Реферат
На тему: «Генетическая регуляция развития»
Мутации и изменения, происходящие в онтогенезе
Если морфология представляет собой проявление сложного комплекса процессов развития, то эти процессы в свою очередь представляют собой проявление действия целого созвездия генов. Таково важное допущение, положенное в основу этой книги, и мы собрали по крайней мере prima facie данные в пользу того, что у Metazoa часть генома специфически участвует в регуляции онтогенеза и что характер эволюции этой части отличается от характера эволюции структурных генов. До сих пор, однако, мы ограничивались изолированными частными примерами генного контроля морфогенеза и не пытались ответить на центральный вопрос: каким образом гены определяют процесс развития? Здесь мы в весьма прямом смысле возвращаемся к программе механики развития, выдвинутой Ру (Roux), однако, вместо того чтобы удалять клетки или другие структуры зародыша, с тем чтобы определить их роли в развитии, как это делали представители классической экспериментальной эмбриологии, генетика развития использует мутации как очень тонкий скальпель, дающий возможность уничтожать или изменять отдельные гены.
Генетическая модель, с помощью которой можно изучать любую систему, теоретически находящуюся под генетическим контролем, состоит в следующем. Для того чтобы проанализировать какой-либо процесс, в данном случае онтогенез, исследователь выявляет мутации, изменяющие этот процесс. Выявив такие мутации, он проводит фенотипическое сравнение мутантных особей с нормальными. Это сравнение помогает ему понять, как данный ген влияет на нормальное развитие. Однако, прежде чем продолжить описание метода проведения такого сравнения, следует указать, что воздействие мутаций на онтогенез проявляется двумя основными способами. Это, во-первых, дизруптивные изменения, при которых процесс нормального развития нарушается, что приводит к морфологическим аномалиям (например, к отсутствию некоторых структур). В наиболее резко выраженной форме такие мутации оказываются летальными. Во-вторых, это гомеозисные изменения, при которых под действием мутации развитие отклоняется от нормы, в результате чего какая-либо структура данного организма замещается гомологичным органом или конечностью. Мы отложим дальнейшее обсуждение мутаций этого второго типа до следующей главы и займемся здесь главным образом дизруптивными изменениями.
Анализ нарушений, вызываемых какой-либо дизруптивной мутацией, лишь в редких случаях сводится к простому сравнению конечного фенотипа гибнущей особи с нормальным фенотипом, потому что развитие – это сложный и высокоинтегрированный процесс. Огромное большинство происходящих в нем событий тесно связано с другими событиями и, в сущности, зависит от них. Это особенно ясно проявляется в том, что многие мутации обладают плейотропным действием, т.е. отсутствие или изменение одного гена может обусловить несколько морфологических изменений. Примером служат изменения, наблюдаемые у людей с так называемой аномалией Пельгера (Pg). Она наследуется у человека как простой доминантный аутосомный признак. У гетерозигот (Pg/+) нет никаких клинических симптомов, но для их нейтрофилов характерны аномально сегментированные ядра. У взрослого человека ядра полиморфноядерных нейтрофилов обычно состоят из четырех или пяти сегментов, у гетерозигот же Pg/+ ядра состоят всего из двух, реже из трех сегментов. Этот же признак обнаружен у кроликов, у которых он наследуется по тому же типу и сходным образом проявляется в картине крови. Скрещивая гетерозиготных кроликов, можно получить гомозиготных особей Pg/Pg. Ядра нейтрофилов у этих особей вообще не разделены на сегменты, и такой генотип сопровождается низкой жизнеспособностью. Для немногих выживших особей помимо этой особенности нейтрофилов характерна крайне выраженная карликовость с недоразвитием конечностей и грудной клетки. Здесь следует задать вопрос: какова причинная зависимость, если она существует, между этими двумя плейотропными нарушениями? Не исключена возможность, что оба этих фенотипических проявления представляют собой на самом деле результат третьего, пока еще неизвестного нарушения, вызванного аллелем Pg.
Анализ времени и места действия генов
Методы, используемые для определения первичного места действия гена, сходны с применяемыми в классической эмбриологии и, в сущности, заимствованы из нее. В своей простейшей форме эти методы состоят в пересадке органа или кусочка ткани от мутантной особи нормальному реципиенту. Производят также и реципрокные пересадки. Эта операция обычно проделывается до проявления того или иного мутантного эффекта. Затем можно определить судьбу развивающегося органа или ткани в их новом окружении. Если генетический дефект рассматриваемого органа или ткани автономен для этой структуры, т.е. если она является первичным местом действия данного гена, то следует ожидать, что мутантная ткань будет продуцировать аномальный фенотип (даже и у нормального хозяина). Эксперименты сходного типа можно проводить на тканях или органах, выращиваемых в культуре, примерно таким же образом, экспериментах по совместному выращиванию дермы и эпидермиса мыши, ящерицы и курицы, только вместо соединения тканей от животных разных видов соединяют ткани или органы мутантных и немутантных особей. Создание мозаичных особей проводилось в еще более широких масштабах Минц (Mintz) и ее сотрудниками. Этим исследователям удалось сращивать in vitro целых мышиных зародышей на стадии морулы. Таких «гибридных» зародышей имплантируют затем псевдобеременной самке. Получающиеся в результате мыши, происходящие от четырех родительских особей, состоят из смеси клеток двух разных генотипов, причем активны оба генотипа. Этот метод можно также использовать для анализа автономности экспрессии мутантных генов, сращивая мутантных и нормальных зародышей на стадии морулы.
