениями функциональной эффективности. Из обширной выборки индуцированных мутантов ячменя (Hordeum vulgare) у 99% адаптивная ценность была понижена (Gustafsson, 1951*).
Этим наблюдениям легко дать объяснение. Все гены, входящие в состав генотипа нормальной, или дикой, формы на протяжении многих поколений подвергались естественному отбору; они прошли сквозь сито отбора, в результате чего сохранились те из них, которые обладали максимальной адаптивной ценностью. Следует ожидать, что любые изменения в таких генах почти наверное окажутся изменениями к худшему, подобно тому, как копанье наугад в часовом механизме скорее нарушит, чем усовершенствует его «функциональную эффективность».
Генные мутации нередко называют случайными изменениями в генах. В данном контексте определение «случайные» требует разъяснения. Мутационные изменения на самом деле могут не быть случайными на молекулярном уровне. Определенные изменения последовательности нуклеотидов могут возникать в цепи ДНК чаще, чем другие. Так называемая случайность мутационного процесса относится не к молекулярному строению, а к адаптивным свойствам мутантных генов. Мутации случайны в том смысле, что они не направлены в сторону какого-либо нынешнего или будущего состояния приспособленности данного организма.
И всё же небольшая доля всей массы генных мутаций, как это было установлено у генетически хорошо изученных организмов, в том или другом отношении превосходит стандартный тип. Так, в одной выборке мутантов ячменя примерно 0.1—0.2% мутантов обладали повышенной урожайностью в стандартной или в обычной для родительской формы среде (Gustafsson, 1951*).
Мутант, обладающий низкой адаптивной ценностью в стандартных условиях среды, может оказаться адаптивно более ценным в других условиях. Один из мутантов Drosophila funebris —мутант eversae — при 15°С обладает пониженной жизнеспособностью (98% от жизнеспособности дикого типа), но при 24 °С его жизнеспособность выше (104%) (Dobzhansky, 1951*). Шесть мутантов львиного зева (Antirrhinum majus) при нормальных условиях в теплице проявляли более низкие качества, чем родительская линия, но при различных аномальных условиях в теплице они превосходили родительскую линию по росту (Brucher, 1943; Gustafsson, 1951*).
Относительное значение в эволюции малых мутаций и макромутаций
Большинство эволюционных генетиков подчеркивают важную роль в эволюции малых мутаций. Согласно же мнению меньшинства, к которому принадлежат Гольдшмидт (Goldschmidt, 1940; 1952; 1953; 1955*) и некоторые другие ученые, главную роль в эволюции играют макромутации. В прошлом между сторонниками двух указанных противоположных точек зрения было много споров, которые, однако, излишни, поскольку эти взгляды не исключают, а напротив, дополняют друг друга. В эволюции играют роль как малые мутации, так и макромутации.
Как сырье для эволюционных изменений малые мутации обладают некоторыми явными преимуществами. Каждая малая мутация вызывает лишь небольшой фенотипический эффект —к лучшему или к худшему. Поэтому аллель, возникший в результате малой мутации и обладающий слабым преимуществом, может включиться в уже существующий генотип, не вызывая какой-либо резкой дисгармонии. При помощи ряда малых мутаций, происходящих в разных локусах, можно создать тот или иной адаптивно количественный эффект, не нарушая функциональной эффективности организма во время промежуточных стадий этого процесса.
Вероятность сохранения новой мутацииКаждый отдельный мутантный аллель при своем первом возникновении бывает редок. Насколько вероятно, что он сохранится в родительской популяции? Мы знаем, что частота нового мутантного аллеля, нe может увеличиваться за счет одного лишь процесса воспроизведения. Вероятность же чисто случайной элиминации мутантного аллеля, напротив, очень велика.
Фишер (Fisher, 1930; 1958*) вычислил вероятность сохранения нового мутантного аллеля, возникшего у одной особи в обширной популяции Как показывают его данные, шансы на сохранение единичной мутации заметно понижаются с каждым поколением. Если мутантный аллель в селективном отношении нейтрален, то вероятность его исчезновения к 31-му поколению составляет 94%, а к 127-му — 98%. Если он обладает небольшим селективным преимуществом (порядка 1%), то вероятность его исчезновения несколько снижается (93 и 97% в поколениях 31 и 127 соответственно), но остается высокой.
Давление мутацийРоль мутационного процесса в эволюции можно определить следующим образом. Мутации служат основным источником новшеств в популяциях. Однако мутационный процесс обычно не представляет собой важной эволюционной силы. Частота возникновения мутаций столь низка, что само по себе мутационное давление не может вызвать существенных изменений частот генов в популяциях. Кроме того, прямые мутации А→а со временем отсеиваются, полностью или частично, в результате обратных мутаций а→А.
Приведенное выше утверждение касается нормы. В тех особых ситуациях, когда давление мутаций становится значительной силой, вполне могут возникнуть исключения. Во всей Британской Колумбии и на Аляске окраска цветков у Epilobium angustifolium по большей части одинакова; однако в одной популяции, растущей на вершине урансодержащих гор вблизи Большого Медвежьего озера (Канада), обнаружены многочисленные мутантные растения с аберрантными бледно-розовыми цветками (Shacklett, 1964*). Высокая частота хромосомных мутаций наблюдалась у растений, выращенных из семян, которые находились в зоне извержения вулкана в Исландии (Love, 1949*).
Так или иначе мутационное давление независимо от его интенсивности не может быть направляющей силой эволюции. Огромное большинство мутаций, как мы видели, вредны и им суждено быть элиминированными из популяции. Ориентация, или направленность, процесса эволюции определяется другими эволюционными силами.
Эволюционные процессы в популяциях
Эволюционное изменение — это явление, относящееся к популяциям и системам популяций. В своей простейшей форме эволюция осуществляется в пределах локальных воспроизводящихся популяций. Это микроэволюция. Локальную воспроизводящуюся популяцию можно рассматривать как плацдарм для микроэволюции. Необходимо поэтому начать наш обзор процессов микроэволюции с разбора общих свойств популяций.
«Популяция — достаточно многочисленная совокупность особей определённого вида, в течение большого числа поколений населяющая определённое пространство (внутри которого нет установившихся изоляционных барьеров) и отделенная от таких же совокупностей особей данного вида той или иной степенью давления тех или иных форм изоляции. Это определение приложимо только к двуполым скрещивающимся формам. Популяцией у организмов, размножающихся бесполым путем или путем облигатного партеногенеза или самооплодотворения, нужно считать группу особей клона или чистой линии (или смеси клонов и чистых линий), занимающих определённый ареал и отделенную от таких же совокупностей особей пространством с меньшей численностью или отсутствием особей данного вида» (Яблоков, Ларина, 1985). Такое определение соответствует широкому кругу реально существующих популяций.
Целесообразно представлять себе популяции и популяционные системы в виде иерархии — от случайно скрещивающейся группы до вида. Скрещивающаяся популяция — популяционная единица, имеющая некую локальную протяженность в этой непрерывной иерархии.
