Ионные механизмы потенциала действия
План
1. Ионные механизмы потенциала действия
1.1 Измерение мембранных токов
1.2 Натриевые и калиевые токи
2. Роста потенциала 2.1 Какое количество ионов входит в клетку и выходит из нее во время потенциала действия? 2.2 Положительная и отрицательная обратная связь во время изменений проводимости 2.3 Измерения проводимостиВывод
1. Ионные механизмы потенциала действия
1.1 Измерение мембранных токов
Количественное описание механизмов, участвующих в генерации потенциала действия, стало возможным благодаря методу измерения мембранных токов в условии фиксации потенциала. Этот метод позволяет определить, какой вклад вносят ионы того или иного типа в мембранный ток, а также вычислить величину и временной ход изменений соответствующих ионных проводимостей. Активация натриевой проводимости носит кратковременный характер, за ней следует инактивация. Увеличение калиевой проводимости продолжается до тех пор, пока не закончится деполяризация. Именно зависимость натриевой и калиевой проводимостей от мембранного потенциала и их попеременная активация качественно определяют как амплитуду, так и временной ход потенциала действия, равно как и другие мембранные характеристики, включая порог и рефрактерный период.
Исследование проводимостей одиночных калиевых и натриевых каналов во время потенциала действия проводились в условиях фиксации потенциала участка мембраны. Наблюдаемые при этом принципы работы отдельных каналов соответствуют результатам, полученным ранее в экспериментах с фиксацией потенциала целой клетки: при деполяризации вероятность открытия натриевых и калиевых каналов возрастает. Возрастание вероятности происходит с тем же временным ходом, что и соответствующие токи в условиях фиксации потенциала. Так, натриевые каналы наиболее часто открываются в начале деполяризующего импульса и вероятность таких открытий падает по мере развития натриевой инактивации.
В генерации потенциала действия могут принимать участие и другие катионные каналы. В некоторых клетках фаза роста потенциала действия определяется активацией кальциевых каналов, а реполяризация происходит благодаря активации различных типов калиевых каналов.
1.2 Натриевые и калиевые токи
Потенциал покоя зависит главным образом от разности концентраций калия (как было предложено Бернштейном в 1902 году), и, в меньшей степени, от концентраций натрия и хлора. Одновременно с созданием Бернштейном теории потенциала покоя, Овертонсделал важное открытие: он показал, что для генерации нервом потенциала действия необходим натрий, и сделал робкое предположение о том, что основой потенциала действия является вход ионов натрия в клетку. Развитие эта гипотеза получила благодаря опытам на аксоне кальмара.
В 1939 году Ходжкин и Хаксли показали, что на пике потенциала действия происходит кратковременная смена знака («овершут») потенциала на внутренней стороне мембраны в область положительных значений. Это наблюдение подтвердило участие натрия в потенциале действия, поскольку вход натрия в клетку происходит и при положительных значениях мембранного потенциала вплоть до натриевого равновесного потенциала ENa. Спустя десятилетие Ходжкин и Катц добились уменьшения явления овершута путем снижения внеклеточной концентрации натрия, а следовательно, и ЕNa. Они пришли к выводу, что потенциал действия есть результат значительного, хотя и кратковременного, увеличения натриевой проводимости мембраны. Сегодня известно, что это увеличение происходит за счет открытия огромного числа потенциалзависимых натриевых каналов.
Чем объясняется фаза спада потенциала действия? Можно предположить, что спад происходит просто в результате закрытия натриевых каналов. Это, действительно, один из факторов. Однако, если бы это был единственный механизм, то возврат мембранного потенциала к уровню потенциала покоя в большинстве клеток происходил бы гораздо медленнее, чем в действительности. Причина этого в том, что общая проводимость мембраны в покое достаточно мала, и выход накопленных в результате потенциала действия положительных ионов из клетки занял бы несколько миллисекунд, а то и десятки миллисекунд. В действительности, возврат к уровню потенциала покоя происходит гораздо быстрее благодаря значительному увеличению другой мембранной проводимости, а именно, за счет открытия калиевых каналов. С той же быстротой, с которой мембранный потенциал деполяризовался в направлении ENa, он гиперполяризуется на фазе спада потенциала действия в направлении ЕK. Увеличение калиевой проводимости может продолжаться до нескольких миллисекунд, что приводит во многих клетках не только к реполяризации, но и к гиперполяризации мембраны до значений более отрицательных, чем потенциал покоя.
2. Рост потенциала
2.1 Какое количество ионов входит в клетку и выходит из нее во время потенциала действия?
