1. Эволюционное учение Ж. Ламарка. 2
2. Эволюционная теория естественного отбора Ч. Дарвина. 4
3. Эволюционное учение свободных падений. 8
4. Вопросы происхождения основных царств живой природы.. 13
5. Происхождение многоклеточности у животных и растений. 15
6. Вопрос о переходных теориях организма. 17
7. Антропогенез. 19
Список литературы.. 22
1. Эволюционное учение Ж. Ламарка
В 1809 году вышла в свет "Философия зоологии" французского учёного Жана Батиста Пьера Антуана де Моне Ламарка (1744-1829). В этой работе была предпринята первая попытка создания теории эволюции видов. Она оказалась неудачной. Ламарк построил свою теорию на следующих двух положениях:
Во всех живых существах заложено внутреннее стремление к совершенствованию. Оно является движущей силой эволюции. Действием этого фактора определяется развитие живой природы, постепенное, но неуклонное повышение организации живых существ от простейших до самых совершенных. Его результатом является одновременное существование в природе организмов разной степени сложности, как бы образующих иерархическую лестницу существ.
Внешняя среда непосредственным образом влияет на изменение формы тех или органов живых существ. "У каждого животного, не завершившего ещё своего развития, более частое и продолжительное употребление какого-нибудь органа укрепляет этот орган, развивает его, увеличивает и придаёт ему силу, пропорциональную продолжительности употребления, тогда как постоянное отсутствие употребления какого-либо органа постепенно его ослабляет, приводит к упадку и заставляет его исчезнуть".
Он ошибался так как, естественно, не знал различий между генотипом и фенотипом. Упражнением или неупражнением можно изменить фенотип, но не генотип. Чтобы доказать, что фенотипические изменения не передаются по наследству, А. Вейсман провёл многолетний эксперимент, в котором на протяжении многих последовательных поколений укорачивал мышам хвосты. По теории Ламарка вынужденное неупотребление хвостов должно было привести к их укорочению у потомства, но этого не произошло. Вейсман на этом основании постулировал, что признаки, приобретаемые телом и приводящие к изменению фенотипа, не оказывают прямого воздействия на половые клетки (гаметы), с помощью которых признаки передаются следующему поколению.
В противоположность Ж. Ламарку Ж Кювье, исходя из особенностей строения нервной системы, сформулировал в 1812 г. учение о 4 "ветвях" (типах) организации животных: позвоночные, членистые, мягкотелые и лучистые. Между этими типами он не признавал никаких связей и переходов. В пределах типа позвоночных он различал 4 класса: млекопитающие, птицы, земноводные, рыбы. Описал большое число ископаемых форм и выявил принадлежность многих из них к определённым слоям земной коры.Ж. Кювье впервые предложил по ископаемым останкам определять возраст геологических слоёв, и наоборот. Основываясь на принципах "корреляции органов" и "функциональной корреляции", он разработал метод реконструкции ископаемых форм по немногим сохранившимся фрагментам скелета.
Отстаивая религиозные представления о сотворении и неизменности видов и отсутствии переходных форм между видами, он, для объяснения смены фаун и флор, наблюдаемых в последовательных геологических пластах, выдвинул теорию катастроф. В катастрофизме геологическая история Земли рассматривается как чередование длительных эпох относительного покоя и сравнительно коротких катастрофических событий, резко преображавших лик планеты. Согласно этой теории, в результате стихийных периодических бедствий на значительной части земного шара погибало всё живое, после чего его поверхность заселялась новыми формами, пришедшими из других мест. Концепция катастрофизма и неоднократных творческих актов согласовывалась с библейской концепцией творения мира. Благодаря идеям Ж. Кювье, широко распространились идеи о прогрессе в органическом мире и об эпизодических событиях, нарушающих однообразие в истории Земли. Это способствовало формированию представлений о сочетании эволюционного и скачкообразного развития.
В 1830-33 г. был издан труд английского естествоиспытателя Ч. Лайелла "Основы геологии". В этой работе в противовес популярной в то время теории катастрофизма он разработал учение о медленном и непрерывном изменении земной поверхности под влиянием постоянных геологических факторов, действующих и в современную эпоху (атмосферные осадки, текучие воды, извержения вулканов и т.д.). Эволюционная теория Ч. Лайелла (актуализм) явилась крупным шагом на пути к материалистическому пониманию природы. Ч. Лайелл считал действующие на Земле силы постоянными по качеству и интенсивности, не видел их изменения во времени и связанного с этим развития Земли.