Структура популяцииСкрещивающаяся популяция представляет собой репродуктивную единицу. У организмов с половым размножением — это сообщество особей, связанных между собой узами скрещивания и взаимоотношениями родители — потомки (Dobzhansky, 1950*). У организмов с бесполым размножением сохраняются связи родители — потомки, но перекрестные связи между особями, обусловленные скрещиванием, сильно ослаблены; не следует, однако, полностью исключать наличие у бесполых организмов таких связей, поскольку у многих организмов, которые обычно размножаются бесполым путем, существуют те или иные парасексуальные способы размножения или же время от времени происходит возврат к половому размножению. Популяция, как правило, представляет собой свободно скрещивающуюся группу, независимо от того, происходит ли скрещивание регулярно или эпизодически, и во всех случаях это некая репродуктивная единица. Популяция представляет собой также экологическую единицу. Составляющие её особи генотипически сходны по своей экологической толерантности, занимают определённую область в той или иной экологической нише или местообитании и предъявляют сходные требования к условиям среды.
Реально существующие популяции очень разнообразны по величине и форме. Структура популяции слагается из четырёх главных компонент: величины популяции, пространственной конфигурации, системы размножения и скорости расселения.
Величина популяции, т. е. число половозрелых размножающихся особей в каждом поколении (N), может колебаться от нескольких единиц до многих миллионов. Что касается пространственного распределения популяции, то можно выделить три основные категории: 1) большие непрерывные популяции; 2) мелкие изолированные колониальные популяции (или популяции, соответствующие островному типу); 3) линейные популяции. Кроме того, существуют разнообразные варианты, промежуточные между этими трёмя основными типами.
Примером больших непрерывных популяций служат популяции злаков, растущих на равнинах и покрывающих площади шириной в десятки или сотни километров. Организмы с колониальной популяционной системой образуют ряд разбросанных, разобщённых и нередко мелких популяций. Примерами могут служить наземные животные, обитающие на архипелагах; пресноводные формы, населяющие цепь озер, обитатели горных вершин в горной местности и организмы, ограниченные определённым типом почвы или горной породы с пятнистым распределением. Линейные популяции возникают вдоль рек, на побережьях морей и в аналогичных местообитаниях, обладающих большой протяженностью и более или менее непрерывных в одном измерении, но коротких и ограниченных в другом.
Часто встречаются и различные промежуточные состояния. Большая популяция может быть непрерывной в одних частях занимаемой ею области, но прерывистой или полунепрерывной в других. Подобным же образом колонии, населяющие систему островов, могут быть изолированы лишь частично, а не полностью. В следующем разделе мы опишем конкретный пример — структуру популяции гигантской секвойи, или мамонтова дерева (Sequoiadendron giganteum), в которой сочетаются самые разнообразные состояния — от изолированных колоний на севере до прерывистого лесного пояса на юге.
Что касается систем размножения, то их диапазон очень широк — от свободного неродственного скрещивания до самооплодотворения. Часто встречаются такие промежуточные типы, как свободное скрещивание между близкими соседями; инбридинг, осуществляемый иными способами, нежели самооплодотворение (например скрещивание между сибсами у животных); сочетание неродственного скрещивания с самоопылением, как у гермафродитных, но самосовместимых цветковых растений.
Варианты пространственного распространения и систем размножения встречаются во всевозможных сочетаниях, создавая в результате чрезвычайно разнообразные структуры популяций. Так, большая непрерывная популяция может состоять из свободно скрещивающихся особей, как у многих опыляемых ветром травянистых растений равнин, но она может также состоять из особей с ограниченной свободой скрещивания или из инбредных особей. Подобно этому небольшая изолированная колония может состоять или из свободно скрещивающихся или из инбредных особей. Структура популяции оказывает влияние на характер её изменчивости, о чем будет сказано ниже.
Популяции Мамонтова дереваМамонтово дерево (Sequoiadendron giganteum) — перекрёстноопыляющееся при помощи ветра хвойное дерево — встречается в сосново-пихтовых лесах на небольших высотах (1500 — 2400 м) на западных склонах гор Сьерра-Невада в Калифорнии. Область его распространения образует узкую полосу протяженностью около 400 км . В пределах этой области мамонтово дерево встречается в виде ряда обособленных и более или менее разобщённых популяций
Число локальных популяций по разным оценкам колеблется от 32 до 75 в зависимости от того, считать ли популяции в форме гантелей за одну или за несколько. По данным разных ботаников, число рощ равно 32 (Jepson, 1909), 71 (Fry, White, 1938) и 75 (Rundel, 1972a)*. Мы следуем оценке Джепсона (Jepson) с тем, чтобы использовать его старые демографические данные.
В самом начале XX в. 32 различаемые здесь локальные популяции имели размеры, приведённые. Мелкие популяции называют рощами, а крупные, за немногими исключениями — лесами. В период проведения учетов, результаты которых представлены, эти леса интенсивно вырубались, но пни от поваленных деревьев ещё оставались на месте. Поэтому данные таблицы отражают величину прежних, ненарушенных популяций. Величина популяций сильно варьирует — от рощиц, состоящих всего из нескольких деревьев, До лесов, в которых насчитываются тысячи индивидуумов. По данным более позднего учета (Fry, White, 1938*), число деревьев в большинстве популяций уменьшилось.
В настоящее время границы локальных популяций определяются, по крайней мере в некоторых исследованных случаях, влажностью почвы. Рощи находятся в тех местах, где в засушливые летние месяцы почва содержит достаточно влаги. Источником почвенной влаги в высоких горах служат летние дожди. Некоторые участки, расположенные на средней высоте, в период дождей накапливают грунтовые воды, а другие нет; мамонтово дерево растет на участках первого типа (Rundel, 1972b*).
ПолиморфизмСогласно определению, полиморфизм — это существование в популяции двух или более резко различающихся (прерывистых) форм, при котором частота более редкой формы определяется не одним лишь мутированием (Ford, 1964, 1965*). Иными словами, полиморфизм — это такая изменчивость в локальной воспроизводящейся популяции, при которой проявляется чётко выраженное или резкое менделевское расщепление.
Такое определение полиморфизма не позволяет относить к нему некоторые типы изменчивости. Оно исключает чисто фенотипическую изменчивость (поскольку это негенетическая изменчивость); оно исключает географическую изменчивость (которой не существует в одной популяции); оно исключает полигенную изменчивость (при которой не происходит расщепления на резко различающиеся классы); и, наконец, оно исключает генетическую изменчивость, обусловленную новыми или повторными мутациями.
На основе разных критериев можно выделять различные типы полиморфизма. Важно различать генетический полиморфизм и хромосомную изменчивость. Генетический полиморфизм — это прерывистая изменчивость по гомологичным аллелям одного и того же генного локуса. Хромосомным называют полиморфизм по типам хромосом, например по половым хромосомам, или по таким перестройкам, как инверсии.
