Навигация
ТЕОРИЯ ЕСТЕСТВЕННОГО ОТБОРА
36102
знака
0
таблиц
2
изображения
3. ТЕОРИЯ ЕСТЕСТВЕННОГО ОТБОРА
ФОРМЫ ЕСТЕСТВЕННОГО ОТБОРА
Выживание наиболее приспособленных особей Дарвин называл естественным отбором. Его не следует понимать как какой-то выбор, так как здесь мы имеем лишь естественное следствие гибели менее приспособленных. Естественный отбор реализуется через действие естественных факторов среды (температура, влажность, свет, паразиты, конкуренты, враги, трудности добывания пищи и т. п.), действует через сохранение и накопление мелких наследственных изменений. Например, даже незначительное удлинение хоботка у пчелы делает ее более способной добывать нектар из цветков, недоступных для других пчел. Пчелы с удлиненным хоботком и будут иметь преимущество по сравнению с теми, у которых сохраняется .в поколении обычная длина этого органа [9].
Отбор происходит непрерывно на протяжении бесконечного ряда следующих друг за другом поколений и сохраняет главным образом те формы, которые в большей мере соответствуют данным условиям. Естественный отбор и элиминация части особей вида неразрывно связаны между собой и являются необходимым условием эволюции видов в природе.
Схема действия естественного отбора в системе вида по Дарвину сводится к следующему:
1) Изменчивость свойственна любой группе животных и растений, и организмы отличаются друг от друга во многих отношениях;
2) Число организмов каждого вида, рождающихся на свет, превышает число тех, которые могут найти пропитание и выжить. Тем не менее, поскольку численность каждого вида в естественных условиях постоянна, следует предполагать, что большая часть потомства гибнет. Если бы все потомки какого-либо вида выживали и размножались, то весьма скоро они вытеснили бы все другие виды на земном шаре;
3) Поскольку рождается больше особей, чем может выжить, происходит борьба за существование, конкуренция за пищу и место обитания. Это может быть активная борьба не на жизнь, а на смерть, или менее явная, но не менее действенная конкуренция, как, например, для растений в период засухи или холода;
4) Среди множества изменений, наблюдающихся у живых существ, одни облегчают выживание в борьбе за существование, другие же приводят к тому, что их обладатели гибнут. Концепция «выживания наиболее приспособленных» представляет собой ядро теории естественного отбора;
5) Выживающие особи дают начало следующему поколению, и таким образом «удачные» изменения передаются последующим поколениям. В результате каждое следующее поколение оказывается более приспособленным к среде обитания; по мере изменения среды возникают дальнейшие приспособления. Если естественный отбор действует на протяжении многих лет, то последние отпрыски могут оказаться настолько несхожими со своими предками, что их целесообразно будет выделить в самостоятельный вид [5].
Может также случиться, что некоторые члены данной группы особей приобретут одни изменения и окажутся приспособленными к окружающей среде одним способом, тогда как другие ее члены, обладающие другим комплексом изменений, окажутся приспособленными иначе; таким путем от одного предкового вида при условии изоляции подобных групп может возникнуть два и более видов.
Движущий отбор. Естественный отбор всегда ведет к увеличению средней приспособленности популяций. Изменение внешних условий может приводить к изменению приспособленности отдельных генотипов. В ответ на эти изменения, естественный отбор, используя огромный запас генетического разнообразия по множеству разных признаков, ведет к значительным сдвигам в генетической структуре популяции. Если внешняя среда меняется постоянно в определенном направлении, то естественный отбор меняет генетическую структуру популяции таким образом, чтобы ее приспособленность в этих меняющихся условиях оставалась максимальной. При этом меняются частоты отдельных аллелей в популяции. Меняется и средние значения приспособительных признаков в популяциях. В ряду поколений прослеживается их постепенное смещение в определенном направлении. Такую форму отбора называют движущим отбором.
Классическим примером движущего отбора является эволюция окраски у березовой пяденицы. Окраска крыльев этой бабочки имитирует окраску покрытой лишайниками коры деревьев, на которых она проводит светлое время суток. Очевидно, такая покровительственная окраска сформировалась за многие поколения предшествующей эволюции. Однако с началом индустриальной революции в Англии это приспособление стало терять свое значение. Загрязнение атмосферы привело к массовой гибели лишайников и потемнению стволов деревьев. Светлые бабочки на темном фоне стали легко заметны для птиц. Начиная с середины XIX века, в популяциях березовой пяденицы стали появляться мутантные темные (меланистические) формы бабочек. Частота их быстро возрастала. К концу XIX века некоторые городские популяции березовой пяденицы почти целиком состояли из темных форм, в то время как в сельских популяциях по-прежнему преобладали светлые формы. Это явление было названо индустриальным меланизмом. Ученые обнаружили, что в загрязненных районах птицы чаще поедают светлые формы, а в чистых – темные. Введение ограничений на загрязнение атмосферы в 1950-х годах привело к тому, что естественный отбор вновь изменил направление, и частота темных форм в городских популяциях начала снижаться. В наше время они почти так же редки, как и до начала индустриальной революции.