Следует упомянуть еще об одном методе такого типа – о парабиозе. Он состоит в сращивании целых животных, а не просто органов или тканей. Необходимо указать, однако, что при таком сращивании не получается действительно интегрированных мозаичных особей. При всех этих методах необходимо, чтобы трансплантируемые ткани, органы или сращиваемые зародыши были совместимы. У низших позвоночных, таких как амфибии, это не представляет серьезной проблемы; однако у млекопитающих дополнительное осложнение возникает в связи с возможностью отторжения трансплантата, так что следует сначала убедиться в том, что мутантная и нормальная особи иммунологически совместимы. Остается привести еще два метода, специфичные для генетики развития и применяемые почти исключительно при работе с Drosophila melanogaster. Это создание гинандроморфов и индукция мозаичных особей путем митотической рекомбинации. Гинандроморфами называют взрослых мух, тело которых состоит как из мужских, так и из женских тканей. Используя тип дробления, характерный для двукрылых, и их особую кольцевую Х-хромосому, таких мозаичных особей можно постоянно получать в лабораторных линиях. В норме X-хромосома дрозофилы имеет форму палочки, на одном конце которой находится центромера. Одна из мутантных форм этой хромосомы представляет собой замкнутое кольцо. Кольцо это обладает интересной особенностью: при нескольких первых делениях дробления оно нестабильно. Эта нестабильность может привести, в частности, к утрате кольцевой хромосомы одним из двух дочерних ядер, образующихся при первом делении дробления. Если утрата происходит на этой стадии, то в результате дальнейших делений дробления создается популяция ядер, одна половина которых содержит кольцевую Х-хромосому, а другая не содержит ее. Если зигота, начиная делиться, представляет собой гетерозиготную самку – палочка-Х/кольцо-Х, то после такой утраты половина ее ядер окажется женскими и будет содержать две Х-хромосомы – кольцо-Х/палочка-Х, а другая половина – мужскими и будет содержать только одну Х-хромосому – палочка-Х/О (пол у дрозофилы определяется соотношением Х-хромосом и аутосом, а не Y-хромосомой, как у млекопитающих). После восьми синцитиальных делений в яйце образуется скопление ядер. Это скопление, однако, не представляет собой случайной смеси типов ХО и XX. Ядра этих двух типов образуют две пространственно разделенные группы, расположение которых определяется плоскостью первого деления дробления. Поэтому, когда такая популяция ядер мигрирует к периферической цитоплазме, с тем чтобы образовать клеточную бластодерму, она мигрирует в виде двух соприкасающихся, но обособленных групп мужских и женских ядер. Взрослая муха, развивающаяся из такого гинандро-морфного зародыша, также будет мозаичной. Как показано на рис. 7–3, количество и расположение взрослой ткани мужской или женской природы непостоянно. Это происходит потому, что плоскость первого деления дробления располагается случайным образом по отношению к осям яйца. Поэтому если плоскость первого дробления делит яйцо по длинной оси на правую и левую половины, то из него разовьется билатеральный гинандроморф. Вариации этого простого случая приведут к большей или меньшей доле мужской ткани в зависимости от того, сколько ядер ХО находится в тех участках бластодермы, которым суждено сформировать взрослые ткани.