Различают также переходный и сбалансированный полиморфизм. В случае переходного полиморфизма разнообразие носит временный характер: оно наблюдается до тех пор, пока происходит процесс замещения одной формы другой при контролирующем действии естественного отбора. При сбалансированном полиморфизме разные типы представляют собой более или менее постоянные компоненты данной популяции благодаря естественному отбору, который благоприятствует сохранению разнообразия (Ford, 1964, 1965*).
Все формы полиморфизма — генетический, хромосомный, переходный и сбалансированный — весьма обычны и широко распространены в живой природе. В популяциях организмов с половым размножением полиморфизм в сущности наблюдается всегда. В следующем разделе мы опишем конкретный пример — полиморфизм по группам крови у человека.
Полиморфизм по группам крови у человекаРазные люди по-разному реагируют на переливание крови. В некоторых случаях переливание крови приводит к агглютинации, или слипанию, эритроцитов, тогда как в других этого не происходит. Реакция агглютинации обусловлена иммунологическим взаимодействием антигенов, содержащихся в эритроцитах донора, и антител, содержащихся в сыворотке крови реципиента.
В зависимости от типа содержащегося в крови антигена различают четыре группы крови (А, В, АВ и О). Каждый человек относится к одному из этих четырёх фенотипических классов. Индивидуум, обладающий кровью группы А, может дать свою кровь для переливания другому индивидууму с той же группой крови, у которого при этом не возникнет агглютинации. Подобным же образом кровь группы В можно переливать людям, имеющим группу В. Но переливание крови группы А людям с группой В или крови группы В людям с группой А вызывает сильную агглютинацию. Нет нужды описывать здесь реакции, возникающие при всех возможных сочетаниях групп крови (см. Stern, 1960; Race, Sanger, 1962*). В общем агглютинация происходит в тех случаях, когда донор и реципиент имеют разные группы крови.
Группы крови определяются серией из трёх аллелей: IA, IB и I0. Аллель I0 рецессивен по отношению к аллелям IA и IB (иногда его обозначают буквой i). Аллели IA и IB — кодоминантны. Шесть диплоидных генотипов, образуемых этими трёмя аллелями, фенотипически проявляются как четыре группы крови в системе АВО (табл. 3.2). На самом деле у аллеля IA имеются различные, но иммунологически сходные изоаллели (IA1, IA2 и т. д.), поэтому число возможных генотипов больше шести; однако в нашем обсуждении этими тонкими различиями аллеля IA можно пренебречь.
Популяции человека обычно полиморфны по группам крови системы АВО. Частоты различных групп крови и лежащих в их основе аллелей известны для сотен локальных популяций из самых различных областей земного шара. В табл. 3.3 приведены частоты разных аллелей для трёх популяций. Как показывают эти примеры, разные популяции человека сходны в том, что все они полиморфны по группам крови АВО, однако различаются по частотам аллелей. У каждой локальной популяции имеется свой генофонд с характерным для нее составом аллелей гена I.
Локальные популяции представляют собой части более крупных региональных расовых групп. У родственных локальных популяций, обитающих в одной и той же области, генофонды обычно несколько различаются. Так, частота аллеля IA среди населения шведского города Упсала составляет, как указано в табл. 3.3, 31.9%, а среди населения другого шведского города, Фалуна — 28.4%. Напротив, между географическими расами наблюдаются устойчивые различия ко частоте аллелей.
Почти во всех популяциях коренных обитателей Западной Европы наблюдается высокая частота аллеля IA и низкая (менее 10%) частота аллеля IB. В Центральной Азии наблюдается высокая (20 — 30%) частота аллеля IB. Среди американских индейцев аллель I0 встречается с высокой частотой, тогда как аллель IB редок или отсутствует вовсе (Mourant, 1954; Mourant et al., 1976*). Равновесие между разными типами несколько сдвигается при переходе из одной географической области в другуюЧрезвычайно интересно, что параллельная полиморфная изменчивость по группам крови АВО обнаружена у человекообразных обезьян. У шимпанзе найдены группы А и 0, у орангутана и гиббона имеются группы А, В и АВ (Mourant, 1954; Wiener, Moor-Jankowsky, 1971*). Таким образом, полиморфизм по группам крови системы АВО возник в процессе эволюции раньше, чем сам вид Homo sapiens, и им обладают также ближайшие родичи человека в отряде приматов.
У человека существует ещё несколько систем групп крови: система Rh, система MN и другие. Популяции человека полиморфны также и по этим системам (Race, Sanger, 1962; Mourant et al., 1976*). Полиморфная изменчивость по системам Rh и другим, по-видимому, независима от изменчивости по системе АВ0.
Полиморфизм по ферментамМетод гель-электрофореза даёт возможность обнаруживать полиморфизм по ферментам и по некоторым белкам, который не удавалось выявить обычными генетическими методами. Экстракт какой-либо ткани помещают в гель и подвергают действию электрического поля. Вследствие характерных особенностей подвижности в электрическом поле разных ферментов последние физически разделяются; затем гель окрашивают, и разные ферменты выявляются в виде обособленных пятен. Этим методом можно выявить аллельные различия для одной ферментной системы и генные различия между ферментами.
Гетерозиготность и нормаГенофонды свободно скрещивающихся животных и растений обычно содержат, как было отмечено выше, во многих генных локусах по многу аллелей. Таким образом, в результате свободного скрещивания возникают высокогетерозиготные диплоидные особи. Многие аллели в каждом локусе уже подвергались в предыдущих поколениях отбору на хорошую комбинационную способность в диплоидных гетерозиготных генотипах. Особи с нормальным фенотипом и нормальной жизнеспособностью представляют собой гетерозиготы. Кроме того, нормальное фенотипическое состояние создаётся различными гетерозиготными комбинациями генов. Об этом свидетельствуют результаты инбридинга таких свободно скрещивающихся диплоидных организмов.
В популяциях Drosophila melanogaster встречаются иногда аберрантные особи с дополнительными жилками на крыльях; частота таких особей очень низка. Дубинин (1948)* использовал большое число нормальных самок дикого типа из алма-атинской популяции D. melanogaster в качестве исходного материала для выведения чистых линий. Оказалось, что 60 различных инбредных линий давали потомков с дополнительными жилками. После инбридинга на протяжении ряда поколений этот аберрантный признак был выявлен в 68% изученных линий. Этот признак экспрессировался в результате инбридинга и в других популяциях, помимо алма-атинской.
Концепция популяцииКонцепцию генетически изменчивых популяций как репродуктивных единиц ни в коей мере нельзя считать самоочевидной. Этой концепции не существовало ни в XVIII, ни в начале XIX в., а в некоторых областях биологии она отсутствует до сих пор. Согласно Майру (Mayr, 1972a, 1982*), её ввел в биологию Дарвин в 1859 г. Концепция популяции была одним из элементов переворота, произведенного Дарвином в научном мышлении.
