Навигация
1.3 Актиномицеты
Лучистые грибы, или актиномицеты, являются промежуточной группой между бактериями и грибами. Для актиномицетов характерно наличие одноклеточного, лучисто разрастающегося, очень тонкого (около 1 мкм в диаметре) мицелия, на котором развиваются характерные спорообразующие ответвления, дающие артроспоры, или конидии.
Большинство актиномицетов с мицелиальным строением размножаются спорами. Споры могут быть одиночными или собранными в цепочки разной длины и формы, формируются на спороносцах или в спорангиях. Спороносцы - это прямые, волнистые или спиральные образования, располагающиеся на мицелии поочередно, кистями или мутовками. Обычно споры неподвижны, однако у некоторых актиномицетов они имеют жгутики - перитрихальные или монотрихальные, благодаря которым клетки могут передвигаться в водной среде. Поверхность спор гладкая или с разнообразными выростами - шипами или ворсинками.
Большинство актиномицетов - сапрофиты, участвующие в разрушении органических веществ в почве. Некоторые актиномицеты являются патогенными для растений и животных, вызывая актиномикозы. Актиномицеты не имеют истинного ядра, их ядерный материал находится в диффузном состоянии, что приближает их к бактериям. Взрослый мицелий (совокупность гиф) актиномицетов распадается на отдельные клетки, напоминающие палочки и кокки бактерий. С бактериями актиномицетов сближают также размеры клеток и приуроченность главным образом к щелочной среде.
Актиномицеты делятся на порядки, семейства, роды и виды. В основу деления положены характер плодоношения и строение вегетативных органов. Среди актиномицетов, поражающих растения, для фитопатологии имеют значение представители рода Actinomyces, которые на клубнях картофеля и корнеплодах свеклы вызывают заболевание наружных тканей - паршу. На поверхности пораженного органа образуются коростинки, струпьевидные корочки, язвочки и т.п. Возбудителями парши является несколько видов лучистых грибов, отличающихся рядом признаков, в том числе и по типу поражения. Особенно сильно поражаются клубни на щелочных, сильно известкованных почвах и при внесении под картофель свежего неперепревшего навоза.
1.4 Микоплазмы
Микоплазменные организмы имеют клеточное строение, полиморфны, размер их от 50 до 800 нм и более, покрыты двухслойной мембраной. Микоплазменные организмы не имеют ригидной клеточной стенки, не способны синтезировать пептидогликан. Это плеоморфные организмы, размножающиеся почкованием, фрагментацией и бинарным делением. Клетки микоплазм окружены цитоплазматической мембраной, в состав которой входят стерины, в частности эргостерин. Сами микоплазмы данные стерины не синтезируют, а удовлетворяют потребность в указанных веществах, получая их из внешней среды - от других животных организмов, с которыми находятся во взаимосвязи. Ряд микоплазм синтезируют каротиноиды, накапливающиеся в мембране. Микоплазмы содержат два типа нуклеиновых кислот - РНК и ДНК, имеют набор ферментов и обладают самостоятельным обменом веществ. Форма клеток может быть сферической или овальной, палочковидной, дисковидной, встречаются и тонкие нити с тенденцией к образованию разветвленных мицелевидных структур.
Микоплазмы неподвижны. Факультативные анаэробы, хемоорганогетеротрофы. Распространены на растениях и животных, в водоемах, сточных водах, в почве.
Микоплазмы являются возбудителями более 20 болезней растений: желтухи астр, желтой карликовости риса, карликовости овса, столбура пасленовых, позеленения лепестков земляники, желтухи свеклы и других.
1.5 Грибы
Подавляющее большинство болезней (около 80%) вызывается грибами. Грибы относятся к особой группе организмов, характеризующейся своеобразным комплексом признаков. Важнейшей особенностью грибов является гетеротрофный способ питания, обусловленный отсутствием у грибов хлорофилла и других пигментов, имеющихся у автотрофных растений, с помощью которых эти растения способны самостоятельно создавать органические вещества из неорганических. Грибы, лишенные способности к автотрофному питанию, используют готовые органические вещества растительного или животного происхождения. К морфологическим особенностям грибов относятся наличие у них вегетативного тела нитчатого строения и способность к размножению спорами.
