Навигация
Бактерии, участвующие в маслянокислом брожении
73996
знаков
10
таблиц
0
изображений
1.2 Бактерии, участвующие в маслянокислом брожении
К клостридиям относят большое количество видов бактерий, число которых постоянно возрастает. Это один из самых крупных родов среди эубактерий. Принадлежность к роду определяется на основании только трех признаков:1)способности образовывать эндоспоры 2) облигатно анаэробного характера энергетического метаболизма 3) неспособности осуществлять диссимиляционное восстановление сульфата. Отсюда понятно, что эта группа эубактерий чрезвычайно гетерогенна, о чем, в частности, свидетельствует интервал значений ГЦ - оснований ДНК, молярное содержание которых с учетом описанных новых видов занимает область от 21 до 57%.
Из этого также можно сделать вывод, что организмы, объединенные в род Clostridium, нельзя рассматривать как эволюционно однотипные. Последующая характеристика их метаболических особенностей дает достаточно четкое представление об этом. Изучение эубактерий, относимых к клостридиям, наоборот, указывает на раннее расхождение видов рода в процессе эволюции. За исключением C. coccoides, вегетативные клетки бактерий из рода Clostridium имеют форму прямых или слегка изогнутых палочек с закругленными концами. Большинство видов грамположительные, подвижные. Движение осуществляется с помощью перитрихиально расположенных жгутиков. По мере старения в процессе цикла развития клетки теряют подвижность, накапливают гранулезу (запасное вещество типа крахмала) и переходят к спорообразованию. Образующиеся споры овальной или сферической формы. Диаметр их, как правило, превышает диаметр вегетативной клетки, поэтому, если, формирующаяся спора расположена в центре клетки, последние меняют форму, становясь веретеновидными; если же споры образуются у одного из клеточных концов, клетки приобретают форму барабанных палочек.
Клостридии - облигатные анаэробы. Однако спектр их чувствительности к молекулярному кислороду достаточно широк, что связано с обнаружением в клетках большинства клостридиев супероксиддисмутазы и с другими приспособлениями на уровне клеточных популяций, помогающими нейтрализовать токсические эффекты О2 и его производных. Именно при работе с клостридиями Л. Пастер в 1961г. открыл форму жизни без кислорода (Емцев, Мишустин, 2006).
В зависимости от вида сбраживаемого субстрата выделяют несколько физиологических групп клостридиев: сахаролитические клостридии, использующие в качестве субстратов брожения вещества углеводной природы (моносахара, крахмал, клетчатка); протеолитические клостридии, субстратами брожения которых являются белки, пептиды, аминокислоты пуринолитические клостридии, специфически приспособленные к сбраживанию гетероциклических соединений (пурины и пиримидины). Среди них есть виды, обладающие довольно широкими возможностями (субстратами брожения служат как углеводы, так и белки), и узкоспециализированные виды, способные использовать в качестве источника энергии и углерода какое-либо или очень небольшое число соединений.
Субстратом брожения сахаролитических клостридий является такие моносахара, как глюкоза, фруктоза, ксилоза и др. Некоторые виды могут использовать крахмал, целлюлозу, пектин, хитин, предварительно гидролизуемые соответствующими экзоферментами. Типичными представителями сахаролитических клостридий, осуществляющих классическое маслянокислое брожение, являются C. butyricum и C. pasteurianum.
Таблица 1. Основные продукты брожения некоторых сахаролитических клостридий, не образующих масляной кислоты.