Концепция популяции прямо противоположна эссенциализму. Сторонники эссенциализма считают, что наблюдаемые в мире явления представляют собой отражения лежащих в их основе сущностей. Явления предстают в различных формах, сущность же их неизменна. Следовательно, члены того или иного класса объектов, в том числе и особи, составляющие какую-либо популяцию, — это различные выражения одной и той же сущности. Эссенциализм в том или ином варианте был традиционным философским учением в Европе. Философия Платона, христианское богословие и философский идеализм представляли собой различные модификации эссенциализма. Естественно, что эссенциализм господствовал в научном мышлении в ранний период развития биологии. Здесь он принял форму, которую Майр (Mayr, 1957a, b, 1972a; 1982*) назвал типологическим мышлением. Согласно этим взглядам, отдельные организмы представляют собой несовершенные, а потому изменчивые проявления архетипа того вида, к которому они принадлежат.
Борьба за существование. Геометрическая прогрессия размножения и борьба за существование
Вы, наверное, слышали эту старинную задачу. "Индийский царь предложил изобретателю шахмат, чтобы он сам выбрал себе награду за создание этой игры. Тот попросил выдать ему за первую клетку шахматной доски одно пшеничное зерно, за вторую - два, за третью - еще в два раза больше и т.д. Сколько зерен должен получить изобретатель шахмат?"
Сформулируем эту задачу по-другому. Пусть у нас есть одно зерно пшеницы. Посадим его в землю. Из него вырастет растение, в колосе которого будет всего 2 зерна. Высадим их вновь и так далее. Сколько растений у нас будет через 64 поколения? Ответ: 9 223 372 036 854 775 808 растений. Если каждое растение занимает площадь 1 кв. см., то общая площадь нашего пшеничного поля через 64 поколения почти вдвое превысит площадь Земного шара.
Давно известно, что все живые организмы размножаются в геометрической прогрессии. Как писал Дарвин: «Нет ни одного исключения из правила, по которому любое органическое существо численно возрастает естественным путем с такой большой скоростью, что не подвергайся оно истреблению, потомство одной пары очень скоро заняло бы всю землю. Считается, что из всех известных животных наименьшая воспроизводительная способность у слона, и я старался вычислить вероятную минимальную скорость естественного возрастания его численности; он начинает плодиться, всего вероятнее, в 13-летнем возрасте и плодится до 90 лет, принося за это время не более шести детенышей, а живет до ста лет; если это так, то по истечении 740—750 лет от одной пары получилось бы около 19 миллионов живых слонов».
Давно известно и то, что в каждом поколении огромное количество живых организмов гибнет на разных стадиях своего жизненного цикла. Из множества рожденных потомков только немногие доживают до вступления в цикл размножения. Не все дожившие участвуют в размножении. При оплодотворении каждая зигота получает только крохотный шанс на жизнь. За саму жизнь приходится бороться. Но этого мало. Мало просто выжить и благополучно состариться. Для того, чтобы оставить свой след в эволюции, нужно произвести потомство. Но и этого мало. Мало просто произвести потомство, нужно это потомство сохранить. За это тоже надо бороться.
Еще Дарвин предупреждал, что борьбу за существование не следует понимать, как примитивную драку. Он писал: «Я должен предупредить, что применяю этот термин в широком и метафорическом смысле, включая сюда зависимость одного существа от другого, а также включая (что еще важнее) не только жизнь особи, но и успех в оставлении потомства».
Принято выделять три формы борьбы за существование: борьбу за жизнь с абиотическими факторами (конституциональную борьбу), с представителями других видов (межвидовую борьбу) и с представителями своего собственного вида (внутривидовую борьбу). Это выделение весьма условно и, по существу, как мы увидим далее, эти формы тесно связаны друг с другом.
Конституциональная борьба за существование
Жизнь на Земле заполнила множество экологических ниш, которые очень сильно отличаются друг от друга. Даже организмы, принадлежащие к одному отряду, могут обитать в очень разных условиях. Например, грызуны обитают и далеко за Полярным кругом, и в раскаленных пустынях. Каждую из этих ниш занимает своя группа видов, уже прошедших через многие поколения борьбы за существования в этих условиях.
Эта борьба не прекращается ни на минуту. В каждом поколении рождаются особи, которые генетически отличаются друг от друга. Среди них выживают только те, кто наилучшим образом приспособлен к тем условиям, которые есть «здесь» и «сейчас» - в данной экологической нише и в данный момент времени. Но внешние условия непостоянны. Температура и влажность резко меняются в течение каждого года. В таких условиях преимущество в борьбе за жизнь получают особи, которые способны адаптироваться ко всему спектру этих изменений. Борьба за жизнь происходит каждую минуту, но победителями в этой борьбе оказывается те, кто продержится дольше, те, кто обладает наиболее надежными и хорошо скоординированными адаптациями.
Внешние условия непостоянны не только в течение года, они постепенно меняются год за годом. Меняется климат на всей Земле. Приходят и уходят ледниковые периоды. Те приспособления, которые обеспечивали победу в борьбе за жизнь вчера, могут оказаться неадекватными завтра. Физический мир вокруг живых организмов постоянно и непредсказуемо меняется. Вместе с ним меняются и правила борьбы за жизнь, меняются критерии приспособленности.
Межвидовая борьба за существование
Жизнь каждого организма зависит не только от абиотических условий, но и от множества других видов животных, растений, микроорганизмов, с которыми он, так или иначе, взаимодействует. Взаимоотношения типа хищник-жертва, паразит-хозяин играют важнейшую роль в жизни каждого организма.
Каждое усовершенствование любого вида в экосистеме ведет к ухудшению условий для других видов. Поэтому, для того, чтобы выжить, все виды, входящие в экосистему должны непрерывно эволюционировать. Эта закономерность получила название «принцип Красной Королевы» по имени героини книги Л. Кэрролла «Алиса в Зазеркалье». Крылатая фраза Красной Королевы «В этом мире нужно бежать из всех сил, только для того, чтобы остаться на месте» отражает самое существо борьбы за существование.
Большинство живых организмов погибает или оказывается исключенными из размножения не под действием физических факторов, а в результате действий других видов – паразитов, хищников, конкурентов. В межвидовой борьбе, как и в борьбе с абиотическими факторами, не бывает передышек. Она идет постоянно, день за днем, поколение за поколением и правила этой борьбы постоянно меняются. Однако эти изменения носят совсем иной характер, чем в борьбе с абиотическими факторами. Климат меняется постоянно и непредсказуемо, но он меняется не обязательно во вред живым организмам. Хищник всегда меняется во вред жертве, совершенствуя свои методы охоты. Жертва меняется во вред хищнику, совершенствуя способы защиты от него. Паразит меняется таким образом, чтобы использовать максимум ресурсов, которые он может черпать из организма хозяина, а хозяин развивает и усовершенствует все новые и новые средства борьбы с паразитом. Возникает ситуация замкнутого круга, когда усовершенствование жертвы в противостоянии хищнику влечет за собой усовершенствование способов охоты у хищников, которое в свою очередь влечет за собой усовершенствование жертвы, и так круг за кругом. Всем видам, входящим в экосистему приходится «бежать из всех сил, только для того, чтобы остаться на месте» - приходится постоянно меняться только для того, чтобы сохранить своем место в экосистеме.