Типичным вегетативным телом грибов является грибница, представляющая собой систему ветвящихся гиф - нитей трубчатого строения. Совокупность разрастающихся гиф и их сплетения называют мицелием, или грибницей. В течение длительного времени нарастание мицелия происходит при неограниченном вертикальном росте гиф, в результате чего длина мицелия может достигать многих метров. У некоторых простейший низших грибов нитчатый типичный мицелий отсутствует, и вегетативное тело у них представлено одной клеткой, нередко лишенной оболочки, или так называемого амебоида, обычно располагающегося внутри питающей клетки растения-хозяина. В других случаях у низших грибов одна или комплекс нескольких клеток имеют оболочки и снабжены тонкими, лишенными собственных ядер ответвлениями - ризоидами, органами прикрепления и питания. Характерной особенностью мицелия низших грибов является отсутствие перегородок. У высших грибов мицелий всегда многоклеточный, с большим количеством поперечных перегородок. Клетки такого мицелия содержат от одного до нескольких ядер.
Гифы грибницы чаще бывают бесцветными, прозрачными, но могут быть окрашенными. Пигменты, обуславливающие цвет мицелия, могут откладываться как в клеточных оболочках, так и в содержимом клеток. Часто у нормального нитчатого мицелия имеются: ризоиды - органы прикрепления к субстрату у гиф и спорангиеносцев (мукоровые грибы); аппрессории - присоски, предназначенные для прикрепления ростковых гиф и мицелия к субстрату (мучнисторосяные, ржавчинные грибы); гаустории - органы питания, проникающие внутрь живых клеток растения в виде простых булавовидно вздутых или часто разветвленных специализированных гиф. У мицелия грибов имеются анастомозы - выросты двух соседних гиф одного или разных мицелий, срастающихся друг с другом в форме соединительных мостиков. Через такие анастомозы происходит обмен содержимым (в том числе и ядерным). В результате подобного обмена в связи с наличием гетерокариозиса обеспечивается разнокачественность мицелия. У различных грибов появление анастомозов часто связано с недостатком питания, прорастанием спор.
У многих сапрофитных форм, а иногда у некоторых паразитов мицелий может простираться по поверхности питающего субстрата. Такой тип мицелия называется экзофитным или эпифитным. Большинство фитопатогенных грибов имеет эндофитный мицелий, т.е. располагающийся внутри тканей растения. При этом мицелий может проникать внутрь клеток растения-хозяина и пронизывать их (внутриклеточный мицелий) или располагаться в межклетниках (межклеточный мицелий), проникая в соседние клетки растения при помощи гаусторий. Межклеточный мицелий характерен для паразитных форм. Нередко мицелий может развиваться в межклетниках и пронизывать клетки тканей.
Распространение мицелия по растению или внутри тканей обычно имеет ограниченный характер и захватывает небольшие участки листа, плода или другого органа (местный, или локальный, мицелий). В более редких случаях встречается диффузный мицелий, охватывающий все или почти все органы растения. Диффузное разрастание мицелия обычно бывает у растений, развивающихся из зараженных семян, клубней, луковиц, у которых были заражены корневища или спящие почки. Из них затем вырастают диффузно зараженные побеги. Иногда у головневых, ржавчинных, некоторых пероноспоровых наблюдается многолетнее развитие мицелия в зимующих органах.
У высших грибов (сумчатых и базидиальных) наблюдается особенно обильное развитие мицелия (разрастание и уплотнение разнообразного характера), которое приводит к формированию особых образований. Простейшей формой такого мицелиального образования являются мицелиальные пленки. Они часто встречаются у дереворазрушающих трутовых грибов. При параллельном росте большого количества гиф в одном направлении часто наблюдается образование тяжей и шнуров, у которых отдельные гифы срастаются друг с другом боковыми стенками.