2. Эволюционная теория естественного отбора Ч. Дарвина
Чарльз Дарвин в своем основном труде "Происхождение видов путем естественного отбора" (1859) обобщил эмпирический материал современной ему биологии и селекционной практики, на основе результатов собственных наблюдений во время путешествий, кругосветного плавания на корабле "Бигль", раскрыл основные факторы эволюции органического мира. В книге "Изменение домашних животных и культурных растений" (т.1-2, 1868) он изложил дополнительный фактический материал к основному труду, а в книге "Происхождение человека и половой отбор" (1871) выдвинул гипотезу происхождения человека от обезьяноподобного предка. В основе теории Дарвина - свойство организмов повторять в череде поколений сходные типы обмена веществ и индивидуального развития в целом - свойство наследственности.
Наследственность вместе с изменчивостью обеспечивает постоянство и многообразие форм жизни и лежит в основе эволюции живой природы. Одно из основных понятий своей теории эволюции - понятие "борьба за существование" - Дарвин употреблял для обозначения отношений между организмами, а также отношений между организмами и абиотическими условиями, приводящих к гибели менее приспособленных и выживанию более приспособленных особей.
Понятие "борьба за существование" отражает те факты, что каждый вид производит больше особей, чем их доживает до взрослого состояния, и что каждая особь в течение своей жизнедеятельности вступает во множество отношений с биотическими и абиотическими факторами среды.
Дарвин выделил две основные формы изменчивости:
Определенную изменчивость - способность всех особей одного и того же вида в определенных условиях внешней среды одинаковым образом реагировать на эти условия (климат, почву).
Неопределенную изменчивость, характер которой не соответствует изменениям внешних условий. В современной терминологии неопределенная изменчивость называется мутацией.
Мутация - неопределенная изменчивость в отличие от изменчивости определенной носит наследственный характер. По Дарвину, незначительные изменения в первом поколении усиливаются в последующих поколениях. Дарвин подчеркивал, что решающую роль в эволюции играет именно неопределенная изменчивость. Она связана обычно с вредными и нейтральными мутациями, но возможны и такие мутации, которые оказываются перспективными.
Неизбежным результатом борьбы за существование и наследственной изменчивости организмов, по Дарвину, является процесс выживания и воспроизведения организмов, наиболее приспособленных к условиям среды, и гибели в ходе эволюции неприспособленных - естественный отбор.
Механизм естественного отбора в природе действует аналогично селекционерам, то есть складывает незначительные и неопределенные индивидуальные различия и формирует из них у организмов необходимые приспособления, а также межвидовые различия. Этот механизм выбраковывает ненужные формы и образовывает новые виды.
Тезис о естественном отборе наряду с принципами борьбы за существование, наследственности и изменчивости - основа дарвиновской теории эволюции.
Во времена Дарвина наследственность представляли как некое общее свойство организма, присущее ему как целому. В связи с этим шотландский исследователь Флеминг Дженкинс вошел в историю биологии, выдвинув возражения против теории Дарвина. Он считал, что новые полезные признаки некоторых особей данного вида должны быстро исчезнуть при скрещивании с другими, более многочисленными особями. Возражения Дженкинса сам Дарвин считал очень серьезными, окрестив их "кошмаром Дженкинса". Эти возражения были опровергнуты, только когда стало ясно, что аппарат наследственности сформирован отдельными структурными и функциональными единицами - генами.
Согласно Ч. Дарвину и А. Уоллесу, механизмом, с помощью которого из предсуществовавших видов возникают новые виды, служит естественный отбор. Эта гипотеза основана на трёх наблюдениях и двух выводах:
Наблюдение 1. Особи, входящие в состав популяции, обладают большим репродуктивным потенциалом. Дарвин и Уоллес зафиксировали этот факт во многом благодаря работе Т. Мальтуса "Трактат о народонаселении". В ней Т. Мальтус привлёк внимание к репродуктивному потенциалу человека и отметил, что численность народонаселения возрастает по экспоненте.
Наблюдение 2. Число особей в каждой данной популяции примерно постоянно. Численность всех популяций ограничивается или контролируется различными факторами среды, такими как пищевые ресурсы, пространство и свет. Размеры популяции возрастают до тех пор, пока среда ещё может выдерживать их дальнейшее увеличение, после чего достигается некое равновесие. Численность колеблется около этого равновесного уровня.
Вывод 1. Многим особям не удаётся выжить и оставить потомство. В популяции происходит "борьба за существование". Непрерывная конкуренция между индивидуумами за факторы среды в пределах одного вида или между представителями разных видов приводит к тому, что некоторые организмы не смогут выжить или оставить потомство.