Парадоксальная особенность межвидовой борьбы за существование состоит в том, что в ней бывают побежденные, но не бывает безусловных победителей. Если один из видов, включенных в экосистему, «проигрывает» в этой борьбе и вымирает, то жизнь видов-победителей от этого не становится легче. Если вид-хищник истребляет вид-жертву, он тем самым ставит под угрозу свое собственное существование. Если жертве удается «победить» хищника, став трудно достижимой для него добычей, то это оказывается Пирровой победой. За ней следует увеличение численности вида «победителя», быстрое истощение необходимых ему ресурсов, резкое увеличение численности паразитов и в результате сам «победитель» оказывается на грани вымирания. Вымирание тех или иных видов не снижает остроты межвидовой борьбы в биоценозе – на смену вымершим видам приходят их бывшие конкуренты.
Особой остроты межвидовая борьба достигает в тех случаях, когда противоборствуют разные виды, обитающие в сходных экологических условиях и использующие одинаковые источники питания. В результате межвидовой конкуренции происходит либо вытеснение одного из видов, либо приспособление видов к разным условиям в пределах единого ареала, либо, наконец, их территориальное разобщение.
Внутривидовая борьба за существование. Представьте себе ситуацию. Два зайца удирают от волка. Один из них говорит другому: «На что ты надеешься? Тебе все равно не удастся бежать быстрее волка». «А мне не нужно бежать быстрее волка, мне нужно бежать быстрее, чем ты: тогда волку достанешься ты, а не я, - отвечает другой. Зайцы соревнуются в скорости бега не с волками, а друг с другом. Волк, который гонится за зайцем, соревнуются не с ним, а с другим волком, который в другом лесу гонится за другим зайцем. Именно на внутривидовом уровне реализуется и межвидовая борьба, и борьба с абиотическими факторами.
Особи, принадлежащие к одному виду, отличаются друг от друга по множеству признаков. Среди множества особей данного вида выживают и размножаются только те, которые лучше, чем их соплеменники противостоят превратностям климата, спасаются от хищников, добывают пищу, защищаются от паразитов. Таким образом, борьба с абиотическими факторами и межвидовая борьба являются компонентами внутривидовой конкуренции. Однако конкуренция между особями одного вида этим не ограничивается.
Тенденция каждого вида к неограниченному размножению вступает в противоречие с ограниченностью жизненных ресурсов: пищи, воды, укрытий, солнечного света и т.п. За все эти ресурсы возникает жесткая конкуренция между особями одного вида, поскольку все они имеют одинаковые потребности. Эта конкуренция может быть прямой и легко заметной для наблюдателя. Так, например, борьба за подходящие укрытия, за наиболее богатые кормовые участки, за самок может приводить к открытым конфликтам между представителями одного и того же вида. Однако чаще всего внутривидовая конкуренция оказывается непрямой и скрытой от наблюдения. Быстро растущие деревья затеняют медленно растущие и тем самым лишают их солнечного света – важнейшего и, казалось бы, неограниченного ресурса. Лев, съедая антилопу, тем самым отнимает пищу у другого льва. В дупле, уже занятом одной парой птиц, не может поселиться другая пара. Победители во внутривидовой борьбе, таким образом, получают преимущество в размножении и, соответственно, растет их генетический вклад в следующее поколение.
Мы редко наблюдаем в природе прямые конфликты между животными. В то же время многие натуралисты отмечают распространенность внутривидовой взаимопомощи, кооперации и даже самопожертвования в мире животных. Муравьи, пчелы, термиты совместно и согласованно возводят общественные сооружения. Птица ставит свою жизнь под угрозу, отвлекая хищника от гнезда с птенцами. Пчела самоубийственно атакует врага, который приближается к улью. Эти наблюдения иногда трактуют как свидетельства против дарвиновского тезиса о борьбе за существование. На самом деле внутривидовая взаимопомощь является весьма эффективным способом борьбы за существование, если мы, следуя Дарвину, понимаем ее «в широком и метафорическом смысле, включая не только жизнь особи, но и успех в оставлении потомства». Если мы внимательно проанализируем примеры «самопожертвования», мы обнаружим, что животные, как правило, жертвуют собой ради своих потомков или других близких родственников. Во внутривидовой кооперации почти всегда участвуют представители близко родственных групп. В результате, доля потомков и родственников особей, проявляющих взаимопомощь, оказываются более значительной в составе следующего поколения, чем доля потомков и родственников особей, не вступающих в кооперацию. Из поколения в поколение увеличивается генетический вклад особей, склонных к взаимопомощи.
Чем больше родство особей вовлеченных в кооперацию, кооперация тем более полное и согласованное взаимодействие особей мы наблюдаем. Наиболее яркие примеры кооперации и взаимопомощи наблюдаются в сообществах социальных насекомых (пчел, муравьев, термитов). Во всех этих случаях каждое сообщество состоит из ближайших родственников. Все население каждого улья, муравейника, термитника – это, как правило, дети одной единственной самки.
Таким образом, борьба за жизнь является следствием противоречия между тенденцией всех живых организмов к неограниченному размножению и ограниченностью жизненных ресурсов. Она может принимать разные формы, но результат ее всегда один – из множества рожденных особей каждого вида выживают и размножаются только те, которые лучше других приспособлены к тем условиям, которые существуют в популяции в каждый момент времени.
Естественный отбор, адаптация - результат действия отбора
Естественный отбор — природный процесс, при котором из всех живых организмов сохраняются во времени только те, которые обладают качествами, способствующими успешному воспроизведению себе подобных. По представлениям СТЭ, естественный отбор является одним из важнейших факторов эволюции.
Механизм естественного отбораИдею о том, что в живой природе действует механизм, подобный искусственному отбору, впервые высказали английские учёные Чарльз Дарвин и Альфред Уоллес. Суть их идеи состоит в том, что для появления удачных созданий, природе вовсе не обязательно понимать и анализировать ситуацию, а можно действовать наугад. Достаточно создавать широкий спектр разнообразных особей — и, в конечном счёте, выживут наиболее приспособленные.
1. Сначала появляется особь с новыми, совершенно случайными, свойствами
2. Потом она оказывается или не оказывается способной оставить потомство, в зависимости от этих свойств
3. Наконец, если исход предыдущего этапа оказывается положительным, то она оставляет потомство и её потомки наследуют новоприобретённые свойства
В настоящее время, отчасти наивные взгляды самого Дарвина оказались частично переработаны. Так, Дарвин представлял, что изменения должны происходить очень плавно, а спектр изменчивости является непрерывным. Сегодня, однако, механизмы естественного отбора объясняются при помощи генетики, которая вносит некоторое своеобразие в эту картину. Мутации в генах, которые работают на первом этапе описанного выше процесса, являются существенно дискретными. Ясно, однако, что основная суть идеи Дарвина осталась без изменений.
Формы естественного отбора Движущий отборДвижущий отбор — форма естественного отбора, когда условия среды способствуют определённому направлению изменения какого-либо признака или группы признаков. При этом иные возможности изменения признака подвергаются отрицательному отбору. В результате в популяции от поколения к поколению происходит сдвиг средней величины признака в определённом направлении. При этом давление движущего отбора должно отвечать приспособительным возможностям популяции и скорости мутационных изменений (в ином случае давление среды может привести к вымиранию).
