Навигация
1. Конъюгированные антигены
Исследование этого класса антигенов сыграло важную роль для понимания организации детерминантных групп природных антигенов, реальной оценки информационной «емкости» иммунной системы в филогенезе и онтогенезе.
Ниже приведен пример синтеза конъюгированного антигена на основе реакции азосочетания диазобензолсульфоновой кислоты с остатками тирозина в молекуле белка, например, бычьего сывороточного альбумина:
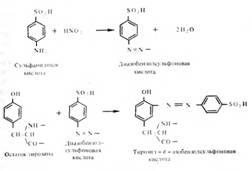
При иммунизации кроликов таким конъюгатом образуется несколько видов антител, реагирующих как с белком, так и с остатком сульфаниловой кислоты. Наличие антител к сульфанилату можно установить, синтезировав тест-антиген и аналог детерминантной группы — гаптен.
Тест-антиген получают, конъюгировав с диазобензол-сульфоновой кислотой другой тирозинсодержащий белок; последний не должен иметь сходства антигенной структуры с белком — носителем детерминантной группы, использованным для иммунизации. В избранном нами случае таким белком может быть яичный альбумин. Если при добавлении тест-антигена к исследуемой антисыворотке образуется преципитат, существование антител против сульфаниловой кислоты можно считать доказанным.
Для точной оценки структуры детерминантной группы синтезируют гаптены. В рассматриваемом примере гаптеном послужит конъюгат тирозина с сульфаниловой кислотой. Такой гаптен не преципитирует антител, но, соединяясь с ними, блокирует активные центры. В результате антитела утратят способность взаимодействовать с тест-антигеном. Реакция ингибирования — эффективный метод тестирования антител против простых по химическому строению гаптенов.
Реакцию низкомолекулярного гаптена с антителами можно оценить с помощью прямых реакций, среди которых наибольшее применение получил метод равновесного диализа. Полученные в эксперименте данные позволяют рассчитать константу равновесия в системе гаптен-антитело. Если определять величины константы равнозесия при реакции антител к определенному конъюгированному антигену с рядом сходных по строению гаптенов, можно оценить вклад каждого радикала в структуру детерминантной группы. При этом удобно сопоставлять сродство к антителу какого-то аналога детерминантной группы со сродством наиболее близкого к ней по строению гаптена. Таким способом получают относительную величину константы связывания: ДотН= = Kx/Kv.T, где Кх — константа связывания исследуемого аналога, Ср.г — константа связывания референс-гаптена.
Ниже приведены величины /Сотн для ряда нитрофе-нильных производных, реагирующих с антителами кролика против динитрофенилированного гемоцианина, т. е. степень сродства антидинитрофенильных антител кролика к различным гаптенам. Антиген получали, модифицируя белок по е-аминогруппам лизина динитрофенилсульфоно-вой кислотой. Поскольку е-динитрофениллизин наиболее близок по строению детерминантной группе использованного конъюгированного антигена, его использовали в качестве референс-гаптена.
Из анализа приведенных ниже данных следует, что антитела распознают как ДНФ-группу, так и аминокислотный остаток, к которому эта группа присоединена. При замене лизина на любую другую аминокислоту сродство антител к гаптеиу резко снижается. Даже в случае использования в качестве гаптена е-динитрофениллизина, в котором взят неприродный оптический изомер, отчетливо заметно снижение константы связывания. Особенно низкая константа связывания найдена в случае отсутствия в молекуле гаптена аминокислотного остатка (динитрофенол). Практически не связываются с изучаемыми антителами мононитрофенильные производные аминокислот. Данные были получены и для других антител к конъюгированным антигенам.

Необходимость стерического соответствия антигенной детерминанты и активного центра — антидетерминанты — антитела со всей очевидностью вытекает, в частности, из того факта, что антитела, направленные к орто-, мета- и пара-аминобензойным кислотам, практически не реагируют перекрестно. Антитела также отчетливо различают право- и левовращающие изомеры виннокаменной кислоты. Менее очевидным является факт строгой специфичности распознавания антителами р-аминобензойной и р-аминофенилсульфоновой кислот:
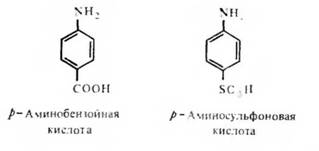
Оба ароматических соединения имеют в параположении отрицательно заряженный заместитель. Заместители отличаются степенью своей нуклеофильности, которая у сульфогруппы ощутимо выше, чем у карбоксильной группы, что приводит к более выраженному смещению облака я-электронов ароматического кольца в направлении заместителя, представленного сульфогруппой в сравнении с карбоксильной группой. Это обстоятельство и различия в размере замещающей группы достаточны, очевидно, для того, чтобы каждое соединение реагировало только с направленным к нему антителом.