Наблюдение 3. Во всех популяциях существует изменчивость. Огромный фактический материал, собранный во время путешествий Дарвином и Уоллесом убедили их в значимости внутривидовой изменчивости. Но они не смогли выявить источники всех этих форм изменчивости. Это позже сделает Г. Мендель.
Вывод 2. В "борьбе за существование" те особи, признаки которых наилучшим образом приспособлены к условиям жизни, обладают "репродуктивным преимуществом" и производят больше потомства, чем менее приспособленные особи. Решающий фактор, определяющий выживание, - это приспособленность к среде. Любое, самое незначительное физическое, физиологическое или поведенческое изменение, дающее одному организму преимущество перед другим, будет действовать как "селективное преимущество". Благоприятные изменения будут передаваться следующим поколениям, а неблагоприятные отметаются отбором, так как они невыгодны организму. Вывод 2 содержит гипотезу о естественном отборе, который может служить механизмом эволюции.
Теория эволюции, предложенная Дарвином и Уоллесом, была расширена и разработана в свете современных данных генетики, палеонтологии, молекулярной биологии, экологии и этологии и получила название неодарвинизма. Его можно определить как теорию органической эволюции путём естественного отбора признаков, детерминированных генетически. Чтобы признать эту теорию достоверной необходимо:
Установить факт изменения форм жизни во времени (эволюция в прошлом).
Выявить механизм, производящий эволюционные изменения (естественный отбор генов)
Продемонстрировать эволюцию, происходящую в настоящее время (эволюция в действии).
Свидетельствами эволюции, происходившей в прошлом, служат ископаемые остатки организмов и стратиграфия. Данные о механизме эволюции получают путём экспериментальных исследований и наблюдений, касающихся естественного отбора наследуемых признаков и механизма наследования, демонстрируемого классической генетикой. Сведения о действии этих процессов в наше время даёт изучение популяций современных организмов, результаты искусственного отбора и генной инженерии.
Пока не существует твёрдо установленных законов эволюции. Мы обладаем лишь хорошо подкреплёнными фактическими данными из области палеонтологии, географии распространения видов, системы классификации К. Линнея, селекции растений и животных, сравнительной анатомии, сравнительной эмбриологии, сравнительной биохимии гипотезами, которые в совокупности составляют достаточно обоснованную теорию.
3. Эволюционное учение свободных падений
Само понятие номогенез, и аргументы в пользу того, что, вопреки Дарвину, эволюция отнюдь не случайный, но закономерный процесс подробно обосновал Л.С. Берг в своих классических работах 20-х годов, из которых главная и наиболее известная - "Номогенез, или эволюция на основе закономерностей". Берг формулирует проблему так:
Является ли эволюция случайным процессом, который обусловлен лишь двумя факторами: хаотическими мутациями и естественным отбором, или же эволюция - это процесс в своей основе закономерный, выявление некоторой тенденции, имманентного закона, который и направляет ее ход?
В такой постановке вопрос может показаться не вполне корректным и даже беспредметным, ведь и случайные в своей основе процессы могут подчиняться весьма строгим статистическим законам. Более точно его суть можно уяснить из простой аналогии: хотя на развитие отдельного оргaнизмa влияет множество случайных факторов, но нет сомнения и в том, что главным, определяющим является фактор внутренней информации, заложенной в генах. Вся его история есть рaзворaчивaние, реaлизaция программы, от которой только и зависит, что же вырастет, например, из данного семени береза или сосна.
Вся эволюция биосферы есть, согласно Бергу разворачивание, какого-то закона, или может быть правильнее сказать, многовариантной программы, в которой содержатся и многочисленные способы ее реализации. Поэтому Берг и назвал свою концепцию номогенезом, противопоставив ее дарвиновской концепции "тихогенеза", то есть развитию, основанному на случайности. Можем ли мы сегодня, хотя бы в самых общих контурах представить себе, как же выглядит этот закон? Нет, но наше незнание вовсе не означает, что такого закона нет.
Представим себе, что некий математик, исследующий таблицы случайных чисел, с удивлением обнаруживает в них устойчивые повторения, "мотивы", "ритмы и рифмы", "гомологии", присутствие которых никак нельзя объяснить игрой случая. Пусть далее нечто подобное он сможет найти и в других последовательностях, полученных с помощью независимых и различных по устройству генераторов. Какую гипотезу вправе выдвинуть такой математик? Он может, прежде всего, предположить, что исследуемые им ряды вовсе не случайны, но есть достаточно замысловатое проявление неизвестной ранее природной закономерности.