Мутации с материнским эффектом
У таких разных организмов, как морские ежи и лягушки, события, происходящие на ранних стадиях дробления, и, в сущности, большая часть, если не все развитие, предшествующее гаструляции, не зависят от генома зиготы. Информацию, необходимую для выполнения этих начальных и решающих этапов онтогенеза, определяет материнский геном при образовании яйцеклетки. На примере закручивания раковины у Limnaea, такое заключение подтверждается существованием у широкого круга различных организмов так называемых генов с материнским эффектом. Мутации этих генов передаются по наследству чрезвычайно своеобразным способом. При скрещивании двух особей, гетерозиготных по какому-либо рецессивному признаку, следует ожидать, что этот признак проявится у 25% потомков. Однако в случае материнских (mat) мутаций особи mat/mat развиваются нормально. Более того, мужские особи с таким генотипом фертильны и при скрещивании с нормальными женскими особями дают нормальных потомков. В отличие от этого гомозиготные самки дают аномальных потомков. Это объясняется тем, что у таких самок образуются аномальные яйцеклетки, которые не могут завершить нормальное развитие. Самка mat/mat выживает, потому что она происходит от гетерозиготной (mat/+) матери, способной продуцировать нормальные яйца. Хотелось бы сделать вывод, что гены, дающие такие мутации, продуцируют какие-то «морфогены», которые образуются в развивающемся ооците в качестве «инструкции» для раннего развития. Однако возможно также, что яйцо неспособно развиваться просто вследствие какого-то общего нарушения метаболизма. Подходящим примером служит группа из пяти различных дефектов, наследуемых по материнскому типу и определяемых генами, локализованными в Х-хромосоме Drosophila melanogaster: tin (cinnamon), dor (deep orange), amx (almondex), fu (fused) и r (rudimentary). Все эти признаки, помимо того что они наследуются по материнскому типу, вызывают у взрослых особей заметные морфологические отклонения, по которым они и получили свои красочные названия. Гемизиготные самцы, обладающие любой одной из этих мутаций, жизнеспособны и фертильны, так же как и гетерозиготные самки. Скрещивая мутантных самцов с гетерозиготными самками, можно получить гомозиготных самок, которые при скрещивании с мутантными самцами оказываются совершенно стерильными. Например, самки dor/dor продуцируют яйца, развитие которых прекращается на стадии гаструляции. Остальные четыре мутации также вызывают гибель зародышей, но на несколько другой стадии, чем мутации dor. В характере наследования всех этих пяти мутаций есть еще одна аномальная особенность. Скрещивая гомозиготных мутантных самок с нормальными самцами, можно получить некоторое число потомков. Все это – гетерозиготные самки, развившиеся из яиц, оплодотворенных сперматозоидом, несущим Х-хромосому. Ни один самец не выживает. По-видимому, присутствие аллеля дикого типа рассматриваемого гена может несколько снизить дефектность яйца, даже если этот аллель вносится сперматозоидом. Это, конечно, подразумевает, что по крайней мере часть генома зиготы активна во время гаструляции.
Мутации, затрагивающие органогенез
Как мы убедились выше, события, происходящие на ранних стадиях развития, в значительной степени зависят от информации, поставляемой материнским организмом. Однако примерно ко времени гаструляции важную роль в дальнейшем развитии начинает играть генетическая информация самого зародыша, и организм приобретает возможность контролировать свою судьбу. Для морфогенетических событий, следующих за формированием бластодермы, необходим синтез РНК и ее трансляция в белок. О необходимости генетической информации зародыша можно также судить по большому числу мутаций, оказывающих влияние на события, происходящие после гаструляции, и указывающих, таким образом, на существование генов, регулирующих эти события. У этих мутаций не наблюдается наследования по материнскому типу.
В качестве примера можно воспользоваться локусом N (Notch) у Drosophila melanogaster. Notch-сцепленная с полом доминантная мутация, являющаяся одновременно рецессивной деталью. Гомозиготные самки (N/N) и гемизиготные самцы (N/Y) гибнут на стадии зародыша спустя примерно 6 ч после оплодотворения. Это время соответствует моменту, непосредственно следующему за гаструляцией, когда зародыш проделал примерно четвертую часть эмбриогенеза. Гистологические и морфологические исследования, проведенные Паулсоном (Paulson) на этих зародышах, показали, что вентральная и латеральная эктодерма, которая обычно дает начало эпидермису и нервным клеткам, образует только клетки, похожие на нейробласты, а эпидермиса не образует вовсе. Поэтому можно предполагать, что локус Notch необходим для дифференцировки из зародышевой эктодермы нервной ткани в противовес эпидермальной. Оказалось, однако, что это несколько упрощенное объяснение. Шелленберджер (Shellenbarger) и его сотрудники выделили из локуса Notch температурочувствительный аллель и охарактеризовали его. Мухи, содержащие этот аллель, при температуре 22 °С развиваются нормально, тогда как при 29 °С наблюдается описанная выше гибель зародышей. В экспериментах со сдвигами температур, подобных рассмотренным в начале этой главы для мутантов shibire, было установлено, что к дефекту локуса Notch и недостаточности продукта этого локуса зародыш чувствителен не только в этот ранний период эмбриогенеза. При помощи кратковременных повышений