Навигация
Состав и метаболизм аминокислот и белков
27690
знаков
3
таблицы
0
изображений
2. Состав и метаболизм аминокислот и белков
Впервые сравнительное исследование аминокислотного состава нейронов и нейроглии было проведено Г. Роузом. Полученные экспериментальные данные позволили сделать заключение о том, что содержание свободных аминокислот в нейронах выше по сравнению с нейроглией. Исключение отмечено лишь для глутаминовой кислоты, содержание которой несколько выше в клетках нейроглии.
Одновременно с изучением распределения свободных аминокислот был исследован их метаболизм. Оказалось, что при использовании в качестве предшественника биосинтеза аминокислот иС-глюкозы или иС-пирувата нейроглиальные аминокислоты включают радиоактивный углерод в среднем в три раза интенсивнее. Несмотря на то, что эти исследования, выполненные в опытах in vitro, естественно, не могут в полной мере охарактеризовать свойства нейронов и нейроглии, все-таки можно предположить, что одной из характерных особенностей нейроглиальных клеток является более высокий метаболизм свободных аминокислот.
Особое положение занимает вопрос о взаимопревращениях глутамата и глутамина в клетках нейроглии и нейрона. В экспериментах с интрацеребральным введением С-глутамата через 15–30 мин удельная радиоактивность глутамина в нейронах была ниже, чем глутамата. Напротив, в нейроглии уровень радиоактивности глутамина превышал средний уровень радиоактивности глутамата. Это были первые указания на существование нескольких метаболических компартментов для глутамата и на своеобразное «разделение труда» между нейронами и глией в отношении синтеза, распада и перемещений двух нейромедиа-торов – глутамата и гамма-аминомасляной кислоты. Особенностью внешней мембраны нейрона явилась низкая проницаемость для глутамата и высокая для глутамина. Пока трудно дать хорошо обоснованное объяснение этому факту. Можно лишь полагать, что это связано с двойственной ролью глутамата в ЦНС: обычной – в качестве компонента синтезируемых белков, и специальной – как нейромедиатора и как предшественника другого нейромедиатора – ГАМК. В результате глута-мат из внеклеточной среды поглощается глией, превращающей его в форму, способную войти в нейроны, – глутамин. Последний, выйдя из глии и войдя в глутаматергические нейроны, дезаминируется, образуя вновь глутамат. Далее он включается в синаптические Лузырьки – хранители медиатора. При прохождении импульса они опорожняются в синаптическую щель, глутамат опять поступает в глию и таким образом цикл замыкается- В нейронах другого типа – ГАМКергических – поступивший глутамин не только вновь превращается в глутамат, но и декарбоксилируется, превращаясь в ГАМК. Последняя, опять-таки при прохождении импульса, выходит в синап-тическую щель, и часть ее поступает в глию, где участвует в процессах ресинтеза глутамина. Понятны и важны в этом плане данные о способности нейроглиальных клеток очень активно поглощать аминокислоты из инкубационной среди против градиента концентрации на фоне очень слабой аккумуляции аминокислот нейронами. Наиболее активно глиальные клетки поглощают ГАМК и глутамат. Отношение содержания ГАМК в клетках нейроглии к ее содержанию в инкубационной среде достигает 100, в то время как для нейронов эта величина колеблется в пределах 10. Процесс поглощения ГАМК глиальными клетками зависит от таких факторов, как температура инкубационной среды и наличие ионов К+, Na+ и Mg^+. Ионы К+ стимулируют высвобождение ГАМК из нейронов. Пикротоксин и стрихнин не влияют на захват ГАМК нейронами, тогда как в глии наблюдается усиление поступления ГАМК при действии этих фармакологических веществ; аминазин, напротив, неконкурентно ингибирует процесс поглощенная ГАМК глией.
Глиальные клетки наряду с активным потреблением ГАМК из окружающей среды могут активно ее синтезировать. При инкубации обогащенных клеточных фракций с С-глутаматом радиоактивность ГАМК в нейроглии была в среднем в 40 раз выше, чем в нейронах, при практически одинаковой активности глутаматдекарбоксилазы в нейронах и нейроглии.