В своих работах Берг суммирует огромный фактический материал, накопленный уже к началу 20 века, который и свидетельствует в пользу номогенетической природы эволюции. Этот материал говорит о присутствующих в системе форм живого многочисленных "ритмах и рифмах", которые невозможно назвать случайными. В качестве примера приведём факт так называемого предварения признаков (филогенетическое ускорение). Известно, что в эмбриональной фазе наблюдаются признаки тех стадий, через которые предположительно прошла эволюция данной группы. В свое время Э. Геккель, горячий сторонник и пропагандист дарвинизма, сформулировал правило, получившее название биогенетического закона: онтогения повторяет филогению. Почему-то считается, что он служит прямым аргументом в пользу дарвиновской концепции, хотя его можно понимать лишь как свидетельство того, что эволюция вообще имеет место, в чем, конечно же, мало кто сомневается.
Гораздо реже обсуждается факт, что имеет место и обратное, симметричное по времени явление: "индивидуальное развитие может не только повторять филогению, но и предварять ее". Это правило применимо не только к отдельным организмам, но и к целым их группам: филогения какой-либо группы может опережать свой век, осуществляя формы, которые в норме свойственны более высоко стоящим в системе организмам. Это значит, что признаки, которые появляются в результате предварения, не могли получиться как результат действия дарвиновского механизма. Как индивидуальное развитие, эволюция есть процесс разворачивания, реализации уже существующей программы.
Номогенетическое направление в теории эволюции аналогично номотетическому в систематике. Для лучшего понимания полезно сказать несколько слов о классификации вариантов эволюционных теорий:
номогенетический - наличие специфических законов развития или ограниченности формообразования;
эктогенетический - роль внешних факторов в эволюции;
телогенетический - роль активной адаптации.
Теорию номогенеза можно разделить на два направления:
Учение об ограниченности формообразования.
Изменчивость во времена Дарвина считалась неограниченной, подобно "восковой пластичности". Это доказывалось тем, что любой признак показывал большую или меньшую изменчивость. Задолго до Менделя заметили, что при плодовитом скрещивании наблюдается какая-то особенно расшатанная изменчивость, как будто не подчиняющаяся никаким законам. Мендель этот мнимый хаос подчинил строгим математическим законам: при этом вместо восковой пластичности, допускающей бесконечно большое число возможных модальностей, мы получаем не только конечное, но даже не очень большое число их. Несколько поднявшись по таксономическому уровню, мы встречаем закон гомологических рядов Н.И. Вавилова (1920).
Третьей формой номогенеза в этом понимании можно считать то, что называется биохимическим номогенезом, например, биохимические отличия первично - и вторичноротых. Важное вещество, используемое первичноротыми для построения наружного скелета - хитин - полностью отсутствует в двух главных типах вторичноротых - у иглокожих и позвоночных. У иглокожих и позвоночных развивается внутренний известковый скелет на соединительно-тканной основе и сгибаемость корпуса или антимер получается от сочленения внутренних известковых элементов или позвонков (позвоночный столб позвоночных и руки).
Наконец, четвертую форму номогенеза в смысле ограниченности формообразования можно назвать телогенетическим номогенезом, то есть сходным разрешением определенных задач, независимо от природы факторов, осуществляющих это разрешение. Это явление давно известно (сходство ихтиозавров, дельфинов и рыб, глаз позвоночных и головоногих), но вся широта осознается недостаточно и некоторые интересные направления мало известны.
Учение об ограниченности формообразования намечает возможности прогноза форм живых существ на других планетах. Здесь наблюдается огромный разнобой мнений от допущения вероятности возникновения и существования организмов, настолько близких человеку, что возможно продуктивное скрещивание, до отрицания какого-либо предвидения особенностей строения инопланетных существ. Первая крайность вряд ли может защищаться серьезным биологом, но такие образования, как, скажем, ДНК, хромосомы, клетка, кишечник, метамерия, конечности, хитин, возникают или могут возникать в очень сходном виде не только параллельно, но и конвергентно*. Принципиальная возможность существования растений и животных на других планетах не отрицается, следовательно, для высших таксонов мыслимо самостоятельное возникновение.
Можно предположить, что типы, а может быть и некоторые классы, могут возникнуть самостоятельно, и на далеких планетах мы вправе ожидать встречи с организмами, которые мы отнесем к простейшим, кишечнополостным, аннелидам, членистоногим и даже насекомым. Встретив на другой планете разумное существо, мы, конечно, отличим его от человека. Но, скорее всего, кое-какие признаки сходства будут: у него впереди будет голова, в которой будет развитой мозг, будут парные глаза, построенные согласно требованиям геометрической оптики, будут парные конечности, передние конечности будут орудиями труда, а не передвижения, значит, будут иметь подобие пальцев, хотя число и строение этих пальцев могут быть совершенно отличными от наших. Одно из названий геометрии Лобачевского было "воображаемая геометрия". Сейчас назрела надобность в "воображаемой биологии"
Учение о направленных путях развития.