Современным случаем движущего отбора является «индустриальный меланизм английских бабочек». «Индустриальный меланизм» представляет собой резкое повышение доли меланистических (имеющих тёмную окраску) особей в тех популяциях бабочек, которые обитают в промышленных районах. Из-за промышленного воздействия стволы деревьев значительно потемнели, а также погибли светлые лишайники, из-за чего светлые бабочки стали лучше видны для птиц, а тёмные — хуже. В XX веке в ряде районов доля тёмноокрашенных бабочек достигла 95 %, в то время как впервые тёмная бабочка (Morfa carbonaria) была отловлена в 1848 году.
Движущий отбор осуществляется при изменении окружающей среды или приспособлении к новым условиям при расширении ареала. Он сохраняет наследственные изменения в определенном направлении, перемещая соответственно и норму реакции. Например, при освоении почвы, как среды обитания у различных неродственных групп животных конечности превратились в роющие.
Стабилизирующий отбор
Стабилизирующий отбор — форма естественного отбора, при котором действие направлено против особей, имеющих крайние отклонения от средней нормы, в пользу особей со средней выраженностью признака.
Описано множество примеров действия стабилизующего отбора в природе. Например, на первый взгляд кажется, что наибольший вклад в генофонд следующего поколения должны вносить особи с максимальной плодовитостью. Однако наблюдения над природными популяциями птиц и млекопитающих показывают, что это не так. Чем больше птенцов или детенышей в гнезде, тем труднее их выкормить, тем каждый из них меньше и слабее. В результате наиболее приспособленными оказываются особи со средней плодовитостью.
Отбор в пользу средних значений был обнаружен по множеству признаков. У млекопитающих новорожденные с очень низким и очень высоким весом чаще погибают при рождении или в первые недели жизни, чем новорожденные со средним весом. Учет размера крыльев у птиц, погибших после бури, показал, что большинство из них имели слишком маленькие или слишком большие крылья. И в этом случае наиболее приспособленными оказались средние особи.
Дизруптивный отбор
Дизруптивный (разрывающий) отбор — форма естественного отбора, при котором условия благоприятствуют двум или нескольким крайним вариантам (направлениям) изменчивости, но не благоприятствуют промежуточному, среднему состоянию признака. В результате может появиться несколько новых форм из одной исходной. Дизруптивный отбор способствует возникновению и поддержанию полиморфизма популяций, а в некоторых случаях может служить причиной видообразования.
Одна из возможных в природе ситуаций, в которой, вступает в действие дизруптивный отбор, — когда полиморфная популяция занимает неоднородное местообитание. При этом разные формы приспосабливаются к различным экологическим нишам или субнишам.
Примером дизруптивного отбора является образование двух рас у погремка лугового на сенокосных лугах. В нормальных условиях сроки цветения и созревания семян у этого растения покрывают всё лето. Но на сенокосных лугах семена дают преимущественно те растения, которые успевают отцвести и созреть либо до периода покоса, либо цветут в конце лета, после покоса. В результате образуются две расы погремка — ранне- и позднецветущая.
Дизруптивный отбор осуществлялся искусственно в экспериментах с дрозофилами. Отбор проводился по числу щетинок, оставлялись лишь особи с малым и большим количеством щетинок. В результате примерно с 30-го поколения две линии разошлись очень сильно, несмотря на то, что мухи продолжали скрещиваться между собой, осуществляя обмен генами. В ряде других экспериментов (с растениями) интенсивное скрещивание препятствовало эффективному действию дизруптивного отбора.
Отсекающий отбор
Отсекающий отбор — форма естественного отбора. Его действие противоположно положительному отбору. Отсекающий отбор выбраковывает из популяции подавляющее большинство особей, несущих признаки, резко снижающие жизнеспособность при данных условиях среды. С помощью отсекающего отбора из популяции удаляются сильно вредные аллели. Также отсекающему отбору могут подвергаться особи с хромосомными перестройками и набором хромосом, резко нарушающими нормальную работу генетического аппарата.
Положительный отбор
Положительный отбор — форма естественного отбора. Его действие противоположно отсекающему отбору. Положительный отбор увеличивает в популяции число особей, обладающих полезными признаками, повышающими жизнеспособность вида в целом. С помощью положительного отбора и отсекающего отбора совершается изменение видов (а не только посредством уничтожения ненужных особей, тогда любое развитие должно остановиться, но этого не происходит).
Среди примеров положительного отбора: чучело археоптерикса можно использовать как планер, а чучело ласточки или чайки нельзя. Но первые птицы летали лучше археоптерикса.
Другой пример положительного отбора — появление хищников, превосходящих своими «умственными способностями» многих других теплокровных.
Или появление таких рептилий, как крокодилы, обладающих четырехкамерным сердцем и способных жить как на земле, так и в воде.
Палеонтолог Иван Ефремов утверждал, что человек прошел не только отбор на лучшую приспособленность к условиям окружающей среды, но и «отбор на социальность» — выживали те сообщества, члены которых лучше поддерживали друг друга. Это еще один из примеров положительного отбора.
Частные направления естественного отбора· Выживание наиболее приспособленных к условиям обитания видов и популяций, например видов, обладающих жабрами в воде, поскольку приспособленность позволяет выигрывать борьбу за выживание.
· Выживание физически здоровых организмов.
· Выживание физически сильнейших организмов, поскольку физическая борьба за ресурсы является неотъемлемой частью жизни. Имеет значение во внутривидовой борьбе.
· Выживание наиболее сексуально успешных организмов, поскольку половое размножение является доминирующим способом размножения. В данном случае в дело вступает половой отбор.
Однако все эти случаи являются частными, а главным остаётся успешное сохранение во времени. Поэтому иногда эти направления нарушаются ради следования главной цели.
Роль естественного отбора в эволюцииЧ. Дарвин полагал естественный отбор основополагающим фактором эволюции живого (селекционизм в биологии). Накопление в конце XIX - начале XX века сведений по генетике, в частности обнаружение дискретного характера наследования фенотипических признаков, подтолкнуло многих исследователей к пересмотру указанного тезиса Дарвина: в качестве чрезвычайно важных факторов эволюции стали рассматриваться мутации генотипа (мутационизм Г. де Фриза, сальтационизм Р. Гольдшмитда и др.). С другой стороны, открытие известных корреляций среди признаков родственных видов (закон гомологических рядов) Н. И. Вавилова привело к формулировке гипотез об эволюции на основе закономерностей, а не случайной изменчивости (номогенез Л. С. Берга, батмогенез Э. Д. Копа и др.). В 1920-1940-е г. г. в эволюционной биологии интерес к селекционистским теориям возродился благодаря синтезу классической генетики и теории естественного отбора.
Разработанная в результате этого синтетическая теория эволюции (СТЭ), часто называемая неодарвинизмом, опирается на количественный анализ частоты аллелей в популяциях, изменяющейся под влиянием естественного отбора.