Чтобы еше яснее оценить особую роль глии в отношении обмена глутамата и ГАМК, укажем, что по способности аккумулировать другие нейромедиаторы, такие как норадреналин, серотонин, дофамин, нейрональные и нейроглиальные клетки различаются незначительно. Кроме того, активность ацетилхо-линэстеразы, фермента, участвующего в инактивации ацетилхолина в нейронах и нейроглии, практически одинакова.
Иным является отношение глии и нейронов к другой аминокислоте – триптофану – предшественнику серотонина. Нейроны имеют систему, которая характеризуется высоким сродством к триптофану. Психотропные вещества, в частности аминазин и имипрамин, оказывают тормозящее влияние на поглощение триптофана.
Аккумуляция ряда медиаторов глией осуществляется при посредстве расположенных на поверхности клеток так называемых белков-транспортеров. Они имеют много общего по структуре с метаботропными рецепторами.
Накоплено немало данных о наличии, на астропитах не только белков-транспортеров, но и типичных рецепторов глутамата, ГАМК и норадреналина. Роль их неясна, хотя следует иметь их ввиду, учитывая гипотезу о движении сигналов через сеть астроцитов, рассмотренную выше в связи с осцилляцией концентраций ионов Са*.
Исследование особенностей количественного состава и метаболизма свободных аминокислот тесно связано с изучением белкового состава нейронов и нейроглии, которые в значительной степени определяют морфологическую и функциональную специфику этих клеточных популяций в ЦНС.
Анализ общего содержания белка в обогащенных нейронами и нейроглией фракциях свидетельствуют о том, что в глиальных клетках содержание белка несколько выше по сравнению с нейронами. Очевидны принципиальные различия, обусловленные отсутствием в глии аксональных транспортных систем, терминалей, органелл, накапливающих и выбрасывающих в синаптическую щель медиаторы, сложных систем межнейронального узнавания и адгезии и т.п.
Большое значение для понимания роли белков в системе ней-рон-нейроглия имеют исследования их метаболизма. Эти исследования позволяют изучить не только динамическое состояние нейрональных и нейроглиальных белков, но и их взаимоотношения. Экспериментальные данные свидетельствуют о том, что синтез нейрональных белков протекает в 2–3 раза интенсивнее по сравнению с нейроглиальными белками. Метаболизм белков различен не только в зависимости от клеточной популяции, но и внутри самой популяции. Так, установлено, что метаболизм белков крупных нейронов имеет более высокий уровень по сравнению с мелкими нейронами, а белки астроцитов метаболиру-ют интенсивнее белков олигодендроглии. Эта закономерность прослеживается как у взрослых, так и у растущих животных.
Исследование метаболизма белков нейронов и нейроглии проводится in vitro и in vivo, причем следует подчеркнуть, что в опытах in vivo также прослеживается отличие в их биосинтезе на уровне нейронов и нейроглии. Включение различных аминокислот в белки имеет некоторую избирательность. При инкубации срезов коры головного мозга кроликов с мечеными аминокислотами с последующим выделением обогащенных фракций оказалось, что лейцин включается в нейрональные белки в 5–6 раз, а глицин, глутамат и фенил ал анин – в 2,5 раза интенсивнее, чем в белки нейроглии. В отличие от аминокислот: включение иС-глюкозы в белки нейронов и нейроглии практически одинаково, а в некоторых опытах даже выше в глиальной фракции. Несомненный интерес представляют длительные по времени наблюдения белкового метаболизма в субклеточных фракциях нейронов и нейроглии. Установлено, что в зависимости от времени радиоактивной экспозиции наблюдается перераспределение радиоактивной метки между субклеточными фракциями нейронов и нейроглии. Так, через 10 мин после введения С-фенилаланина наибольшая радиоактивность обнаруживается в микросомах, через 20 мин – в митохондриях, а через 45 мин – в ядерной фракции. При исследовании водорастворимой фракции максимальная радиоактивность регистрируется через 15 мин, а затем она снижается и остается на постоянном уровне, что связано с миграцией цитоплазматиче-ских белков в аксон.