Ограниченность формообразования не накладывает никаких ограничений на форму путей развития. Но наряду с ненаправленной, зигзагообразной эволюцией, которая, несомненно, существует на низшем уровне эволюции, существуют и направленные формы, которые давно обозначались разными терминами, особенно популярен термин "ортогенез", или прямолинейное развитие. Ортодоксальные дарвинисты резко критиковали теорию ортогенеза: отрицалось, что ортогенез есть главный и даже единственный модус эволюции, и это возражение вполне резонно; критиковалось причинное объяснение ортогенеза как результата воздействия внешних условий и предлагался другой термин - "ортоэволюция"; указывались случаи развития по спирали, где говорить о прямолинейном развитии не приходится; наконец, так как главное свидетельство ортогенеза - параллельное развитие - получает все более мощную фактическую поддержку, дарвинисты пытаются спасти положение, объясняя этот параллелизм селекцией в том же направлении, предложив термин "ортоселекция".
Конечно, термины "ортоселекция", "ортогенез" и прочие не вполне точны. Но есть и другая форма номогенетического направленного развития, которую можно назвать номогенезом только в начальной стадии, подобно тому, как ядро летит из пушки первое время почти по прямой линии. Эта форма развития заключает три отрезка:
Очень быстрая прогрессивная эволюция, частный случай - ароморфозы по терминологии А.Н. Северцева;
Переход вертикальной линии в горизонтальную - консервативная эволюция;
Идиодаптация Северцева - это частный случай, а не общий закон, так как эволюция может и не быть адаптивной;
Регрессивный этап: потеря изменчивости, регрессивное развитие и вымирание. Видимо, в ряде случаев возможен выход из тупика эволюции путем педогенеза или иначе.
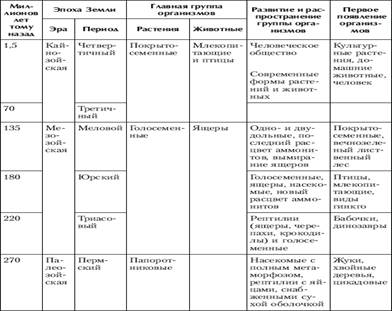
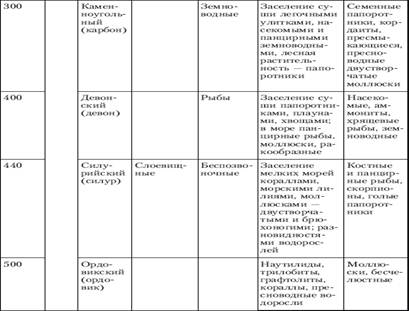
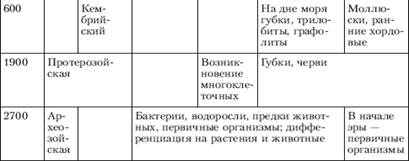
4. Вопросы происхождения основных царств живой природы
Единицей классификации, как для растений, так и для животных, служит вид. Можно в самом общем смысле определить вид как популяцию особей, обладающих сходными морфологическими и функциональными признаками, имеющих общее происхождение и в естественных условиях скрещивающихся только между собой.
Можно также определить вид как совокупность популяций, внутри которых возможно скрещивание или как группу популяций с общим генофондом. Любое из этих определений подразумевает как главное: один вид отделён от другого репродуктивной преградой, между ними невозможно скрещивание.
Решить проблему видообразования - значит, объяснить каким образом элементарные эволюционные изменения в популяции могут привести к образованию новых видов, родов, семейств и отрядов и как возникают преграды, препятствующие скрещиванию между зарождающимися видами. Всякий фактор, затрудняющий скрещивание между группами или организмами, называется изолирующим механизмом.
Один из самых обычных видов изоляции - это географическая изоляция, при которой группы родственных организмов бывают разделены какой-то физической преградой. Например, в горах на данную площадь обычно приходится больше разных видов, чем на такую же площадь на равнине. Как правило, географическая изоляция не бывает постоянной: разобщённые близкородственные группы иногда вновь встречаются и могут продолжить скрещивание, если только за это время между ними не возникло генетической изоляции, то есть стерильности при скрещивании. Генетическая изоляция бывает обусловлена мутациями, возникающими случайно, независимо от других мутаций, влияющих на морфологические или физиологические признаки. Поэтому в одних случаях она может наступить очень нескоро, когда длительная географическая изоляция создаст заметные различия между двумя группами организмов, а в других случаях может возникнуть в пределах одной, во всём остальном гомогенной группы.