Более сложной формой мицелиального образования являются ризоморфы, характеризующиеся наличием покровной части, которая состоит из уплотненных участков с пигментированными оболочками клеток, утративших внутреннее содержимое, и живых, параллельно расположенных внутренних гиф, содержащих значительные запасы питательных веществ. Они предназначены для охватывания субстрата, для размножения и образования плодовых тел. Более сложными мицеальными образованиями являются склероции - плотные скопления мицелия. Наружные слои, или покровная часть, склероциев выполняет защитную функцию. Она состоит из толстостенных клеток, лишенных внутреннего содержимого. Внутренняя часть склероция состоит из тонкостенных живых клеток, содержащих большое количество запасных питательных веществ - углеводов, жиров. В состав склероция входит от 10 до 30% жировых веществ.
Склероции у грибов являются покоящейся стадией, в которой гриб способен сохраняться в период неблагоприятных условий: засуха, низкие температуры. Многие грибы зимуют в состоянии склероциев, которые в качестве обязательной стадии входят в их цикл развития. После периода покоя склероции прорастают обычно с образованием спороносящих органов или мицелия.
В состав склероция в одних случаях входят только гифы гриба, а в других - склероции построены из измененных тканей питающего растения. Такое образование склероция называется мумификацией. При мумификации с накоплением в грибнице запасных питательных веществ происходит обезвоживание тканей растения. Мумии также являются покоящимися стадиями гриба (плодовая гниль яблок, груш).
Истинных тканей у грибов нет. В подавляющем большинстве случаев образования возникают в результате плотного сплетения грибных гиф. Ложные ткани грибов называются плектенхимой. Различают прозоплектенхимы - ложные ткани, в состав которых входят удлиненные клетки, и параплектенхимы - ткани, состоящие из изодиаметрических клеток. Чаще параплектенхима является защитной покровной тканью, внутреннее содержимое многих склероциев или покровы и внутренняя ткань некоторых плодовых тел представлены прозоплектенхимой. К видоизменениям мицелия относятся также оидии, бластноспоры, хламидоспоры, геммы.
Бластоспоры известны у дрожжей и дрожжеподобных грибов, вегетативная часть которых представлена свободноживущими отдельными клетками. На таких клетках обычно формируются клетки (почки), в свою очередь часто почкующиеся и увеличивающиеся в размерах. После чего они опадают и ведут самостоятельное существование.
Оидии, или артроспоры - цилиндрические, эллипсовидные клетки, формирующиеся при распадении мицелия или отдельных гиф и служащие для размножения. Формирование оодий идет путем образования поперечных перегородок в неклеточном мицелии или дополнительных перегородок в многоклеточном мицелии при последующем распаде всего мицелия или отдельных гиф на отдельные клетки, обособляющиеся друг от друга. У многих грибов оидии входят в цикл развития в качестве обязательной стадии. Хламидоспоры - толстостенные клетки, образующиеся одиночно или группами, часто в цепочках на вегетативном мицелии. Формирование у них идет более сложным путем. У мицелия с большим количеством перегородок отдельные клетки сохраняют свое содержимое, прочие клетки оказываются опустевшими и в последствие разрушаются, освобождая обособившиеся хламидоспоры, которые являются одновременно покоящимися клетками. Эти клетки снабжены плотными утолщенными оболочками. Хламидоспоры при отсутствии достаточной влажности сохраняют жизнеспособность в течение многих лет, но большинство из них при благоприятных условиях способно прорастать сразу же после образования. Обычно хламидоспоры прорастают в органы спороношения, характерные для вида, реже в ростковую гифу, удлиняющуюся затем в мицелий.
Геммы по строению и по биологическим особенностям близки к хламидоспорам. Образование гемм характерно для энтомофторовых, некоторых головневых, многих сумчатых и несовершенных грибов.