температуры были выявлены еще три периода, когда мутантному организму жизненно необходима нормальная активность локуса Notch. Воздействия непермиссивной температуры на личинок второго или третьего возраста или на куколок приводили к летальному исходу. Более того, кратковременные воздействия (pulses) в определенные периоды третьей личиночной стадии или на стадии куколки вызывали такие же рубцы на глазах и дефекты щетинок, как у мутантов shibire. Поэтому, подобно shibire, мутация Notch обладает гораздо более широким действием, чем можно было бы ожидать на основании ее главного фенотипического проявления. Все структуры, на которые она действует, эктодермального происхождения, и в своих дальнейших экспериментах с гинандроморфами Шелленберджеру удалось показать, что наблюдаемые морфологические дефекты автономны и присущи только эктодермальным клеткам. Таким образом, снова, как и в случае мутаций shibire, создается впечатление, что один и тот же продукт мутанта Notch необходим разным клеткам эктодермального происхождения в течение нескольких дискретных периодов на всем протяжении развития. Следует также отметить, что для завершения одного и того же набора онтогенетических событий необходимы два разных гена, Notch+ и shibire+, и что отсутствие того или другого из них приводит к удивительно сходному комплексу нарушений. У домовой мыши (Mus musculus) имеется один сложный ген, Т-локус, который во многом сходен с только что описанной системой Notch. Первый аллель этого локуса был описан как аутосомный доминантный аллель, названный Brachyury (Т). У мышей, гетерозиготных по этому гену, Т/+, хвосты короткие. В гомозиготном состоянии (Т/Т) этот ген летален и зародыши гибнут внутриутробно. Вскоре после обнаружения этой доминантной мутации было установлено, что потомки от скрещивания гетерозигот (Т/+) с мышами дикого типа часто вовсе лишены хвоста. Оказалось, что эти бесхвостые мыши – результат рецессивных аллелей Т-локуса, часто встречающихся в природных популяциях мышей. Следовательно, эти бесхвостые мыши имели генотип T/t. От скрещиваний между такими гетерозиготными мышами T/t были получены бесхвостые мыши, размножающиеся в чистоте. Позднее было показано, что это обусловлено «сбалансированной системой леталей». Как оказалось, среди потомков от скрещивания гетерозигот летальными были не только гомозиготы Т/Т, чего следовало ожидать, но и гомозиготы t/t. Таким образом, выживали только гетерозиготы T/t, которые и давали следующее поколение. Эта интригующая ситуация была изучена как генетически, так и эмбриологически в ряде изящных работ Денна (Dunn), его учеников Беннета (Bennett) и Глюксон-Вэлша (Gluechsohn-Waelsch).
В результате генетических исследований вновь полученных линий, сбалансированных по леталям, было установлено, что скрещивания между бесхвостыми мышами, получившими свой рецессивный аллель t от разных популяций, часто давали нормальных потомков. В частности, от скрещивания T/ta x T/tb были получены нормальные и бесхвостые потомки в соотношении 2:1. Можно показать, что потомки с нормальными хвостами имеют генотип ta/tb. Таким образом, этот генотип был не только нелетальным, но и нормальным морфологически. Еще одна особенность этого локуса была обнаружена при скрещиваниях между самими линиями, сбалансированными по летал ям. Обычно скрещивание гетерозиготы T/ta с такой же гетерозиготой дает только бесхвостых потомков. Однако изредка (1:500 – 1:1000) среди потомков встречаются мыши с нормальными хвостами. Эти нормальные мыши почти всегда получаются в результате редкой генетической рекомбинации в 17-й хромосоме, в участке, соответствующем локусу Т или примыкающем к нему. Можно показать, что наличие хвоста у этих реком-бинантных потомков обусловлено тем, что одновременно с рекомбинацией произошло превращение первоначального аллеля ta в новый аллель tx, который комплементарен аллелю ta таким же образом, как некоторые t-аллели, происходящие от мышей дикого типа, комплементарны друг друга. Различие, конечно, состоит в том, что tx в данном случае непосредственно связан с ta своим происхождением. Было показано, что такое превращение одного аллеля t в другие комплементарные типы происходит с большинством выделенных рецессивов. Некоторые возникающие при этом аллели, например tx, могут в свою очередь при помощи того же механизма превратиться в другой комплементарный аллель, например ty. Такое превращение одного рецессивного аллеля t в другой приводит к образованию ряда постепенно переходящих один в другой аллелей, а в конечном итоге к созданию так называемых t viable, или tv, аллелей. Все эти последние аллели нелетальны, и бесхвостый фенотип экспрессируется только в случае гетерозигот Т/tv; у особей tv/tvхвосты нормальные. Еще один класс рецессивных t-мутаций – это полулетали, жизнеспособность которых колеблется от 2 до 51% нормальной. Как и полностью летальные аллели, они путем постепенных превращений дают tv-типы. Комплементарные скрещивания (T/ta x T/tb) всех как природных, так и полученных экспериментально рецессивных аллелей показали, что 111 существующих мутаций распадаются на 8 отдельных групп, причем ни одна из них не комплементарна Т-аллелю. Число членов во всех группах различно – от одного в группе (tw73 до 66 в группе аллелей tv. Из пяти доминантных Т-мутаций одна была вызвана воздействием рентгеновских лучей, а все другие мутации, обнаруженные в этом локусе, по-видимому, возникли спонтанно.