Обычно потомки от скрещивания между разными видами бывают стерильны, однако, иногда в результате гибридизации представителей двух разных, но очень близких видов, возникает новый вид. Гибридная форма может объединить в себе лучшие признаки обоих родительских видов, в результате чего получится новая форма, лучше приспособленная к среде, чем каждая из исходных форм или, наоборот, - худшие признаки с соответствующим исходом.
Изоляция, необходимая на начальных стадиях видообразования, может обеспечиваться не только географическими преградами между популяциями: иногда обособленные группировки особей возникают в пределах одной популяции, и это может привести к формированию новых видов. Такой способ видообразования называется "симпатрическим" (от латинских слов sim - вместе и patria - родина). Этот способ отличается от предыдущего только факторами изоляции, причины же, приводящие к морфологической дивергенции и становлению системы изолирующих механизмов, те же, что и в случае географического видообразования.
При экологическом видообразовании изолирующим фактором являются естественный отбор (особая его форма - дизруптивный или раздробляющий отбор) в сочетании с неоднородностью среды обитания. Для успешного завершения процесса видообразования изоляция должна быть как можно более полной и существовать длительное время. Эти условия в природной обстановке трудновыполнимы, поэтому примеры экологического видообразования довольно редки.
Теоретически экологическое видообразование может происходить и при отсутствии первичной изоляции между зарождающимися видами. Для этого необходимо, чтобы в популяции действовал дизруптивный отбор, непосредственно направленный на формирование системы изолирующих механизмов. Такой вывод сделан на основе анализа компьютерных моделей и подтверждается в экспериментах с плодовой мушкой дрозофилой. Вероятно, что именно таким образом - в результате экологического видообразования без первичной изоляции - произошли комплексы близкородственных видов рыб в изолированных озерах.
5. Происхождение многоклеточности у животных и растений
Эволюция каждой данной формы живых организмов происходит на протяжении многих поколений. За это время многие особи рождаются и умирают, но популяция сохраняет непрерывность. Таким образом, эволюционирующей единицей оказывается не особь, а популяция. Популяция сходных особей, живущих на ограниченной территории и скрещивающихся между собой, называется демом или генетической популяцией. Следующей более крупной категорией служит вид, состоящий из ряда слабо разграниченных демов.
В природе демы и виды имеют тенденцию оставаться неизменными на протяжении многих поколений. Такая неизменность означает, что за это время не произошло никаких изменений ни в генетической конституции дема, ни в условиях окружающей среды влияющих на выживание данных организмов. Каждая популяция характеризуется определённым генофондом. Каждая особь в популяции в генетическом отношении уникальна.
Процесс видообразования должен состоять из двух неразрывных составляющих:
Отклонения формы (центробежная составляющая).
Удержания формы - (центростремительная составляющая).
Образование новой формы - как таксономической реальности представляет собой результат взаимодействия этих двух процессов.
Отсюда два вопроса:
Первый вопрос - какова природа изменчивости, то есть что является ее внутренним источником, каковы ее свойства, и что провоцирует изменчивость (в данном контексте, какие внешние обстоятельства).
Второй вопрос - (особенно важный при принятии идеи трансформизма) - как удерживается фактическое разнообразие, то есть, что определяет постоянство формы в поколениях - на исторической и, более того, геологической шкале.
Можно говорить о "внутренних" причинах и "внешних" условиях видообразования. Два факта - факт разнообразия форм (видов, пород, сортов, особей) и факт постоянства формы в одной генеалогической линии - привели к пониманию изменчивости и наследственности как двух независимых основополагающих факторов, в принципе определяющих возможность такого изменения форм, которое приводит к дискретному и устойчивому во времени разнообразию таксонов. В процессуальном плане изменчивость и наследуемость выступают как факторы центробежный (меняющий) и центростремительный (удерживающий). Их сложное взаимодействие определяет процесс и конечный результат (устойчивое изменение). Эти факторы называют "внутренними" причинами видообразования.
Есть "внешние условия", благоприятствующие (провоцирующие) или неблагоприятствующие (подавляющие - избирательно или тотально) фенотипическую изменчивость. Это:
а) демографический фактор, часто сопряженный с географическим или биотопическим фактором (в сумме - пространственное обособление малой группы или - появление малого изолята);
б) экологический фактор (физическая и репродуктивная выживаемость формы в конкретном экологическом контексте). Последний фактор выступает как мера достаточного соответствия формы среде обитания и как элиминирующий отбор при отсутствии такого соответствия.