У грибов происходит как вегетативное, так и бесполое и половое, с рядом переходных форм между этими двумя способами.
Вегетативное размножение осуществляется обрывками (кусочками) мицелия или его видоизменениями. Небольшие участки гриба, попав в благоприятные условия, дают начало новым гифам, разрастающимся в новый разветвленный мицелий. Бесполое размножение осуществляется при помощи специальных образований, называемых спорами. Споры могут развиваться внутри специальных споровместилищ или на концах особых выростов мицелия - конидиеносцах. У многих низших грибов бесполое размножение происходит при помощи зооспор, снабженных жгутиками и способных к самостоятельному движению в воде. Зооспоры развиваются в зооспорангиях. У наиболее примитивных современных форм грибов зооспорангий формируется прямо из вегетативного тела. При этом все тело особи (амебоид) состоит из одной клетки, которая лишена собственной оболочки и находится внутри живой клетки растения-хозяина. С достижением зрелости амебоид полностью превращается в зооспорангий. При этом амебоид покрывается оболочкой и его содержимое распадается на отдельные одноядерные участки, которые в виде одножгутиковых зооспор выходят наружу и некоторое время плавают в водной среде.
У более высокоорганизованных оомицетов, переходящих на наземный образ жизни, зооспорангии формируются на спороносцах - специализированных образованиях, резко отличающихся от вегетативных гиф по форме, характеру роста и другим признакам. При созревании зооспорангии отрываются от спарангиеносцев и разносятся воздушными потоками или каплями дождя. При прорастании зооспорангия у большинства оомицетов образуются типичные зооспоры с двумя жгутиками. Но у некоторых видов подвижные стадии утрачены, и зооспорангии прорастают с образованием ростковой трубочки, удлиняющейся затем в вегетативный мицелий. Другой формой бесполого размножения является образование спорангиев, формирующихся на спорангиеносцах - простых или сложных ответвлениях мицелия. Вершины таких спорангиеносцев шаровидно вздуваются, в них формируются неподвижные споры с собственными оболочками.
Наиболее совершенным является конидиальное спороношение, характерное для высших грибов - сумчатых, несовершенных, реже базидиальных. Конидии представляют собой споры, образующиеся экзогенно на концах вегетативных гиф или конидиеносцах. Конидии могут быть как одноклеточными, так и многоклеточными.
Половое размножение состоит из слияния мужских и женских половых гамет, в результате чего возникает зигота. Простейшим типом полового процесса является слияние двух подвижных гамет - планогамия. У оомицетов половой процесс называется оогамией. Мужской гаметангий - антеридий - переливает свое содержимое в женский гаметангий - оогоний с одной или несколькими оосферами. В результате такого оплодотворения в оогонии формируется в одних случаях только одна ооспора, в других - несколько спор, которые имеют характер покоящихся спор. У зигомицетов наблюдается половой процесс типа зигогамии, при котором оба гаметангия не отличаются друг от друга и слияние содержимого гаметангиев происходит при их соприкосновении - на границе между ними. Продукт слияния превращается в зигоспору, сохраняющуюся в виде покоящейся споры. После зимовки зигоспора прорастает и образует первичный спорангий с гаплоидными спорами.
Формы полового слияния у сумчатых грибов очень разнообразны. У первичносумчатых грибов половой процесс сходен с зигогамией. При этом зигота после оплодотворения развивается в сумку, внутри которой формируются аскоспоры. У более высокоорганизованных истинных сумчатых грибов женский орган - архикарп состоит из расширенной многоядерной клетки - аскогона - с ножками у основания и удлиненной клетки - трихогины, расположенной в верхней части архикарпа. Мужской гаметангий представлен антеридием с несколькими ядрами. Содержимое антеридия через трихогину переливается в архикарп, в котором происходят плазмогамия и образование дикарионов. После чего наступает размножение дикарионов и от аскогена отходят аскогенные гифы, на концах которых осуществляется последнее деление дикариона и формируется сумка. В сумке сначала происходит кариогамия, затем следует деления диплоидного ядра. Вокруг образовавшихся 8 гаплоидных ядер внутри сумки формируется 8 аскоспор, которые при прорастании дают начало гаплоидному циклу. После окончания полового слияния раздельнополых клеток у высших аскомицетов отдельные части вегетативного мицелия начинают интенсивно разрастаться и обрастать аскогенными гифами.