Уровень метаболизма белков нейронов и нейроглии при исследовании в опытах in vitro в значительной степени зависит от условий инкубации клеточных фракций. Так, например, включение Н-лейцина в нейрональные белки значительно увеличивается по мере нарастания парциального давления 02, тогда как в клетках нейроглии практически не наблюдается каких-либо изменений.
Особый интерес для понимания механизмов, лежащих в основе работы системы нейрон-нейроглия, представляют исследования, в которых проводится изучение процессов метаболизма белков при изменении функционального состояния ЦНС. Имеющиеся экспериментальные данные свидетельствуют о том, что изменение функционального состояния влечет за собой неодинаковые изменения в процессах метаболизма белков в нейронах и нейроглии. Так, например, при 3-часовой гипоксии включение Н-лейцина увеличивалось в белки нейронов и уменьшалось в белки нейроглии. В этот период различия в уровне метаболизма в нейронах и нейроглии были не очень значительными. Однако на фоне 16-часовой гипоксии наблюдалось резкое увеличение включения изотопа в белки нейронов. При локальном у-облучении коры головного мозга кролика через 2 дня происходит уменьшение включения Н-лейцина в белки как нейронов, так и нейроглии. В течение последующих 2 недель наблюдалось усиление включения метки в белки нейронов на фоне уменьшения включения в белки нейроглии. Такие воздействия, как алкогольная интоксикация и аноксия, вызывают снижение синтеза белков в нейроглии, в то время как в нейронах практически не происходит изменений. В то же время прямые ингибиторы белкового синтеза пуромицин и циклогексимид значительно снижают скорость включения аминокислот в ней-рональные белки.
Рассмотренный экспериментальный материал по метаболизму белков в экстремальных условиях показывает, что в нейроглии происходит значительное снижение скорости метаболизма белка, в нейронах эти воздействия не вызывают снижения метаболической активности белков, а даже наоборот, при облучении и гипоксии наблюдается усиление их обмена, что, по-видимому, обеспечивает «нормальную» работу нейронов при увеличении функциональной нагрузки. В то же время действие прямых ингибиторов белкового синтеза вызывает более значительное угнетение белкового