Отношение к "внутренним" и "внешним" факторам видообразования в разных концепциях меняется. Это отношение и определяет суть концепций, то есть, их концептуальный арсенал, смысловое значение центральных понятий (часто одних и тех же терминологически) и лексические приемы.
6. Вопрос о переходных теориях организма
После повсеместного распространения учения Ч. Дарвина одним из первых критиков, указавших на слабое место в теории, был шотландский исследователь Ф. Дженкинс. В 1867 г. он заметил, что в дарвиновской теории нет ясности в вопросе о том, как осуществляется накопление в потомстве тех или иных изменений. Ведь сначала изменения признака происходят только у некоторых особей. После скрещивания с нормальными особями должно наблюдаться не накопление, а разбавление данного признака в потомстве. То есть в первом поколении остаётся ½ изменения, во втором - ¼ изменения и т.д. вплоть до полного исчезновения этого признака. Ч. Дарвин так и не нашёл ответа на этот вопрос.
Между тем решение этого вопроса существовало. Его получил преподаватель монастырской школы в Брно (Чехия) Г. Мендель. В 1865 году были опубликованы результаты его работ по гибридизации сортов гороха, где были открыты важнейшие законы наследственности. Автор показал, что признаки организмов определяются дискретными наследственными факторами.
Он ещё до выхода в свет книги Ч. Дарвина хотел проследить судьбу изменений генотипов в разных поколениях гибридов. Объектом исследования стал горох. Мендель взял два сорта гороха - с жёлтыми и с зелёными семенами. Скрестив эти два сорта, он обнаружил в первом поколении гибридов горох только с жёлтыми семенами. Путём самоопыления полученных гибридов он получил второе поколение. В нём появились особи с зелёными семенами, но их было заметно меньше, чем с жёлтыми. Подсчитав число тех и других, Мендель пришёл к выводу, что число особей с желтыми семенами относится к числу особей с зелёными как приблизительно 3:
1.
Параллельно он проводил серию других опытов с растениями, прослеживая какой-либо признак в нескольких поколениях. В каждом опыте в первом поколении проявлялся только один из родительских признаков. Мендель назвал его доминантным. Временно исчезающий признак он назвал рецессивным. Во всех опытах отношение числа особей с доминантным признаком к числу особей с рецессивным признаком среди гибридов второго поколения было в среднем равно 3:
1.
Итак, можно было утверждать, что при скрещивании растений с противоположными признаками происходит не разбавление признаков, а подавление одного признака другим, в связи с этим необходимо различать доминантные и рецессивные признаки.
Мендель пошёл в своих экспериментах дальше. Он произвёл самоопыление гибридов второго поколения и получил гибриды третьего, а затем и четвёртого поколения. Он обнаружил, что гибриды второго поколения с рецессивным признаком при дальнейшем размножении не расщепляются ни в третьем, ни в четвёртом поколениях. Так же ведёт себя примерно треть гибридов второго поколения с доминантным признаком. Две трети гибридов с доминантным признаком расщепляются при переходе к гибридам третьего поколения, причём опять-таки в отношении 3:
1. Получившиеся при этом расщеплении гибриды третьего поколения с рецессивным признаком и треть гибридов с доминантным признаком при переходе к четвёртому поколению не расщепляются, а остальные гибриды третьего поколения расщепляются, причём снова в отношении 3:
1.
Этот факт демонстрирует важное обстоятельство: особи с одинаковыми внешними признаками могут обладать разными наследственными свойствами, то есть по фенотипу нельзя судить с достаточной полнотой о генотипе. Если особь не обнаруживает в потомстве расщепления, то её называют гомозиготной, если обнаруживает - гетерозиготной.
В итоге Г. Менделем был сформулирован закон единообразия гибридов первого поколения: первое поколение гибридов в силу проявления у них лишь доминантных признаков всегда единообразно. Этот закон носит также название первого закона Менделя или закона доминирования. Однако результаты его исследований оставались практически неизвестными почти 35 лет - с 1865 по 1900.
7. Антропогенез
Антропоцентризм и биосферное мышление Антропоцентрическое мышление и биосферное мышление - два кардинально различающихся типа мировоззрения. Это касается:
характера проблем - методологических, исследовательских, хозяйственно-промышленных и т.д.;
множества людей - от отдельных личностей, групп людей, объединенных по социальной, религиозной, национальной или иной принадлежности, до населения стран, материков и человечества в целом;
размера территории, подвергающейся антропогенному воздействию - от десятков - сотен квадратных метров, частей ландшафта до обширных регионов, витасферы и биосферы в целом.