Мы рассмотрели несколько особенностей этого сложного ряда генетических дефектов, связанных между собой либо тем, что они комплементарны и определяются соседними локусами, либо тем, что они происходят друг от друга. Перейдем теперь к эмбриологическим особенностям экспрессии сложного локуса.
Каждая из восьми комплементарных групп обусловливает разного рода дефекты, проявляющиеся в период от ранних до поздних стадий развития зародыша. Морфологические проявления этих дефектов представлены на рис. 7–10 и 7–11. Самые ранние дефекты обнаружены у зародышей, гомозиготных по аллелю t12. Оплодотворение и деления дробления зиготы приводят к образованию шаровидной массы клеток, называемой морулой. Первый признак клеточной дифференцировки у мышей наблюдается при переходе от этой морулы к следующей стадии – бластоцисте, состоящей из трофобласта (трофэктодермы) и внутренней клеточной массы. Гомозиготные t12-зародыши не достигают этой стадии и недифференцированные «морулы» гибнут, не имплантируясь в стенку матки, в отличие от нормальных зародышей, которые делают это примерно на 4-й день после оплодотворения. Кроме того, t12-клетки, по-видимому, автономны в своем летальном действии. Химеры, содержащие эмбриональные клетки t12 и нормальные эмбриональные клетки, летальны, и их развитие не заходит дальше той стадии, на которой гибнут зародыши t12. Поэтому мутация t12, по-видимому, затрагивает локус, необходимый для первого шага в дифференцировке мышиного зародыша – возникновения трофобласта, из которого в конечном итоге формируются хорион и другие характерные для всех плацентарных млекопитающих внезародышевые оболочки, образующиеся из зиготы.
Мутации, воздействующие на развитие определенных органов
В то время как локус Т оказывает, по-видимому, самые разнообразные воздействия на все развитие эктодермы, существуют мутации, вызывающие более специфические дефекты. Примером служит мутация cardiac lethal (с) у аксолотля Ambystoma mexicanum. Эта мутация была впервые обнаружена и исследована Хэмфри (Humphrey). Хэмфри установил, что мутация с наследуется как простой аутосомный рецессивный признак, так что при скрещивании двух гетерозиготных особей (с/+) 25% потомков гибнет на ранних личиночных стадиях, вскоре после вылупления. Эти мутантные особи плавают, как нормальные личинки, но они раздуты переполняющей их жидкостью, а пищеварительная система и жабры у них недоразвиты. Первичная причина этих дефектов – нарушение развития сердца и его неспособность к сокращениям. Поэтому у мутантных личинок отсутствует кровообращение, а дышат они, вероятно, путем диффузии через кожу, что дает им возможность просуществовать лишь в течение ограниченного периода времени. Как показал Хэмфри путем сращивания нормального и мутантного зародышей, это нарушение развития сердца носит автономный характер.
У мутантного и нормального зародышей, взятых до закладки сердца, удаляли по кусочку ткани с боковых поверхностей тела. Затем зародышей соединяли по месту раны и давали им срастись. В тех случаях, когда такие сросшиеся особи завершали развитие, было установлено, что нормальный партнер ослаблял раздутость и другие дефекты зародыша с/с, давая ему возможность выжить. Однако сердце мутантного партнера навсегда оставалось простой трубкой, неспособной к сокращениям, и кровообращение мутанта целиком обеспечивал нормальный партнер. Установлено, что у позвоночных многие органы, в том числе сердце, развиваются в результате определенных индукционных взаимодействий, происходящих во время развития. В частности, Джекобсон и Дункан (Jacobson, Duncan) показали, что у хвостатых амфибий развитие сердца из мезодермальных зачатков индуцируется головным участком энтодермы. Причиной того, что особям с/с не удается образовать сердце, может быть неспособность головного участка энтодермы индуцировать этот процесс или же неспособность сердечной мезодермы реагировать на индуктор. Для того чтобы выяснить, какая из этих двух причин вызывает нарушение развития, Хэмфри пересаживал нормальную сердечную мезодерму с/с-реципиентам, а мезодерму с/с-мутантов – нормальным реципиентам. Оказалось, что мезодерма с/с способна образовать сокращающееся сердце под индукционным воздействием нормального головного участка энтодермы, тогда как мутантные зародыши не могут обеспечить развитие нормального сердца. Эти результаты можно интерпретировать как указание на отсутствие индукционной активности головного участка энтодермы у мутанта с/с. Однако возможно также, что мутантные особи активно подавляют формирование сердца. Эксперименты, проведенные Лемански (Lemanski) и его сотрудниками, делают последнее предположение менее вероятным. Эти авторы выращивали in vitro мезодермальные зачатки сердца мутантных и нормальных зародышей. В использованных ими условиях в нормальной сердечной мезодерме происходили сильные сокращения, а в мутантной их не было. Если считать, что сокращения мутантной ткани подавлялись in situ, то выращивание in vitro должно было снять это воздействие. Более того, при совместном культивировании мезодермы мутанта с/с и головного участка энтодермы нормального зародыша в мутантной ткани начинались сокращения; это показывает, что мутантная мезодерма способна нормально реагировать на соответствующее индукционное воздействие. Таким образом, ген «cardiac lethal», очевидно, обусловливает неспособность головного участка энтодермы обеспечить индукционный сигнал, запускающий дифференцировку сердца из его мезодермального зачатка.