метаболизма в нейронах, что связано с большей чувствительностью белоксинтезируюших систем нейронов по сравнению с нейроглией. Таким образом, на примере метаболизма белков и аминокислот подтверждается вывод о существовании единой, но строго комиартментализованной метаболической системы нейрон-нейроглия, в которой процессы синтеза и распада белков и аминокислот теснейшим образом связаны и взаимообусловлены.
Похожие работы
... междисциплинарными связями. Студенты, приступающие к изучению нейрохимии, должны быть уже вооружены основательными знаниями по общей биохимии. Это обуславливает целесообразность преподавания нейрохимии на старшем курсе специальности «биохимия». Дисциплина относится к региональному (вузовскому компоненту). Распределение времени, отведенного на изучение дисциплины по учебному плану Форма ...
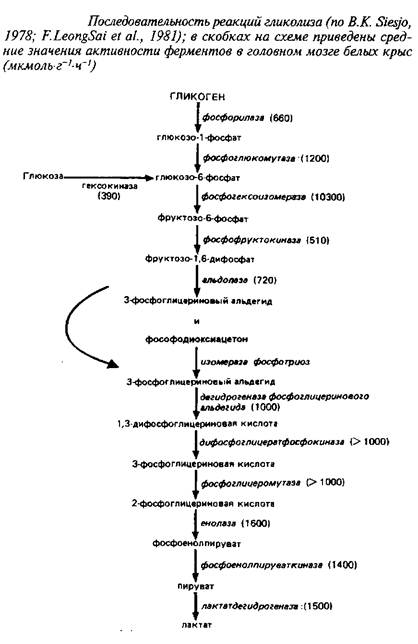
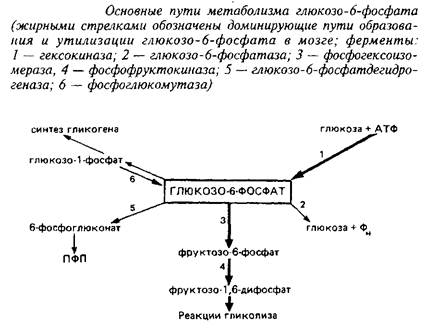
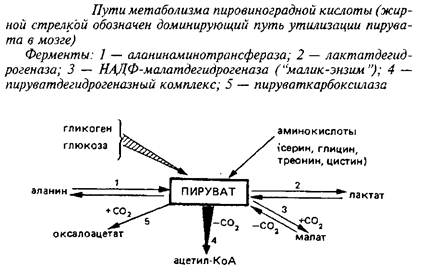
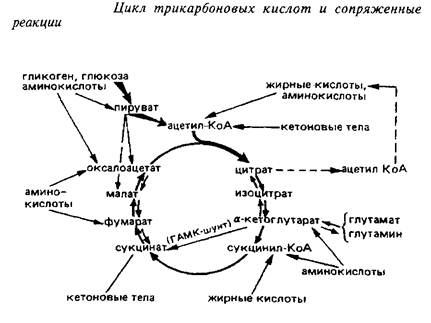
... 4,8±0,6 Аспарагиновая кислота + аспарагин 4,8–5,8 0,4–0,7 6,0±1,5 6,3±0,8 ГАМ К 1,9–2,4 Следы 3,1 + 1,1 4,1±0,4 Из других путей метаболизма аминокислот, которые играют относительно небольшую роль в энергетическом метаболизме головного мозга, можно упомянуть превращение аспартата и аспарагина в оксалоацетат, а также аланина и серина – в пируват. Очень невелико и значение аминокислот как ...
... нерастворимы в воде, но могут быть экстрагированы растворами солей. К другим сократительным белкам относятся тропомиозин и комплекс тропонина (субъединицы Т, 1, С), содержашиеся в тонких нитях. В мышце содержатся также миоглобин, гликолитические ферменты и другие растворимые белки, не выполняющие сократительной функции Белковый состав скелетной мышцы Белок Молекулярная масса, дальтон, тыс. ...
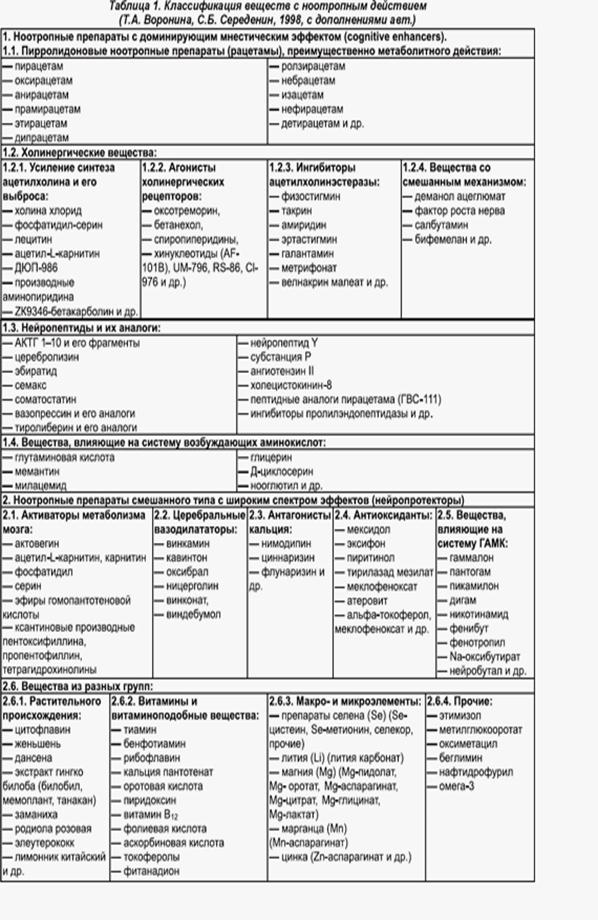
... кальциевых каналов. Нимодипин Нимодипин - блокатор кальциевых каналов, производное дигидропиридина. Избирательно взаимодействует с кальциевыми каналами типа L и блокирует трансмембранное поступление ионов кальция. Фармакологическое действие: Особенностью препарата является преимущественное влияние на кровоснабжение мозга. Препарат оказывает дилатирующее действие на сосуды головного мозга ...
0 комментариев