Если учесть, что на фазе роста потенциала действия в клетку входит большое количество натрия и большое количество калия ее покидает на фазе спада, то становится очевидным, что концентрации этих ионов в цитоплазме должны измениться. Величину этих изменений можно определить либо экспериментально, либо путем расчетов.
Вычисления соотношения между мембранным потенциалом и трансмембранных концентраций. При потенциале -67 мВ на внутренней поверхности мембраны находится приблизительно 4 · 1011 отрицательных зарядов на см2. На пике потенциала действия (+40 мВ) вместо этого отрицательного заряда внутри клетки накапливается около 2,4 · 1011 положительных зарядов, что происходит в результате входа в клетку 6,4 · 1011 ионов натрия на см2. Это соответствует приблизительно 10–12 моль/см2. Экспериментальные измерения входа радиоактивного натрия и выхода радиоактивного калия дали значения между 3 · 10–12 и 4 · 10–12 моль/см2. Различие между теоретически предсказанным и экспериментальным значениями объясняются главным образом тем, что при расчетах не принималось во внимание то, что вход натрия и выход калия частично перекрываются во времени. Таким образом, количество входящего натрия превышает значение, необходимое для деполяризации мембраны до пикового уровня потенциала действия, поскольку выход калия начинается до момента достижения пика.
Каково влияние входа натрия на его внутриклеточную концентрацию? Сегмент аксона кальмара длиной 1 см и диаметром 1 мм имеет площадь поверхности около 0,31 см2, поэтому при скорости входа натрия 3,5 · 10–12 приведет к накоплению приблизительно 10–12 M натрия. Объем этого отрезка аксона составляет 7,85 · 10–12 л, в нем содержится (при плотности 50 ммоль/л) 4 · 10–7 M натрия, поэтому изменение концентрации натрия в результате потенциала действия очень мало и составляет 0,0000025. Выход калия производит сравнимый эффект и изменяет внутриклеточную концентрацию калия в 0,000003 раз.
Потенциал действия в более тонких отростках нейрона может привести к более значительным изменениям внутриклеточных концентраций натрия и калия, чем в гигантском аксоне кальмара. Так, нервное окончание диаметром 1 мкм и длиной 100 мкм имеет площадь поверхности 3 · 10–6 см2 и объем 8· 10–14 л. Во время потенциала действия вход натрия 3 · 10–6 моль/см2 производит увеличение количества натрия внутри терминалы на 10–17 М. При внутриклеточной концентрации 20 мМ в терминалы содержится приблизительно 1,5·10–15 М, так что увеличение концентрации натрия в результате одного потенциала действия составляет 0,7 %. Пачка из 50 импульсов теоретически приведет к увеличению уровня натрия на 35 % и сходному снижению уровня калия. Вход натрия увеличивает активность натрий-калиевого обменника так, что концентрации быстро возвращаются к изначальному уровню.
2.2 Положительная и отрицательная обратная связь во время изменений проводимостиГлавным свойством натриевой и калиевой проводимостей, определяющим характер токов во время потенциала действия, является их потенциалзависимость: вероятность открытия ионных каналов увеличивается с деполяризацией мембранного потенциала. Деполяризация увеличивает натриевую проводимость, а также, с некоторой задержкой, калиевую. Воздействие деполяризации на натриевую проводимость носит регенеративный характер: сначала небольшая деполяризация увеличивает количество открытых каналов; ионы натрия, входящие в клетку по направлению своего электрохимического градиента, производят дальнейшую деполяризацию мембраны, открытие большего количества каналов, что влечет за собой вход еще большего числа ионов, и так далее. Такой самоусиливающийся процесс характеризуется положительной обратной связью. Потенциал--зависимость калиевых каналов, наоборот, характеризуется наличием отрицательной обратной связи. При деполяризации количество открытых калиевых каналов возрастает, и ионы калия входят в клетку в направлении электрохимического градиента. Однако, вход калия не усиливает деполяризацию, а ведет к реполяризации и возвращению калиевой проводимости в состояние покоя.
2.3 Измерения проводимостиТо, что до сих пор обсуждалось, было предложено Ходжкином, Хаксли и Катцем и детально разработано Ходжкином и Хаксли, которые поставили элегантные опыты на гигантском аксоне кальмара. Они экспериментально показали, что при потенциале действия происходят изменения калиевой и натриевой проводимостей, и что последовательность этих изменений в точности определяет временной ход и величину потенциала действия.