Гибель клеток в процессе нормального развития
Дегенерация вольфовых протоков в процессе развития особей женского пола и мюллеровых каналов у особей мужского пола – примеры наличия структур, которые вначале развиваются, а затем подвергаются некрозу. Эти процессы, подобно всем другим рассматриваемым здесь онтогенетическим событиям, находятся под контролем генов. Специфическим примером служит развитие конечностей у четвероногих. У цыпленка почки конечностей закладываются в виде латеральных утолщений соматоплевры примерно на 55-м часу развития. Эти почки вырастают из тела зародыша в виде выступов, покрытых эктодермой и заполненных мезодермальной тканью. По мере продолжения роста начинают проступать контуры конечности, будь то крыло или нога. Процесс формирования контуров сопровождается отмиранием клеток в ряде мезодермальных участков конечности. На ранних стадиях образования почки конечности участки некроза можно выявить с помощью некоторых прижизненных красителей; такие участки есть и на переднем, и на заднем краях почки, там, где она соединяется со стенкой тела, а также в центре почки. Эти три участка называются соответственно передней и задней некротическими зонами и темными пятнами. Передняя и задняя зоны обеспечивают формирование контуров проксимальных областей конечности, а темное пятно – отделение большой берцовой кости от малой в ноге и лучевой от локтевой – в крыле. Происходящее на более поздних стадиях развития конечности разделение пальцев сопровождается некрозом межпальцевых участков.
Сондерс и Фаллон (Saunders, Fallen), изучая заднюю некротическую зону (ЗНЗ) почки крыла цыпленка, выявили несколько интересных особенностей этой группы клеток. Максимальной протяженности ЗНЗ достигает на 96-м часу развития; к этому времени гибнет 1500–2000 клеток. Эти клетки фагоцитируются популяцией из почти 150 макрофагов. Если мезодерму проспективной ЗНЗ извлечь из почки крыла за 40 ч до того, как становится заметным некроз, и пересадить ее на боковую поверхность тела реципиента, она все равно гибнет и притом в те же сроки. Сроки некротизации и гибели заложены в самих клетках ЗНЗ и не зависят от возраста реципиента. Однако мезодермальные клетки, находящиеся в почке конечности, которая примыкает к эксплантированной ЗНЗ, не погибают. Поэтому можно думать, что клетки ЗНЗ обладают механизмом, получившим название «внутренние часы смерти». Эти часы запускают механизм гибели клеток на очень ранних стадиях развития конечности, после чего клетки идут по предначертанному им пути. Ход этих часов, однако, до некоторой степени обратим. Если ЗНЗ пересадить на дорсальную поверхность почки крыла, то ее клетки не гибнут. Причину этого изучали, выращивая клетки ЗНЗ вместе с различными другими фрагментами мезодермы. Выращивание мезодермы из ЗНЗ с соматической мезодермой приводит к такому же результату, как пересадка ее in vitro на боковую поверхность зародыша-реципиента: клетки ЗНЗ гибнут в надлежащие сроки. Если, однако, ЗНЗ культивировать с дорсальной мезодермой крыла, то она не гибнет. Создается впечатление, что мезодерма из ненекротической области крыла защищает ЗНЗ от гибели, останавливая «часы смерти». Это было подтверждено экспериментами, в которых клетки ЗНЗ сначала пересаживали на дорсальную поверхность почки крыла, а затем лишали их защитных воздействий дорсальной мезодермы крыла, удаляя их с места пересадки через разные сроки после того, как по «часам смерти» они должны были бы погибнуть. Будучи удалены с места пересадки, эти клетки погибали; выживали только те из них, которых содержали in vitro в условиях защиты не менее 6 дней. К этому времени «часы» совершенно останавливались и клетки ЗНЗ выживали, даже в отсутствие дорсальной мезодермы крыла. Таким образом, гибель клеток, происходящая в проксимальных частях крыла, представляет собой результат действия двух факторов: неких независимых от остального организма клеточных часов, запускаемых в определенной группе клеток, и местоположения этих клеток относительно остальной мезодермы конечности. Анализ гибели клеток в межпальцевых участках позволил выявить еще один уровень регуляции. Гибель клеток в этих участках лапы утки невелика, и в результате между вторым, третьим и четвертым пальцами образуются перепонки. В промежутке между первым и вторым пальцами, где происходит некроз, перепонки нет. Сондерс и Фаллон создавали химеры, соединяя мезодерму из почки ноги утки с эктодермой цыпленка и наоборот. Такие гибридные почки ноги они пересаживали затем на боковую поверхность тела куриного зародыша, служившего реципиентом, и наблюдали за некрозом межпальцевых клеток. В обоих случаях картина некроза соответствовала тому, что характерно для утки. Отсюда можно сделать вывод, что утиная эктодерма, подобно мезодерме дорсальной поверхности крыла, может подавлять некроз, наблюдаемый обычно при развитии ноги цыпленка. Однако эктодерма куриного зародыша не индуцирует некроз в утиной мезодерме. Возможно, поэтому, что регуляция некроза в межпальцевых участках происходит таким же образом, как в ЗНЗ; т.е. что это автономная особенность межпальцевой мезодермы, которая может проявиться или не проявиться в зависимости от среды, в которой окажутся презумптивные некротические клетки.