| Организм | Основные продукты брожения |
| C. sphenoides , C. glycolicum | этанол, уксусная кислота, СО2, Н2 |
| C. cellobioparum | |
| этанол, уксусная, муравьиная, молочная кислоты; СО2, Н2 | |
| C. clostridioforme | |
| этанол, муравьиная, уксусная, молочная кислоты; СО2, Н2 | |
| C. oroticum | этанол; уксусная, молочная, муравьиная кислоты; СО2 |
| C. coccoides | |
| янтарная, уксусная кислоты | |
| C durum | этанол, пропанол; муравьиная, уксусная, молочная кислоты |
| C nexile | |
| этанол; муравьиная, уксусная, молочная, янтарная кислоты; Н2 | |
| С. quercicolum | уксусная, пропионовая кислоты, Н2 |
| C. ramosum | |
| муравьиная, уксусная, молочная, янтарная кислоты | |
| C. aceticum, C. thermaceticum, C. formicaceticum, C. spiroforme | уксусная кислота |
К протеолитическим относятся клостридии, имеющие активные протеолитические ферменты и поэтому способные использовать в качестве субстратов белки и пептиды, гидролизуя их до аминокислот и подвергая затем последние сбраживанию. В эту группу входят C. putrificum, C. histoliticum, C. sporogenes и другие сапрофитные виды. Близки к этим видам и некоторые патогенные формы: C. botulinum – продуцент ботулина – экзотоксина, являющегося одним из самых сильных биологических ядов; C. tetani – столбнячная палочка, образующая в организме человека столбнячный токсин. К протеолитическим клостридиям примыкают виды, использующие в качестве источника углерода и энергии ограниченное число свободных аминокислот. Например, C. cochlearium растет только на среде с глутаминовой кислотой, глутамином и гистидином; C. sticklandii может сбраживать лизин, аргинин, фенилаланин, серин, а C. propionicum – треонин, аланин, серин, цистеин.
С жизнедеятельностью клостридий связаны различные процессы, протекающие в природе: разложение (гниение) азотсодержащих соединений (белков, нуклеиновых кислот) в анаэробных условиях; анаэробное разложение растительных материалов, таких как клетчатка, хитин. Некоторые сахаролитические клостридии могут использовать в качестве субстрата брожения пектиновые вещества, составляющие покровы растительных клеток. Пектин — полимер метил-D-галактуроновой кислоты. Последняя имеет сложное строение и при воздействии на нее пектиновыми ферментами гидролизуется на ряд сахаров, кислот и метиловый спирт. Клостридии, принадлежащие к виду C felsineum, содержат активную пектиназу и могут, поэтому получать энергию, осуществляя маслянокислое брожение пектиновых веществ. Этот вид играет важную роль в процессе мацерации волокон при мочке льна.
Еще в конце прошлого века было обнаружено, что некоторые клостридии патогенны, т. е. вызывают заболевания человека и животных. В основе патогенности клостридиев лежит их способность синтезировать и выделять из клетки высокоэффективные токсины.
К возбудителям маслянокислого брожения относится в первую очередь вид Clostridium saccharobutyricum (Granulobacter saccharobutyricum). Клетки его цилиндрической формы, длиной от 4-5 до 7-12 мкм и толщиной от 0,5 до 1-1,5мкм, подвижные, спорообразующие. Перед образованием спор в них накапливается гранулеза. Спора появляется чаще в центре клетки, последняя возле споры расширяется (клостридий). Иногда спора располагается на конце клетки, форма которой приобретает вид ракетки (плектридий). Споры довольно устойчивые, они могут выносить непродолжительное кипячение в течение 1-2 мин.
Clostridium saccharobutyricum - облигатный анаэроб, с оптимальной температурой развития 30-40оС, сбраживает многие углеводы и близкие к ним соединения с обильным выделением водорода и углекислоты при одновременном накоплении масляной кислоты. Последняя этими бактериями не потребляется. С.Н. Виноградским описан возбудитель маслянокислого брожения Clostridium pasteurianum, способный усваивать атмосферный азот. По многим признакам он сходен с предыдущим видом, но отличается от него биохимическими особенностям - он не сбраживает крахмал. К маслянокислым бактериям относится также вид Clostridium butyricum и некоторые другие представители. По отношению к источникам азота Маслянокислые бактерии весьма неприхотливы: они усваивают белковый, аминокислотный и аммонийный азот, а некоторые используют азот воздуха.
Маслянокислые бактерии являются также возбудителями ацетонобутилового брожения, анаэробного брожения клетчатки и брожения пектиновых веществ.