Тем не менее, открытия последних десятилетий в различных областях научного знания — от молекулярной биологии с её теорией нейтральных мутаций М. Кимуры и палеонтологии с её теорией прерывистого равновесия С. Дж. Гоулда и Н. Элдриджа (в которой вид понимается как относительно статическая фаза эволюционного процесса) до математики с её теорией бифуркаций и фазовых переходов — свидетельствуют о недостаточности классической СТЭ для адекватного описания всех аспектов биологической эволюции. Дискуссия о роли различных факторов в эволюции продолжается и сегодня, и эволюционная биология подошла к необходимости своего очередного, третьего синтеза.
Возникновение адаптаций в результате естественного отбора
Адаптациями называются свойства и признаки организмов, которые обеспечивают приспособление к той среде, в которой эти организмы живут. Адаптацией также называют процесс возникновения приспособлений. Выше мы рассмотрели, как некоторые адаптации возникают в результате естественного отбора. Популяции березовой пяденицы приспособились к изменившимся внешним условиям благодаря накоплению мутаций темной окраски. В популяциях человека, населяющих малярийные районы, адаптация возникла благодаря распространению мутации серповидно-клеточной анемии. И в том, и в другом случае, адаптация достигается за счет действия естественного отбора.
При этом материалом для отбора служит наследственная изменчивость, накопленная в популяциях. Поскольку разные популяции отличаются друг от друга по набору накопленных мутаций, то к одним и тем же факторам внешней среды они приспосабливаются по-разному. Так, африканские популяции адаптировались к жизни в малярийных районах за счет накопления мутаций серповидно-клеточной анемии HbS, а в популяциях, населяющих юго-восточную Азию устойчивость к малярии сформировалась на основе накопления ряда других мутаций, которые в гомозиготном состоянии также вызывают болезни крови, а в гетерозиготном - обеспечивают защиту от малярии.
Эти примеры иллюстрируют роль естественного отбора в формировании адаптаций. Нужно, однако, ясно понимать, что это – частные случаи относительно простых адаптаций, возникающих за счет селективного размножения носителей единичных «полезных» мутаций. Маловероятно, что большинство адаптаций возникло таким путем.
Покровительственная, предостерегающая и подражательная окраска. Рассмотрим, например, такие широко распространенные адаптации, как покровительственная, предостерегающая и подражательная окраска (мимикрия). Покровительственная окраска позволяет животным становится незаметными, сливаясь с субстратом. Одни насекомые поразительно сходны с листьями деревьев, на которых они обитают, другие напоминают засохшие веточки или шипы на стволах деревьев. Эти морфологические адаптации дополняются поведенческими приспособлениями. Насекомые выбирают для укрытия именно те места, где они менее заметны.
Несъедобные насекомые и ядовитые животные - змеи и лягушки, имеют яркую, предостерегающую окраску. Хищник, раз столкнувшись с таким животным, надолго ассоциирует этот тип окраски с опасностью. Этим пользуются некоторые неядовитые животные. Они приобретают поразительное сходство с ядовитыми, и тем самым снижают опасность со стороны хищников. Уж имитирует окраску гадюки, муха подражает пчеле. Это явление называется мимикрией.
Как возникли все эти удивительные приспособления? Маловероятно, чтобы единичная мутация могла обеспечивать такое точное соответствие между крылом насекомого и живым листом, между мухой и пчелой. Невероятно, чтобы единственная мутация заставляла покровительственно окрашенное насекомое прятаться именно на тех листьях, на которые оно похоже. Очевидно, что такие приспособления как покровительственная и предостерегающая окраски и мимикрия возникали путем постепенного отбора всех тех мелких уклонений в форме тела, в распределении определенных пигментов, во врожденном поведении, которые существовали в популяциях предков этих животных. Одной из важнейших характеристик естественного отбора является его кумулятивность – его способность накапливать и усиливать эти уклонения в ряду поколений, слагая изменения отдельных генов и контролируемых ими систем организмов.
Самая интересная и трудная проблема – это начальные стадии возникновения адаптаций. Понятно, какие преимущества дает почти идеальное сходство богомола с сухим сучком. Но какие преимущества могли быть у его далекого предка, который лишь отдаленно напоминал сучок? Неужели хищники так глупы, что их можно так легко обмануть? Нет, хищники отнюдь не глупы, и естественные отбор из поколения в поколение «учит» их все лучше и лучше распознавать уловки их добычи. Даже идеальное сходство современного богомола с сучком не дает ему 100-процентной гарантии, что ни одна птица его никогда не заметит. Однако его шансы ускользнуть от хищника выше, чем у насекомого с менее совершенной покровительственной окраски. Точно также, у его далекого предка, лишь чуть-чуть похожего на сучок, шансы на жизнь были несколько выше, чем у его родственника вовсе на сучок не похожего. Конечно, птица, которая сидит рядом с ним, в ясный день легко его заметит. Но если день туманный, если птица не сидит рядом, а пролетает мимо и решает не тратить времени на то, что может быть богомолом, а может быть и сучком, тогда и минимальное сходство сохраняет жизнь носителю этого едва заметного сходства. Его потомки, которые унаследуют это минимальное сходство, будут более многочисленны. Их доля в популяции станет больше. Это осложнит жизнь птицам. Среди них станут более успешными те, кто точнее будет распознавать замаскированную добычу. Вступает в действие тот самый принцип Красной Королевы, который мы обсуждали в параграфе, посвященном борьбе за существование. Чтобы сохранить преимущество в борьбе за жизнь, достигнутое за счет минимального сходства, виду-жертве приходится меняться.
Естественный отбор подхватывает все те мельчайшие изменения, которые усиливают сходство в окраске и форме с субстратом, сходство между съедобным видом и тем несъедобным видом, которому он подражает. Следует учитывать, что разные виды хищников пользуются разными методами поиска добычи. Одни обращают внимание на форму, другие на окраску, одни обладают цветным зрением, другие нет. Поэтому естественный отбор автоматически усиливает, насколько это возможно, сходство между имитатором и моделью и приводит к тем изумительным адаптациям, которые мы наблюдаем в живой природе.
Возникновение сложных адаптаций. Многие адаптации производят впечатление тщательно продуманных и направленно спланированных устройств. Как такая сложнейшая структура как глаз человека могла возникнуть путем естественного отбора случайно возникавших мутаций?
Ученые предполагают, что эволюция глаза началась с небольших групп светочувствительных клеток на поверхности тела наших очень далеких предков, живших около 550 млн. лет назад. Способность различать свет и тьму была, безусловно, полезна для них, повышала их шансы на жизнь по сравнению с их абсолютно слепыми сородичами. Случайно возникшее искривление «зрительной» поверхности улучшило зрение, это позволяло определить направление на источник света. Возник глазной бокал. Вновь возникающие мутации могли вести к сужению и расширению отверстия глазного бокала. Сужение постепенно улучшало зрение – свет стал проходить через узкую диафрагму. Как видите, каждый шаг повышал приспособленность тех особей, которые менялись в «правильном» направлении. Светочувствительные клетки формировали сетчатку. Со временем в передней части глазного яблока сформировался хрусталик, выполняющий функцию линзы. Он возник, по-видимому, как прозрачная двухслойная структура, наполненная жидкостью.