Движущий отбор приводит генетический состав популяций в соответствие изменениям во внешней среде так, чтобы средняя приспособленность популяций была максимальной. На острове Тринидад рыбки гуппи обитают в разных водоемах. Множество тех, что живут в низовьях речек и в прудах гибнет в зубах хищных рыб. В верховьях жизнь для гуппи гораздо спокойней – там мало хищников. Эти различия во внешних условиях привели к тому, что «верховые» и «низовые» гуппи эволюционировали в разных направлениях. «Низовые», находящиеся под постоянной угрозой истребления, начинают размножаться в более раннем возрасте и производят множество очень мелких мальков. Шанс на выживание каждого из них очень невелик, но их очень много и некоторые из них успевают размножиться. «Верховые» достигают половой зрелости позднее, их плодовитость ниже, но потомки крупнее. Когда исследователи переносили «низовых» гуппи в незаселенные водоемы в верховьях речек, они наблюдали постепенное изменение типа развития рыбок. Через 11 лет после перемещения они стали значительно крупнее, вступали в размножение позже и производили меньшее количество, но более крупных потомков.
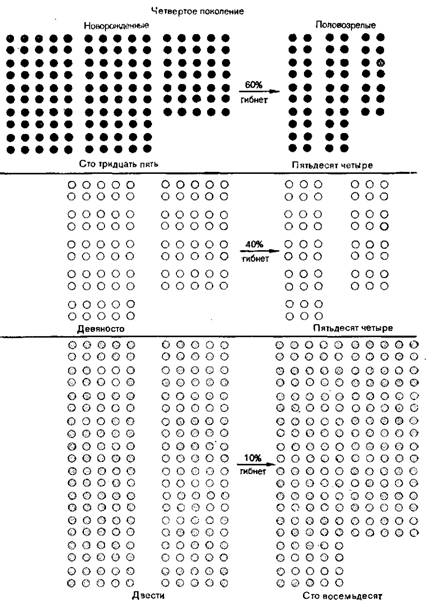
Скорость изменения частот аллелей в популяции и средних значений признаков при действии отбора зависит не только от интенсивности отбора, но и от генетической структуры признаков, по которым идет обор. Отбор против рецессивных мутаций оказывается значительно менее эффективным, чем против доминантных. В гетерозиготе рецессивный аллель не проявляется в фенотипе и поэтому ускользает от отбора. Используя уравнение Харди-Вейнберга можно оценить скорость изменения частоты рецессивного аллеля в популяции в зависимости от интенсивности отбора и начального соотношения частот. Чем ниже частота аллеля, тем медленнее происходит его элиминация. Для того чтобы снизить частоту рецессивной летали от 0,1 до 0,05 нужно всего 10 поколений; 100 поколений - чтобы уменьшить ее от 0,01 до 0,005 и 1000 поколений - от 0,001 до 0,0005 [2].
Движущая форма естественного отбора играет решающую роль в приспособлении живых организмов к меняющимся во времени внешним условиям. Она же обеспечивает широкое распространение жизни, ее проникновение во все возможные экологические ниши. Ошибочно думать, однако, что в стабильных условиях существования естественный отбор прекращается. В таких условиях он продолжает действовать в форме стабилизирующего отбора.
Стабилизирующий отбор. Стабилизирующий отбор сохраняет то состояние популяции, которое обеспечивает ее максимальную приспособленность в постоянных условиях существования. В каждом поколении удаляются особи, отклоняющиеся от среднего оптимального значения по приспособительным признакам.
Описано множество примеров действия стабилизующего отбора в природе. Например, на первый взгляд кажется, что наибольший вклад в генофонд следующего поколения должны вносить особи с максимальной плодовитостью. Однако наблюдения над природными популяциями птиц и млекопитающих показывают, что это не так. Чем больше птенцов или детенышей в гнезде, тем труднее их выкормить, тем каждый из них меньше и слабее. В результате наиболее приспособленными оказываются особи со средней плодовитостью.
Отбор в пользу средних значений был обнаружен по множеству признаков. У млекопитающих новорожденные с очень низким и очень высоким весом чаше погибают при рождении или в первые недели жизни, чем новорожденные со средним весом. Учет размера крыльев у птиц, погибших после бури, показал, что большинство из них имели слишком маленькие или слишком большие крылья. И в этом случае наиболее приспособленными оказались средние особи.