Ацетонобутиловое брожение представляет собой превращения углеводов бактериями с образованием ацетона и бутилового спирта. При этом брожении, кроме ацетона и бутилового спирта, вырабатываются также масляная и уксусная кислоты, водород и углекислый газ. Суммарно данное брожение можно выразить следующим уравнением:
12 С6Н12О6 → СН3СН2СН2СН2ОН + 4СН3СОСН3 + СН3СН2ОН + СН3СН2СН2СООН + 18Н2 + 28СО2 + 2Н2О + Х кДж
Механизм ацетонобутилового брожения
Химизм ацетонобутилового брожения еще недостаточно выяснен. В последнее время для изучения его используют метод меченых атомов. В бродящую среду вводят промежуточные продукты брожения (уксусную, ацетоуксусную и масляную кислоты), содержащие углерод С13, а затем определяют наличие этого углерода в конечных продуктах. Этим способом установлено, что при добавлении уксусной кислоты с С13 около 50% всего меченого углерода содержится в бутиловом спирте, около 15%- в ацетоне, до 18% - углекислом газе. При добавлении масляной кислоты, содержащей С13, около 85% меченого углерода переходит в бутиловый спирт. Он найден также в этиловом спирте, в уксусной и масляной кислотах. Исходя из этих данных, считают, что образование бутилового спирта идет главным образом через масляную кислоту. Кроме того, полагают, что ацетон образуется через масляную кислоту.
В химизме ацетонобутилового брожения имеется много общего с маслянокислым, а частности, первые стадии до образования ацетольдоля протекают подобно маслянокислому брожению. Далее между двумя молекулами ацетальдоля протекает окислительно – восстановительная: оксимасляная кислота и оксибутиловый спирт по следующему уравнению: реакция, в результате которой образуются два соединения: β - оксимасляная кислота и β - оксибутиловый спирт по следующему уравнению:
2СН3СНОСН2СНО + Н2О → СН3СНОНСН2СООН + СН3СНОНСН2СН2ОН
β - Оксимасляная кислота далее отщепляет водород и переходит в ацетоуксусную кислоту, которая затем распадается на ацетон и углекислый газ:
СН3СНОНСН2СООН → СН3СОСН2СООН → СН3СОСН3 + СОв –
оксибутиловый спирт подвергается восстановлению до n - бутилового спирта согласно уравнению:
CН3СНОНСН2СН2ОН + Н2 → СН3СН2СН2СН2ОН + Н2О
Часть уксусного альдегида, по-видимому, также вовлекается в окислительно-восстановительную реакцию с образованием уксусной кислоты и этилового спирта:
СН3СНО + СН3СНО + Н2О →СН3СООН + СН3СН2ОН
В ацетонобутиловом брожении наблюдаются две фазы: первая - кислотная, во время которой усиленно размножаются бактерии, а в среде накапливаются масляная и уксусная кислоты, и вторая - ацетонобутиловая, в ней уменьшается кислотность и происходит усиленное накопление ацетона, бутилового и этилового спиртов. В зависимости от условий брожения, т. е. подавляя какую – либо фазу брожения, можно получить усиленное накопление тех или иных продуктов. Следовательно, ацетонобутиловое брожение отличается от маслянокислого: при маслянокислом брожении накапливающиеся кислоты постепенно замедляют процесс кислотообразования и даже останавливают его, а при ацетонобутиловом – образовавшиеся кислоты потребляются бактериями и превращаются в другие вещества.
Первый возбудитель ацетонобутилового брожения выделен в 1911 г. Бейеринком. Он получил название Granulobacter butyricum. Более активным возбудителем этого брожения явился вид Clostridium acetobutyricum. Клетки его по морфологическим признакам сходны с маслянокислыми бактериями. Это подвижные, грамположительные, палочки со спорами, развивающиеся в анаэробных условиях. Спорообразование - клостридиальное. В клетках накапливается гранулеза. По биохимическим свойствам данный вид сильно отличается от маслянокислых бактерий. Начиная с 1923 г. ацетонобутиловые бактерии используются в промышленных масштабах для получения ацетона и бутилового спирта из крахмалистого сырья и мелассы.