Гены, вступающие в действие на более поздних стадиях развития и в процессе роста
Ясно, что мутации генов, непосредственно определяющих морфогенетические пути, в особенности тех генов, которые действуют на ранних стадиях, могут вызывать чрезвычайно резкие изменения развития. Однако существуют также гены, действующие на более поздних стадиях, и, хотя эти гены оказывают влияние на общую морфологию данного организма, оно во многих случаях не влечет за собой явных вредных последствий. К таким генам относятся гены, контролирующие особенности роста организма после становления общей морфологии и органогенеза. Они были обнаружены по экспрессии мутаций, которые влияют на действие гормонов, что приводит к гигантизму или карликовости. Изменения формы (например, относительных размеров конечностей) могут быть результатом изменений характера роста, вызываемых плейотропными эффектами, сопровождающими основной гормональный эффект мутации. Изменения такого типа несомненно приводили к эволюционным изменениям, однако подобные поздние изменения развития не вызывали коренных перестроек морфологии. Несмотря ни на что, карликовый слон – это все-таки несомненно слон. Известна, однако, одна мутация, изменяющая размеры организма, которая демонстрирует пластичность процесса развития и заслуживает поэтому особого внимания. Это – мутация gt (giant) у Drosophila melanogaster. Этот сцепленный с полом рецессивный признак был впервые обнаружен Бриджесом (Bridges) и Габерчевски (Gaberchevsky) в 1928 г. Весь процесс развития дрозофилы – от оплодотворения до достижения половозрелости – обычно занимает 10 дней. У мутантов gt этот процесс продолжается на 2–5 дней дольше. Образующиеся при этом особи морфологически не отличаются от обычных мух, но они вдвое крупнее. Способ возникновения этого изменения представляет значительный интерес. Развитие мух gt протекает нормально на протяжении всего эмбрионального периода и вплоть до приближения личинок к концу третьего возраста. Однако в тот момент, когда нормальные особи окукливаются и начинают метаморфоз, особи gt продолжают оставаться в личиночном состоянии. Именно на личиночных стадиях и в начале стадии куколки происходит пролиферация имагинальных дисков – клеток, предназначенных для формирования тканей взрослой мухи, а от числа этих клеток зависят размеры имаго. У личинок gt в течение этого растянутого периода личиночного развития, по-видимому, происходит по меньшей мере одно дополнительное клеточное деление. Об этом можно судить потому, что те клетки личинки, которые политенны (например, клетки слюнных желез), участвуют по крайней мере в одном, а иногда и в двух дополнительных циклах синтеза ДНК. Личинки gt после этого 2–5-дневного добавочного периода роста образуют куколку, которая примерно вдвое крупнее обычной. Затем происходит метаморфоз и после несколько удлиненной стадии куколки появляется на свет морфологически нормальная взрослая особь двойного размера. Следовательно, насекомое способно регулировать свое развитие, так что, несмотря на явную дополнительную пролиферацию, предшествующую дифференцировке, никаких дополнительных элементов (органов) не образуется. В эволюции ранних событий онтогенеза у оболочников и низших хордовых. У этих организмов число и сроки делений дробления изменялись в зависимости от относительного значения личиночной стадии у разных видов.
Это привело к тому, что у разных видов доля массы раннего зародыша, участвующая в образовании специализированных личиночных органов, различна. Отсюда следует, что две совершенно различные морфогенетические системы способны согласованно произвести крупные изменения и создать вполне интегрированный организм. К другим генам, действующим в конце развития, относятся гены, регулирующие распределение продуцируемых пигментов по поверхности тела и их количество; эффекты изменчивости этих генов совершенно очевидны в природных популяциях большинства организмов. Хотя такого рода изменения несомненно имеют важное значение для процессов отбора, а тем самым и для эволюции, они, вероятно, несущественны для морфологических изменений per se.