В чем причина постоянного появления малоприспособленных форм в постоянных условиях существования? Почему естественный отбор не способен раз и навсегда очистить популяцию от нежелательных уклоняющихся форм? Причина не только и не столько в постоянном возникновении все новых и новых мутаций. Причина в том, что часто наиболее приспособленными оказываются гетерозиготные генотипы. При скрещивании они постоянно дают расщепление и в их потомстве появляются гомозиготные потомки со сниженной приспособленностью. Это явление получило название сбалансированный полиморфизм.
Половой отбор. У самцов многих видов обнаруживаются явно выраженные вторичные половые признаки, которые на первый взгляд кажутся неадаптивными: хвост павлина, яркие перья райских птиц и попугаев, алые гребни петухов, феерические цвета тропических рыбок, песни птиц и лягушек, и т.п. Многие из этих особенностей осложняют жизнь их носителей, делают их легко заметными для хищников. Казалось бы, эти признаки не дают никаких преимуществ их носителям в борьбе за существование, и тем не менее они очень широко распространены в природе. Какую роль в их возникновении и распространении сыграл естественный отбор?
Известно, что выживание организмов является важным, но не единственным компонентом естественного отбора. Другим важнейшим компонентом является привлекательность для особей противоположного пола. Ч.Дарвин назвал это явление половым отбором. Впервые он упомянул эту форму отбора в «Происхождении видов», а затем подробно проанализировал ее в книге «Происхождение человека и половой отбор». Он считал, что «эта форма отбора определяется не борьбой за существование в отношениях органических существ между собою или с внешними условиями, но соперничеством между особями одного пола, обычно самцами, за обладание особями другого пола».
Половой отбор - это естественный отбор на успех в размножении. Признаки, которые снижают жизнеспособность их носителей, могут возникать и распространяться, если преимущества, которые они дают в успехе размножения значительно выше, чем их недостатки для выживания. Самец, который живет недолго, но нравится самкам и поэтому производит много потомков, имеет гораздо более высокую совокупную приспособленность, чем тот, что живет долго, но оставляет мало потомков. У многих видов животных подавляющее большинство самцов вовсе не участвует в размножении. В каждом поколении между самцами возникает жесточайшая конкуренция за самок. Эта конкуренция может быть прямой, и проявляться в виде борьбы за территории или турнирных боев. Она может происходить и в косвенной форме и быть обусловленной выбором самок. В тех случаях, когда самки выбирают самцов, конкуренция самцов проявляется в демонстрации их яркого внешнего вида или сложного поведения ухаживания. Самки выбирают тех самцов, которые им больше всего нравятся. Как правило, это наиболее яркие самцы. Но почему самкам нравятся яркие самцы?
Приспособленность самки зависит о того, насколько объективно она способна оценить потенциальную приспособленность будущего отца своих детей. Она должна выбрать такого самца, сыновья которого будут обладать высокой приспособленностью и привлекательностью для самок.
Было предложено две основные гипотезы о механизмах полового отбора.
Согласно гипотезе «хороших генов» самка «рассуждает» следующим образом: «Если этот самец, несмотря на его яркое оперение и длинный хвост, каким-то образом умудрился не погибнуть в лапах хищника и дожить до половой зрелости, то, следовательно, он обладает хорошими генами, которые позволили ему это сделать. Значит, его стоит выбрать в качестве отца для своих детей: он передаст им свои хорошие гены». Яркость окраски перьев и гребней у птиц, чешуи и хвоста у рыб является надежной характеристикой здоровья. Так, на рыбке корюшке было показано, что чем ярче окраска самца, тем более он привлекателен для самок и тем менее он заражен паразитами. Яркость гребня у петухов напрямую зависит от содержания тестостерона в крови и от степени зараженности паразитами. Чем выше уровень тестостерона, тем ярче гребень, чем больше паразитов, тем он бледнее. Известно, также, что тестостерон угнетает иммунную систему. Следовательно, самцы с ярким гребнем обладают такой эффективной иммунной системой, которая обеспечивает их устойчивость к паразитам, несмотря на высокий уровень тестостерона. Выбирая ярких самцов, самки выбирают хорошие гены для своих потомков.