Среди возбудителей маслянокислого брожения встречаются такие культуры, которые способны вызывать брожение клетчатки. Способность анаэробных микроорганизмов разрушать клетчатку впервые была установлена в 1875г. Л. Поповым, а сущность этого процесса раскрыта в 1889- 1902 гг. В. Л. Омелянским, который выделил две разновидности целлюлозоразрушающих бактерий. Одна из них вызывает водородное брожение клетчатки Bac. cellulosae hydrogenicus ,а вторая – метановое Bac. cellulosae metanicus. У этих бактерий морфологических различий почти не наблюдается. Молодые формы - необычайно тонкие, прямые, иногда слегка изогнутые палочки, толщиной от 0,3 до 0,5 мкм и длиной от 4 до 8 мкм. По мере развития палочки удлиняются и достигают 10-15 мкм, затем на их конце их образуются споры. При этом клетки приобретают форму барабанной палочки. Со временем спора в виде круглого тельца освобождается от клетки. Одним из существенных морфологических различий между водородными и метановыми бактериями является разная способность к спор к прорастанию. Споры возбудителя метанового брожения прорастают намного быстрее спор водородного брожения.
Химизм этого брожения сложен, так как целлюлозоразлагающие бактерии сначала переводят углеводы в растворимые сахара, которые затем частично сбраживают главным образом до масляной и уксусной кислот. Из других продуктов эти бактерии продуцируют спирт, муравьиную и молочную кислоты, а также выделяют газы - водород или метан (в зависимости от культуры) и углекислый газ.
При водородном брожении газообразные вещества - водород и углекислота - составляют примерно 1/3 массы разложившейся клетчатки. При метановом брожении газообразных веществ (метана, углекислоты) получается больше, они составляют примерно половину массы разложившейся клетчатки.
Возбудители водородного и метанового брожения наряду с клетчаткой способны сбраживать соли масляной, молочной, муравьиной и других кислот, накапливающиеся при брожении клетчатки. В результате такого процесса в анаэробных условиях достигается полная минерализация клетчатки - наиболее сложного и устойчивого полисахарида. В последнее время установлено, что возбудители маслянокислого брожения клетчатки являются активными продуцентами витамина В12. Это свойство их используется для промышленного получения кормового витамина В12.
Пектиновые вещества входят в состав тканей растений. Являясь составной частью клеточной оболочки, они как бы склеивают клетки между собой, образуя межклеточные пластинки. Они находятся в значительном количестве в некоторых органах растений, например, в листьях, в мякоти плодов, ягод, корнеплодов, в кожице цитрусовых и во многих других.
Пектиновые вещества в природе минерализуются микроорганизмами, обладающими ферментами протопектиназой (пектазой) и полигалактуроновой (пектиназой). Фермент протопектиназа расщепляет в пектине связи между метоксилированной полигалактуроновой кислотой и связанным с ней арабаном. Освободившаяся при этом метоксилированная полигалактуроновая кислота (растворимый пектин) под действием пектинэстеразы гидролизуется с образованием метилового спирта и полигалактуроновой кислоты.
Полигалактуроновая кислота подвергается также действию другого фермента - полигалактуроназы микроорганизмов (бактерий и плесневых грибов). Этот фермент гидролизует гликозидные связи между остатками галактуроновой кислоты, не содержащими метоксильных групп. Галактуроновые кислоты наряду с другими соединениями гидролизуются и используются микроорганизмами в качестве источника углерода. В результате гидролиза пектина образуются продукты: галактуроновая кислота, галактоза, арабиноза, ксилоза, уксусная кислота и метиловый спирт. Некоторые из них – моносахариды – используются этими же пектинразлагающими микроорганизмами, так арабиноза сбраживается ими до масляной кислоты и углекислоты, а из галактозы наряду с масляной и угольной кислотами выделяется водород (Емцев, Мишустин, 2006).
Среди пектинразлагающих микроорганизмов известны бактерии вида Clostridium pectinovorum, которые вызывают маслянокислое брожение пектиновых веществ (описаны Бейеринком в 1902 г.). Они представляют собой сравнительно крупные, подвижные, грамположительные палочки длиной 10-15 мкм и шириной 0,8 мкм. В клетках образуются споры, которые располагаются на утолщенных концах. Относятся эти бактерии к облигатным анаэробам. При сбраживании пектиновых веществ они накапливают масляную и уксусную кислоты, водород и углекислый газ. При соответствующих условиях брожения могут вырабатывать, кроме основных продуктов брожения, бутиловый спирт и ацетон.