Одним из главных признаков различия двух мировоззрений является отношение к времени. При антропоцентрическом подходе, как правило, ограничиваются оценками и прогнозами краткосрочными - максимум ближайшее десятилетие, в то время как при биосферном основу должны составлять долгосрочные оценки и прогнозы - минимум десятилетия и столетия. Антропоцентризм делает акцент на судьбах ныне живущих людей и их сиюминутных интересах, и в крайнем случае - их детей и уж совсем абстрактно - внуков. В то время как биосферное мышление будет охватывать череду поколений и действительно приобретет, таким образом, право говорить о судьбе человечества.
Антропоцентризм локализует анализ воздействий на природные комплексы в пространстве. При биосферном подходе сознается важность возможного "расползания" эффектов на обширные территории. Антропоцентрический подход, реализуемый в каком-то промышленном проекте, предъявляет своим противникам требование: "Докажите, что этот проект будет в каком-то отношении вредным". Биосферный подход требует аргументов в пользу того, что наличествующее состояния природы не будет ухудшено. В конечном итоге антропоцентризм формулирует целевую функцию, как "было бы лучше человеку сегодня, а там видно будет", биосферное мышление - "не может быть человеку лучше, если не исключено ухудшение природных комплексов".
Опыт показывает, что антропоцентрический подход довольствуется остаточным принципом финансирования фундаментальных исследований, являющихся, по словам В.И. Вернадского, основой формирования биосферного мышления: "Основной геологической силой, создающей ноосферу, является рост научного знания".
Список литературы
1. Горелов А.А. Концепции современного естествознания: Учебное пособие для студентов высших учебных заведений, обучающихся по гуманитарным специальностям. – М.: Гуманит. изд. центр ВЛАДОС, 2002.
2. Концепция современного естествознания: Под ред. профессора С.И. Самыгина. Изд. третье. Ростов н/Д: "Феникс", 2001 – 576 с.
3. Алексеев В.П. Становление человечества. М., 1984.
4. Бор Н. Атомная физика и человеческое познание. М., 1961 Борн М. Эйнштейновская теория относительности. М., 1964.
5. Вайнберг С. Первые три минуты. Современный взгляд на происхождение Вселенной.М., 1981.
6. Дорфман Я.Г. Всемирная история физики с начала 19 века до середины 20 века. М., 1979.
7. Кемпфер Ф. Путь в современную физику. М., 1972.
8. Мэрион Дж.Б. Физика и физический мир. М., 1975
9. Найдыш В.М. Концепции современного естествознания. Учебное пособие.М., 1999.
10. Николис Г., Пригожин И. Познание сложного. М., 1990.
11. Пригожин И. От существующего к возникающему. М., 1985.
12. Степин В.С. Философская антропология и философия науки. М., 1992.
13. Фейнберг Е.Л. Две культуры. Интуиция и логика в искусстве и науке. М., 1992.
Похожие работы
... виде, или постепенно, в течение «дней» «производить» из воды и земли, от низших форм к высшим силою заложенных Им в природу законов». 3. Критика эволюционного учения и креационизма Теория эволюции подвергается креационистами критике, в основном, по следующим направлениям. 1. Палеонтологическая летопись обнаруживает скорее структуру эволюционных скачков, а не постепенные преобразования. ...
... отношений, появление более совершенных орудий труда, жилища, одежды привели к снижению роли биологического фактора эволюции и усилению ведущего действия социальных законов развития[5]. Заключение Эволюционное учение - это совокупность представлений о механизмах и закономерностях исторических изменений (эволюции) в живой природе. Предметом изучения эволюционного учения являются фактор
... рядов) Н. И. Вавилова привело к формулировке гипотез об эволюции на основе закономерностей, а не случайной изменчивости (номогенез Л. С. Берга, батмогенез Э. Д. Копа и др.). В 1920-1940-е г. г. в эволюционной биологии интерес к селекционистским теориям возродился благодаря синтезу классической генетики и теории естественного отбора. Разработанная в результате этого синтетическая теория эволюции ...
... – основную личностную (Космистскую) функциональность, практическое осуществление которой единственно обеспечивает ему гармоничный (здоровый) психосоматический процесс жизни (онтогенез) в целом. Пробил "эволюционный час" российской философии. Как когда-то, в 15-19 веках, достижения западных мыслителей (в свою очередь основывающихся на макро-эволюционных достижениях восточных и южных философов) ...
0 комментариев