Ученые попытались смоделировать этот процесс на компьютере. Они показали, что глаз, подобный сложному глазу моллюска, мог возникнуть из слоя фоточувствительных клеток при относительно мягком отборе всего за 364 000 поколений. Иными словами, животные, у которых смена поколений происходит каждый год, могли сформировать полностью развитый и оптически совершенный глаз в менее, чем за полмиллиона лет. Эта очень короткий срок для эволюции, если учесть что средний возраст вида у моллюсков равняется нескольким миллионам лет.
Все предполагаемые стадии эволюции глаза человека мы можем обнаружить среди ныне живущих животных. Эволюция глаза шла разными путями в разных типах животных. Благодаря естественному отбору независимо возникло множество разных форм глаза, и человеческий глаз – только один из них, причем не самый совершенный
Если мы внимательно рассмотрим конструкцию глаза человека и других позвоночных животных, мы обнаружим целый ряд странных несообразностей. Когда свет попадает в глаз человека он проходит через хрусталик и попадает на светочувствительные клетки сетчатки. Свет вынужден пробиваться через густую сеть капилляров и нейронов, чтобы попасть на фоторецепторный слой. Как это ни удивительно, но нервные окончания подходят к светочувствительным клеткам не сзади, а спереди! Более того, нервные окончания собираются в оптический нерв, который отходит от центра сетчатки, и создает тем самым слепое пятно. Чтобы компенсировать затенение фоторецепторов нейронами и капиллярами и избавится от слепого пятна, наш глаз постоянно движется, посылая в мозг серию разных проекций одного и того же изображения. Наш мозг производит сложнейшие операции, складывая эти изображения, вычитая тени, и вычисляя реальную картину. Всех этих сложностей можно было бы избежать, если бы нервные окончания подходили к нейронам не спереди, а сзади как, например, у осьминога.
Само несовершенство глаза позвоночных проливает свет на механизмы эволюции путем естественного отбора. Мы уже не раз говорили о том, что отбор всегда действует «здесь и сейчас». Он сортирует разные варианты уже существующих структур, выбирая и слагая вместе лучшие из них: лучшие «здесь и сейчас», безотносительно к тому, во что эти структуры могут превратиться в далеком будущем. Поэтому ключ к объяснению и совершенств и несовершенств современных структур следует искать в прошлом. Ученые считают, что все современные позвоночные произошли от животных подобных ланцетнику. У ланцетника светочувствительные нейроны располагаются на переднем конце нервной трубки. Перед ними расположены нервные и пигментные клетки, которые прикрывают фоторецепторы от света попадающего спереди. Ланцетник принимает световые сигналы, приходящие с боков его прозрачного тела. Можно думать, что у общего предка позвоночных глаз был устроен сходным образом. Затем эта плоская структура стала преобразовываться в глазной бокал. Передняя часть нервной трубки впячивалась внутрь, и нейроны, находившиеся впереди рецепторных клеток, оказались поверх них. Процесс развития глаза у эмбрионов современных позвоночных в известном смысле воспроизводит последовательность событий, происходивших в далеком прошлом.
Эволюция не создает новых конструкций «с чистого листа», она меняет (часто неузнаваемо меняет) старые конструкции, так чтобы каждый этап этих изменений был приспособительным. Любое изменение должно повышать приспособленность его носителей или, хотя бы, не снижать ее. Эта особенность эволюции ведет к неуклонному совершенствованию различных структур. Она же является причиной несовершенства многих адаптаций, странных несообразностей в строении живых организмов.
Следует помнить, однако, что все приспособления, сколь бы совершенны они ни были, носят относительный характер. Понятно, что развитие способности к полету не очень хорошо совмещается с возможностью быстро бегать. Поэтому птицы, обладающие наилучшими способностями к полету, — плохие бегуны. Напротив, страусы, которые не способны летать, прекрасно бегают. Приспособление к определенным условиям может быть бесполезно или даже вредно при появлении новых условий. Однако условия обитания меняются постоянно и иногда очень резко. В этих случаях накопленные ранее адаптации могут затруднить формирование новых, что может вести к вымиранию больших групп организмов, как это случилось более 60—70 млн лет назад с некогда очень многочисленными и разнообразными динозаврами.
Похожие работы
... виде, или постепенно, в течение «дней» «производить» из воды и земли, от низших форм к высшим силою заложенных Им в природу законов». 3. Критика эволюционного учения и креационизма Теория эволюции подвергается креационистами критике, в основном, по следующим направлениям. 1. Палеонтологическая летопись обнаруживает скорее структуру эволюционных скачков, а не постепенные преобразования. ...
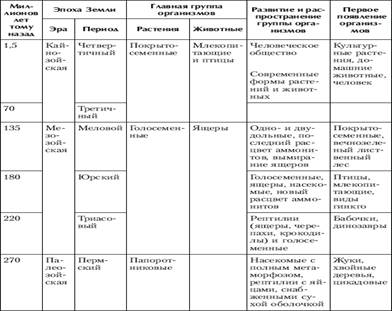
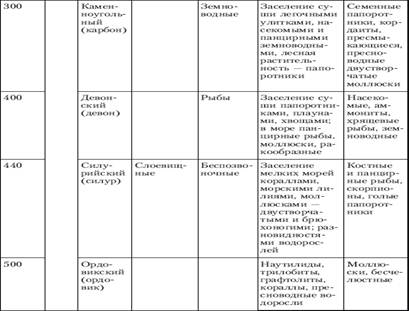
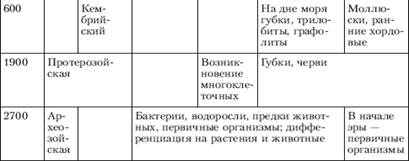
... отношений, появление более совершенных орудий труда, жилища, одежды привели к снижению роли биологического фактора эволюции и усилению ведущего действия социальных законов развития[5]. Заключение Эволюционное учение - это совокупность представлений о механизмах и закономерностях исторических изменений (эволюции) в живой природе. Предметом изучения эволюционного учения являются фактор
... системы классификации К. Линнея, селекции растений и животных, сравнительной анатомии, сравнительной эмбриологии, сравнительной биохимии гипотезами, которые в совокупности составляют достаточно обоснованную теорию. 3. Эволюционное учение свободных падений Само понятие номогенез, и аргументы в пользу того, что, вопреки Дарвину, эволюция отнюдь не случайный, но закономерный процесс подробно ...
... – основную личностную (Космистскую) функциональность, практическое осуществление которой единственно обеспечивает ему гармоничный (здоровый) психосоматический процесс жизни (онтогенез) в целом. Пробил "эволюционный час" российской философии. Как когда-то, в 15-19 веках, достижения западных мыслителей (в свою очередь основывающихся на макро-эволюционных достижениях восточных и южных философов) ...
0 комментариев