Возбудителем пектинового брожения является также Clostridium felsineum. Микроорганизм выделен итальянским исследователем Карбонэ в 1916 г. из стеблей конопли. По форме клеток – споровая палочка одиночная или соединенная в короткие цепочки размерами 3/ 5 * 0,3 мкм. Споры располагаются на конце (барабанная палочка), но иногда бывают и в центре клетки. Культура обладает сравнительно высоким температурным оптимумом. Она сбраживает пектиновые вещества с образованием преимущественно уксусной кислоты и совершенно не продуцирует масляной.
Кроме описанных представителей, известны еще и некоторые другие, которые, разлагая пектиновые вещества, сбраживают продукты гидролиза (арабинозу, галактозу и др.) по типу маслянокислого или ацетонобутилового брожения.
Брожение пектиновых веществ нашло применение в хозяйственно-технической деятельности человека, в частности, для получения прядильных волокон из льна, конопли и других волокнистых растений путем мочки их в естественных водоемах.
Для ускорения и улучшения процесса освобождения волокон мочку сырья производят в бассейнах. При этом способе применяют специально выращенные культуры бактерий. Процесс ведут при температуре 26-28оС в течение 5 суток вместо 1,5-2 недель в естественных водоемах.
Анаэробные бактерии рода Clostridium были открыты Л. Пастером в 1861 г. Ученый обнаружил, что некоторые представители данного рода сбраживают углеводы с образованием масляной кислоты. Сейчас среди представителей Clostridium насчитывается более 80 видов бактерий. Они имеют палочковидные клетки, обычно подвижны, передвигаются при помощи перитрихальных жгутиков. У видов рода образуются споры, имеющие овальную или сферическую форму; они терморезистентны. Как правило, споры раздувают клетку. Грамположительные. Облигатные анаэробы (Богданов и др., 1968).
Род включает психрофильные, мезофильные и термофильные виды. Температурный оптимум для роста большинства мезофильных видов лежит в диапазоне 30-40оС. Для термофильных видов температурный оптимум составляет 60-75оС. Оптимальная реакция среды для клостридий - нейтральная или слабощелочная. Хемоорганогетеротрофы. Клостридии сбраживают сахара, многоатомные спирты, аминокислоты, пурины и пиримидины, другие органические соединения. Ряд видов способен к фиксации молекулярного азота атмосферы. Места обитания клостридий - почва, водоемы, а также пищеварительный тракт человека и животных.
Все виды рода объединены в группы в зависимости от способности сбраживать те или иные органические соединения.
Первая группа - сахаролитические виды Clostridium, сбраживающие главным образом растворимые углеводы, крахмал или пектин с образованием бутирата, ацетата, СО2 и Н2. Брожение при участии некоторых микроорганизмов группы приводит к образованию из сахаров дополнительных нейтральных соединений (бутанола, пропанола, ацетона, небольших количеств этанола). В группу входят бактерии, вызывающие маслянокислое и ацетонобутиловое брожения: C.butyricum, C. pasteurianum, C. tyrobutyricum, C. butylicum, C acetobutyricum и др. Возможно, к ней можно отнести и ряд видов – высокоспециализированных агентов анаэробного разрушения целлюлозы, причем главные конечные продукты брожения - этанол, ацетат и сукцинат, СО2 и Н2.
Вторая группа - протеолитические виды, сбраживающие аминокислоты. Обладают сильными протеолитическими свойствами и способны к интенсивному гидролизу белков с последующим сбраживанием аминокислот. Рост микроорганизмов в средах с белком сопровождается образованием аммиака, СО2, Н2, жирных кислот, и большого количества летучих соединений с неприятным запахом. К группе относятся виды: C. sporogenes, C. perfringens, C. histoliticum, C. botulinum и др. Многие представители рода Clostridium, сбраживающие аминокислоты, способны также к сбраживанию углеводов.