Согласно гипотезе «привлекательных сыновей» логика выбора самок несколько иная. Если яркие самцы, по каким бы то ни было причинам, являются привлекательными для самок, то стоит выбирать яркого отца для своих будущих сыновей, потому что его сыновья унаследуют гены яркой окраски и будут привлекательными для самок в следующем поколении. Таким образом, возникает положительная обратная связь, которая приводит к тому, что из поколения в поколение яркость оперения самцов все более и более усиливается. Процесс идет по нарастающей до тех пор, пока не достигнет предела жизнеспособности. Представим себе ситуацию, когда самки выбирают самцов с более длинным хвостом. Длиннохвостые производят больше потомков, чем самцы с короткими и средними хвостами. Из поколения в поколение длина хвоста увеличивается, потому что самки выбирают самцов не с определенным размером хвоста, но с большим, чем в среднем размером. В конце концов, хвост достигает такой длины, когда его вред для жизнеспособности самца уравновешивается его привлекательностью в глазах самок.
Объясняя эти гипотезы, мы старались понять логику действия самок птиц. Может создаться впечатление, что мы слишком много от них ожидаем, что такие сложные расчеты приспособленности им вряд ли доступны. На самом деле, в выборе самцов самки не более и не менее логичны, чем во всем остальном их поведении. Когда животное чувствует жажду, оно не рассуждает, что ему следует попить воды, для того чтобы восстановить водно-солевой баланс в организме – оно идет на водопой, потому что чувствует жажду. Когда рабочая пчела жалит хищника, напавшего на улей, она не вычисляет, насколько этим своим самопожертвованием она повышает совокупную приспособленность своих сестер – она следует инстинкту. Точно так же и самки, выбирая ярких самцов, следуют своим инстинктами – им нравятся яркие хвосты. Все те, кому инстинкт подсказывал иное поведение, все они не оставили потомства. Таким образом, мы обсуждали не логику самок, а логику борьбы за существование и естественного отбора – слепого и автоматического процесса, который, действуя постоянно из поколения в поколение, сформировал все то удивительное разнообразие форм, окрасок и инстинктов, которое мы наблюдаем в мире живой природы.
Похожие работы
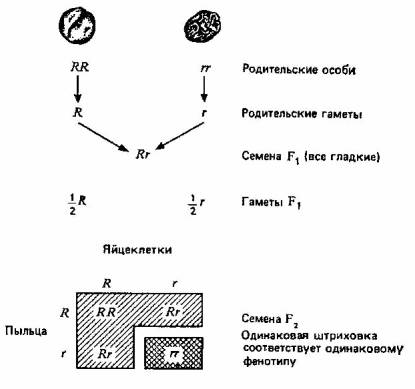
... некими частицами, передаваемыми от родителей потомкам. Теперь называем эти частицы генами. Идея корпускулярной наследственности имеет огромное значение для понимания того, каким образом естественный отбор действует в популяциях. Эволюцию можно рассматривать как изменения любого свойства данной популяции с течением времени. В некоем общефилософском смысле в этом и заключается суть эволюции. ...





... их к изменившимся условиям, стремились бы сохраниться, и естественный отбор имел бы полный простор для своего улучшающего действия. 1. ЕСТЕСТВЕННЫЙ ОТБОР КАК ЭЛЕМЕНТАРНЫЙ ЭВОЛЮЦИОННЫЙ ФАКТОР Сохранение благоприятных индивидуальных различий и изменений и уничтожение вредных я назвал естественным отбором или переживанием наиболее приспособленных Ч. Дарвин В современном понимании ...
... сохранения и накопления малых наследственных изменений , каждое из которых выгодно для сохраняемого существа. Обстоятельства, благоприятствующие образованию новых форм посредством естественного отбора. Значительная часть изменчивости, разумеется, и индивидуальные различия, будут, очевидно, являться обстоятельством благоприятствующим. Большое количество особей, увеличивая шансы появления в ...
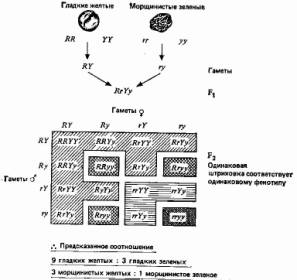

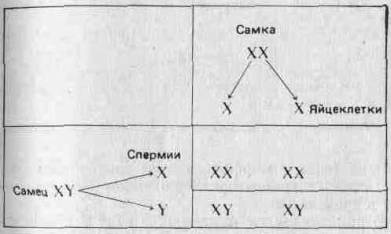
... , а поэтому они играют более важную роль в эволюции. Принципиально важное значение имеет тот факт, что эти мутации случайны, иными словами, они не направленны. 3. Центральная догма и принцип Вейсмана принимаются. 4. Эволюция осуществляется путем изменения частот генов. 5. Эти изменения могут происходить в результате мутаций, поступления генов в популяцию и оттока их из нее, случайного дрейфа и ...
0 комментариев