Наш основной принцип заключается в том, что онтогенез находится под контролем генов. Как было показано, этот контроль осуществляется на нескольких уровнях. С помощью мутаций, наследуемых по материнскому типу, можно продемонстрировать генетическую регуляцию организации яиц у таких видов, как дрозофила, у которой раннее развитие имеет мозаичный характер. Другие мутации, такие как tailless у мыши или cardiac и eyeless у аксолотля, указывают на существование генетического контроля, действующего на последующих стадиях развития, точно так же как для надлежащего функционирования каскадных онтогенетических событий, приводящих к дифференцировке основных зародышевых листков, необходима специфическая генетическая информация, а для органогенеза – соответствующие индукционные взаимодействия. Наконец, генетические изменения могут изменять события, происходящие на поздних стадиях развития, в том числе процессы роста и пигментацию, а тем самым и дефинитивную форму взрослого организма. Эволюционный процесс может отбирать изменения экспрессии генов, играющих важную роль в развитии, и создавать таким образом новые морфогенетические пути. Необходимо, однако, подчеркнуть различие между генами этого класса и природой мутаций (изменений экспрессии), важных для эволюционного изменения. Большая часть обсуждавшихся в этой главе мутаций обладает резко выраженными и гибельными эффектами. Их значение состоит в том, что они позволяют выявить генетические элементы, лежащие в основе некоторых процессов развития. В морфологической же эволюции, по всей вероятности, участвуют мутации, обладающие менее ясно выраженным эффектом. Все мутации, изменяющие сроки или продолжительность событий или силу взаимодействий, будут приводить к эволюционной модификации путей развития. Для становления структуры и для морфогенеза решающее значение имеют также гены и мутационные изменения совершенно иного типа. Им посвящены остальные главы.
Похожие работы

... процессы репликации ДНК и деления клетки. Реальные механизмы регуляции репликации еще не расшифрованы. У эукариот определенную роль в регуляции репликации, по-видимому, играют гистоны. 4. Транскрипция генетической информации Этот этап биосинтеза белков состоит в «переписывании» информации, закодированной в ДНК, на олигонуклеотидную последовательность информационной РНК и осуществляется ...
... реактивности сохраняется в течение многих месяцев. У таких мышей при последующем введении специфического антигена доминирует продукция иммуноглобулинов, не имеющих идиотипа Т15. Нейроэндокринная регуляция иммунного ответа Уже давно известно, что стрессовые ситуации могут служить причиной подавления иммунных функций организма, например снижения его способности преодолевать инфекции. Имеются ...


... , прежде всего, необходимо изменить окружение ребёнка в семье, школе и создать благоприятные условия для коррекции симптомов расстройства и преодоления отставания в развитии высших психических функций [21]. Лечение детей с синдромом дефицита внимания с гиперактивностью должно включать комплекс методик, или, как говорят специалисты, быть «мультимодальным». Это значит, в нём надлежит участвовать ...
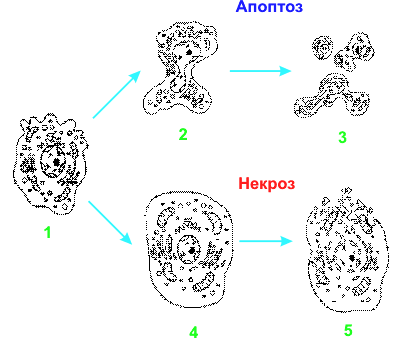
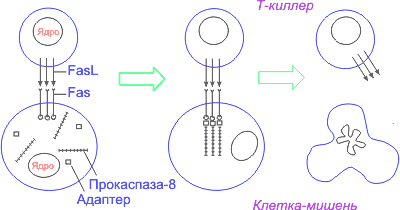
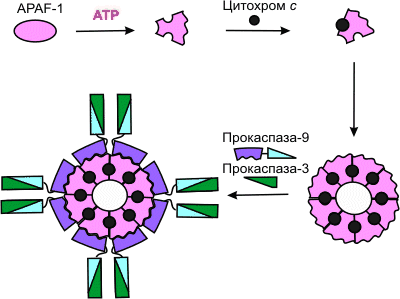
... и в конечном итоге достигает ядра, где и происходит включение программы клеточного самоубийства путем активации летальных и/или репрессии антилетальных генов. Однако существование ПКС (программируемая клеточная смерть) в безъядерных системах (цитопластах – клетках, лишенных ядра) показывает, что наличие ядра не является обязательным для реализации процесса ]. Применительно к клеткам животных и ...
0 комментариев