У базадиальных грибов основным органом размножения является базидия. Это основной орган размножения, несущий на своей поверхности базидиоспоры. У базидиальных грибов в цикле развития преобладает диплоидное состояние (мицелий и построенные из него плодовые тела с базидиями на них). Гаплоидный цикл у базидиальных грибов резко сокращен. Типичные половые органы у базидиальных грибов утрачены, и диплоидизация происходит слиянием ростков вегетативных разнополых гиф или базидиоспор, сидящих на базидии. Диплоидизация идет при участии спермациев, переливающих свое содержимое внутрь особых воспринимающих гиф. Базидия возникает из двухъядерной клетки (дикарион), или при прорастании покоящейся споры, или непосредственно на вегетативном мицелии в плодовом теле. Оба ядра клетки сливаются, и образовавшееся диплоидное ядро делится обычно дважды, с редукцией числа хромосом, в результате чего образуются 4 гаплоидных ядра. К моменту их образования на базидии формируются выросты - стеригмы, в которые переходят ядра. На концах стеригм образуются вздутия, которые увеличиваясь в размерах, превращаются в базидиоспоры.
Сохранение жизнеспособности спор грибов различно в зависимости от видов грибов. Подвижные споры (зооспоры), лишенные оболочки, в природных условиях погибают быстро, этому способствуют прямые солнечные лучи, сухость воздуха, многочисленные враги из числа микроорганизмов, нематод, насекомых. Споры, имеющие оболочку, сохраняют жизнеспособность несколько суток. В течение нескольких месяцев или лет сохраняют жизнеспособность споры, находящиеся в плодовых телах типа пикнид, клейстотециев, перитециев; от 3 до 5 лет и более сохраняют жизнеспособность хламидоспоры, цисты, ооспоры, зигоспоры, имеющие оболочки и большой запас питательных веществ.
Распространяются споры несколькими путями: 1) активно - распространение, обязанное самостоятельному передвижению (зооспоры, передвигающиеся в воде при помощи жгутиков); 2) пассивно - распространение при содействии различных агентов (воздушных течений, воды, насекомых); 3) интраматрикально - распространение внутри тканей хозяина.
Для грибов характерен определенный цикл развития. Циклом развития называется последовательное прохождение различных стадий и спороношений, завершающихся образованием спор. У грибов наблюдается явление плеоморфизма и полиморфизма. Плеоморфизм - свойство организма развивать в онтогенезе несколько типов спороношения, строго следующих одно за другим (ржавчинные грибы). Полиморфизм - способность грибов изменяться в процессе развития, в связи с изменением внешних условий (полиморфные виды). Это явление характерно для сумчатых и несовершенных грибов.
Цикл развития гриба может проходить на одном или нескольких видах растений. В зависимости от этой особенности виды грибов называются однохозяйственными или разнохозяйственными.
Однохозяйственность - способность видов грибов осуществлять полный цикл развития на одном виде питающего растения (возбудители ржавчины малины, льна, мучнисторосяные и др.)
Разнохозяйственность - осуществление полного цикла развития патогена на различных питающих растениях, далеких друг от друга в систематическом отношении (возбудители линейной ржавчины злаков, ржавчины гороха и люцерны и др.)
Для грибов характерна изменчивость их агрессивных, патогенных и других свойств. К изменчивости приводят такие причины, как гибридизация, мутация, изменение условий развития. У патогенных грибов часто наблюдается внутривидовая изменчивость.