Третья группа - виды сбраживающие азотсодержащие циклические соединения - пурины и пиримидины. Пурины (гуанин, гипоксантин, ксантин и др.) под влиянием C. acidi- urici, C. cylindrosporum превращаются в аммиак, ацетат и СО2. C. uracilicum, C. oroticum сбраживают пиримидины, при этом урацил распадается до в- аланина, СО2 и NН3, а оротовая кислота- до уксусной кислоты, СО2 и NН3.
Четвертая группа включает всего один вид - C. kluyveri, сбраживающий смесь этанола с ацетатом до бутирата и капроновой кислоты, а также небольшого количества водорода.
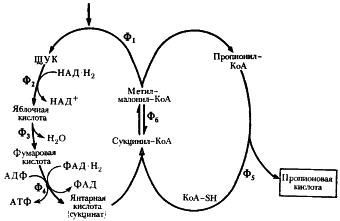
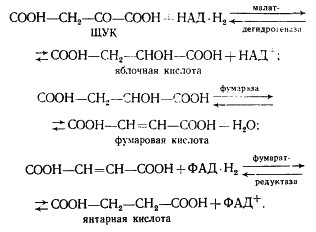
Получение витамина В12 с помощью пропионовокислых бактерий
В настоящее время для получения витамина В12 используют следующие микроорганизмы Prop. freudenreichii ATCC 6207, Prop. shermanii ATCC 13673, Prop. shermanii BKM-103 и их варианты и мутанты. Наибольший интерес представляют разложение клетчатки.
Похожие работы
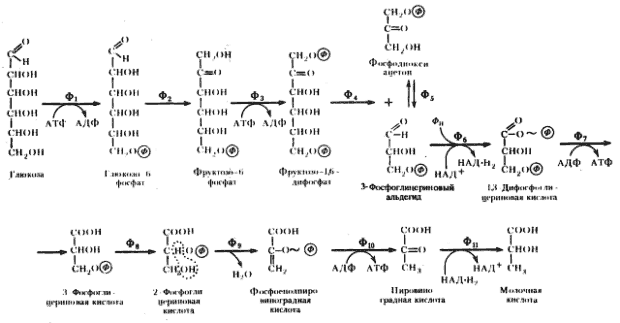
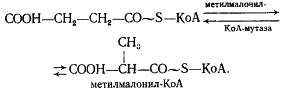
... Тот факт, что в самом "классическом" виде спиртовое брожение проявляется у дрожжей, форм эукариотных, не может, как нам кажется, ставить под сомнение его место в эволюции анаэробных энергетических процессов. ПРОПИОНОВОКИСЛОЕ БРОЖЕНИЕ Из рассмотренных двух типов брожения видно, что ключевым соединением в обоих процессах является пируват, поскольку в конечном итоге специфика брожения определяется ...
... — 0,5 мл. Противостолбнячную сыворотку применяют в неотложных случаях. Действие ее наступает быстро и бывает эффективным (Ассонов, 1989). 1.3 Значение микроорганизмов рода Clostridium в промышленности Роль микроорганизмов в микробиологической, пищевой промышленности, в сельском хозяйстве и других областях трудно переоценить. Особенно важно отметить то, что многие микроорганизмы для ...
... ), от которых отходят длинные цепочки конидий шаровидной формы, в большинстве зеленого или серовато-сине-зеленого реже желто-зеленого или другого цвета. Этот гриб вызывает порчу мясных и молочных продуктов. Высшие несовершенные грибы. Их классификация основана на строении конидиальных спороношений, а также форме конидий, окраске, числе. Цикл развития этих грибов недостаточно изучен, полового ...
... в какой последовательности и как глубоко учащиеся будут изучать материал. По программе Сивоглазова В.И., Сухова Т.С., Козлова Т.А. в книге для учителя «Биология: общие закономерности» тема «Биогеохимическая деятельность микроорганизмов» не рассматривается как самостоятельная на отдельном уроке, а является составной частью других тем. Например, на уроке по теме «Значение прокариот в биоценозах, их ...
0 комментариев