На основании каких опытов был сделан этот вывод? На первый взгляд, измерение натриевой (gNa) и калиевой (gк) проводимостей кажется простой задачей. Нужно только измерить величину тока (I), протекающего через мембрану при том или ином потенциале (Vm), поскольку:
![]()
Однако, прежде необходимо решить две проблемы. Во-первых, ток, протекающий через мембрану, приведет к изменению мембранного потенциала; это, в свою очередь, вызовет изменение проводимостей. Решение было найдено путем разработки метода, позволяющего быстро устанавливать заданный мембранный потенциал и поддерживать его на постоянном уровне, при этом измеряя величину и временной ход мембранного тока. Поскольку потенциал фиксирован в течение всего времени эксперимента, ток будет аккуратно отражать изменения проводимости мембраны. Вторая проблема состоит в том, чтобы разделить различные компоненты мембранного тока для того, чтобы измерить их индивидуальные характеристики. Для преодоления этого затруднения применялись различные методы, в том числе замена натрия на ион, не проникающий через мембрану, а позднее использовались селективные токсины и яды.
Вывод
Потенциал действия является результатом быстрого и значительного увеличения натриевой проводимости мембраны. Вход большого количества ионов хлора и аккумуляция положительного заряда на внутренней поверхности мембраны клетки сдвигает мембранный потенциал в направлении ENa. Реполяризация мембраны происходит в результате последующего увеличения калиевой проводимости и уменьшения внутриклеточного положительного заряда из-за выхода ионов калия из клетки. Мембранный потенциал при этом вновь приближается к Ек. Такое объяснение механизма генерации потенциала действия ведет к пониманию принципов распространения импульса.
Литература
1. Реутов В.П. и др. Проблема оксида азота в биологии и медицине и принцип цикличности.
2. Методология биологии: новые идеи (синергетика, семиотика, коэволюция). Ред. Баксанский О.Е.
3. Новиков Г.Г. Рост и энергетика развития костистых рыб в раннем онтогенезе.
Похожие работы
е реобазы и хронаксии ввёл в физиологию Л. Лапик в 1909, определяя зависимость между силой тока и длительностью его действия при изучении наименьшего (порогового) эффекта возбудимых тканей. Реобаза, как и хронаксия, даёт представление о возбудимости тканей и органов по порогу силы и длительности действия раздражения. Реобаза соответствует порогу раздражения и выражается в вольтах или миллиамперах. ...
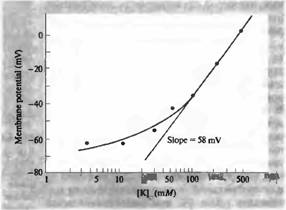
... мембранной проницаемости для натрия, экспериментальные данные расходятся с прямой линий, особенно при низких концентрациях калия. Из уравнения постоянного поля следует один важный принцип. Мембранный потенциал зависит от проводимости (или проницаемости) мембраны для важнейших ионов, а также от их равновесных потенциалов. В реальных клетках проводимости для калия и хлора сравнительно· высоки, ...
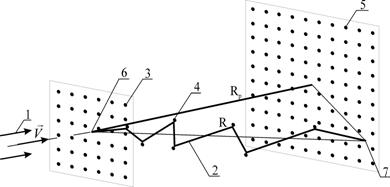
... является то, что рабочий стол 6 с обрабатываемыми образцами 5 размещается внутри данного устройства. Разрабатываемое оборудование позволит осуществлять имплантацию ионов азота с энергией 1 – 10 кэВ ( Дж) в металлы и сплавы, модифицируя их свойства в нужном направлении. Заключение Несмотря на большое количество исследований в области ионной имплантации, остаётся ещё множество вопросов, ...
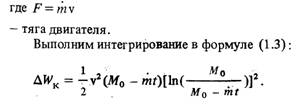
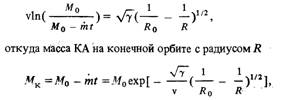
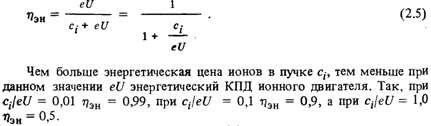
... ионные источники Как уже говорилось, в контактных ионных источниках образование ионов происходит в результате поверхностной ионизации. В ионных электрических ракетных двигателях применяются контактные ионные источники с пористым ионизатором. Пористые ионизаторы изготавливаются различными способами, например, тонкие каналы-поры могут быть пробиты лазерным или электрическим лучом. В качестве ...
0 комментариев