Возбудителями ржавчины и головни хлебных злаков являются грибы класса Basidiomycetes. Корневую гниль злаков вызывают грибы из родов Fuzarium, Bipoiarius, Ophiobolus, Pythium, Pectobacterium, Pseudomonas и др. Причиной спорыньи является гриб Claviceps purpurea Fr Tul. класса Ascomycetes. Fusarium nivale (Fr) Ces. класса Deuteromycetes вызывает фузариозную плесень (выпревание озимых), Erysiphe graminis DC. класса Ascomycetes - мучнистую росу злаков. Возбудителем ржавчины гороха является гриб Uromyces pisi Schroet. класса Basidiomycetes, фитофтороза картофеля и томатов - Phytophtora infestans d By класса Oomycetes. Плодовую гниль яблони и груши вызывает Monila fructigena (Fr.) West. класса Deuteromycetes, мучнистою росу яблони - Podosphaera leucotricha Salm. класса Ascomycetes.
Похожие работы


... листьев. Конидии меланкониевых образуются на коротких конидиеносцах в конидиальных ложах, выступающих на поверхность субстрата в виде плоских или выпуклых подушечек. Антракнозы сельскохозяйственных культур характеризуются большой вредоносностью. Из наиболее распространенных болезней этой группы можно назвать антракноз льна, антракноз фасоли и антракноз тыквенных (рис. 2), вызываемые грибами рода ...
... я растением-хозяином в желаемом направлении. Эта работа явилась началом применения агротехнических способов защиты растений от болезней. Итоги работ украинских фитопатологов по борьбе с головней были в 1932 г. обобщены А.И. Боргардтом в работе «Основы построения системы мероприятий по ликвидации головни в зерновом производстве СССР». Внедрение этой системы в сельское хозяйство привело к тому, что ...
... 416 1,5 9,0 50,1 5,0 1,6 12,9 4.2 Общее фитосанитарное состояние посевов сельскохозяйственных культур Челябинской области за 2007–2008 г. По данным фитопатологического анализа семян зерновых культур под посев в 2008 году – 65,6% семян заражено корневыми гнилями. В 2007 году – 67,2%. Все проверенные партии семян были заражены. Как и прежде, ...
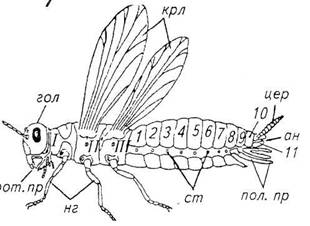
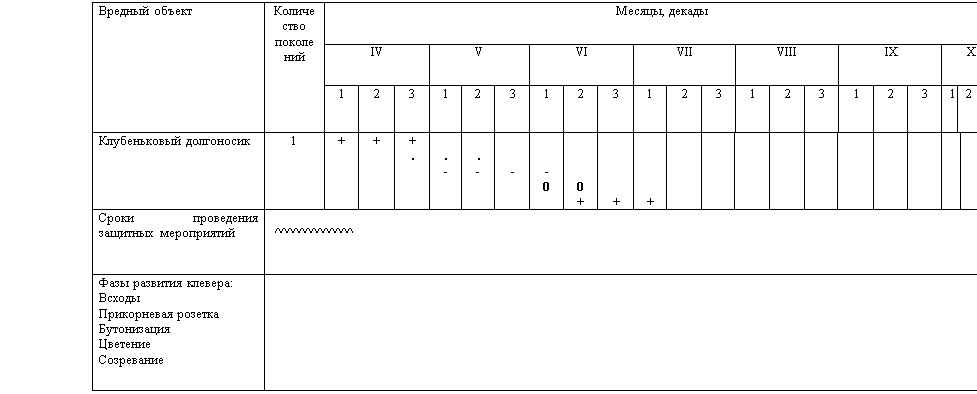

... +. - - 0 - 0 + + + Таблица 3. Фенологические сроки развития клубенькового долгоносика 2.2 Биология возбудителей болезней лугового клевера Антракноз Возбудитель антракноза – несовершенный гриб Colletotrichum trifolii Bain et Essary. ...
0 комментариев