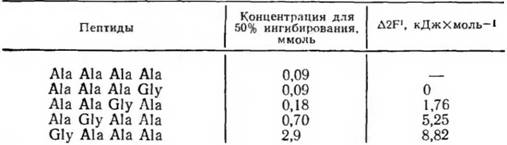
Вопрос о размере детерминантной группы конъюги-рованного антигена, вкладе в ее специфичность различных радикалов был изящно проанализирован П. Шехтером. В качестве антигена использовали конъюгаты белка с олиго-О-аланином. Реакцию между антителами к поли-О-аланину и тест-антигеном ингибировали с помощью различных по размеру олигопептидов из D- и L-аланнна. Как оказалось, ингибирующий эффект гаптенов нарастал от ди- к тетра-О-аланнну. Дальнейшее увеличение длины пептида не усиливало его ннгибирующих свойств. Тетра-Ь-аланин был совершенно не активен как ингибитор. Эти данные означают, во-первых, что антитела четко различают олигопептиды из право- и левовращающих аминокислот, не имеющие регулярной вторичной структуры. Во-вторых, очевидно, что активный центр антитела соответствует по размеру тетрапептиду.
Дальнейшие опыты показали, что вклад каждого остатка аланина в связывание тетрапептида антителом далеко не одинаков. Были синтезированы аналоги гаптена, в которых один из остатков аланина заменяли на глицин. В случае замены N-концевого аланина на глицин константа связывания такого гаптена по сравнению с тетрааланином уменьшалась в 100 раз. Напротив, замена С-концевого аналина на глицин почти не сказывалась на величине константы связывания. Поскольку пептиды присоединяли к белку-носителю через С-концевую группу, можно заключить, что решающий вклад в связывание гаптена антителом вносит наиболее удаленный от молекулы белка-носителя участок присоединенного к ней пептида: в рассматриваемом случае метильная группа N-концевого аланина. Такая группа в молекуле гаптена получила название иммунодоминантной.
Таблица 1. Исследование специфичности антител к поли-Ь-алани-ну с помощью тетрапептидов различного строения
Конъюгированные антигены оказались весьма полезными для изучения многих ключевых проблем клеточной иммунологии, вопросов регуляции иммунного ответа.
Похожие работы
... активные центры) по уровню специфичности уступают только активным центрам антител. Гены МНС класса 3 кодируют отдельные компоненты системы комплемента. Процессинг антигенов- это их судьба в организме. Одной из важнейших функций макрофагов является переработка антигена в иммуногенную форму (это собственно и есть процессинг антигена) и представление его иммунокомпетентным клеткам. В процессинге, ...
... Рассчитывая по Т-критерию достоверность различия выборок получаем что различия ДОСТОВЕРНЫ с вероятностью 95%. Рассчитываем достоверность различий двух выборок уровня CD4/8 антигена в первой группе пациентов. 1,14 1,03 0,209994 испр дисп СД-2 0,458251 испр СКО СД-2 1,20 1,50 γ= 0,999 1,52 1,70 n= 34 ...
... їнська науково-практична конференція, 15-17 квітня 2002 р.: тези доповіді – К., 2002. – С.25. АНОТАЦІЯ Медвідь Я.І. Вплив антигіпертензивних препаратів різних груп на структурно-функціональний стан серцево-судинної системи у хворих з артеріальною гіпертензією та метаболічними порушеннями. – Рукопис. Дисертація на здобуття наукового ступеня кандидата медичних наук за спеціальністю 14.01.02 – ...
... в крови продолжительное время. НВеAg: Этот антиген появляется в крови одновременно с HBsAg. Его обнаружение свидетельствует о высокой активности вируса. Чем больше этого антигена, тем "заразнее" человек. Антитела анти-НВе: Антитела к НВе появляются после исчезновения из крови соответствующего антигена. Снижение содержания в крови HBeAg и появление анти-НВе свидетельствует о том, что ...
0 комментариев