Навигация
Механизм действия гормонов. Роль циклазной системы в механизме действия гормонов
53954
знака
0
таблиц
4
изображения
2. Механизм действия гормонов. Роль циклазной системы в механизме действия гормонов
По механизму действия гормоны делят на два основные типа. Первый – это белковые и пептидные гормоны, катехоламины и гормоноиды. Их молекула, подойдя к клетке- мишени, соединяется с молекулами белковых рецепторов наружной плазматической мембраны, затем с помощью медиаторов (ц АМФ, ц ГМФ, простагландинов, Са2+) оказывает влияние на ферментные системы клетки- мишени и на обмен веществ в ней. К гормонам второго типа относят стероидные и часть тиреоидных гормонов. Их молекула легко проникает в глубь клетки- мишени через поры мембраны; взаимодействует с молекулами гликопротеидных рецепторов, локализированных в цитозоле, митохондриях на ядерной мембране, оказывая воздействие на весь клеточный метаболизм, и в первую очередь процессы транскрипции.
Механизмы действия гормонов на клетки-мишени.
В зависимости от строения гормона существуют два типа взаимодействия. Если молекула гормона липофильна, (например, стероидные гормоны), то она может проникать через липидный слой наружной мембраны клеток-мишеней. Если молекула имеет большие размеры или является полярной, то ее проникновение внутрь клетки невозможно. Поэтому для липофильных гормонов рецепторы находятся внутри клеток-мишеней, а для гидрофильных - рецепторы находятся в наружной мембране.
Для получения клеточного ответа на гормональный сигнал в случае гидрофильных молекул действует внутриклеточный механизм передачи сигнала. Это происходит с участием веществ, которых называют вторыми посредниками. Молекулы гормонов очень разнообразны по форме, а "вторые посредники" - нет.
Надежность передачи сигнала обеспечивает очень высокое сродство гормона к своему белку-рецептору.
Что такое посредники, которые участвуют во внутриклеточной передаче гуморальных сигналов?
Это циклические нуклеотиды (цАМФ и цГМФ), инозитолтрифосфат, кальций-связывающий белок - кальмодулин, ионы кальция, ферменты, участвующие в синтезе циклических нуклеотидов, а также протеинкиназы - ферменты фосфорилирования белков. Все эти вещества участвуют в регуляции активности отдельных ферментных систем в клетках-мишенях.
Разберем более подробно механизмы действия гормонов и внутриклеточных посредников.
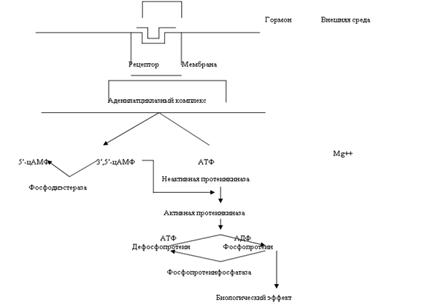
Существует два главных способа передачи сигнала в клетки-мишени от сигнальных молекул с мембранным механизмом действия:
аденилатциклазная (или гуанилатциклазная) системы;
фосфоинозитидный механизм.
Прежде чем выяснить роль циклазной системы в механизме действия гормонов, рассмотрим определение этой системы. Система циклазная – это система, состоящая из содержащихся в клетке аденозинциклофосфата, аденилатциклазы и фосфодиэстеразы, регулирующая проницаемость клеточных мембран, участвует в регуляции многих обменных процессов живой клетки, опосредует действие некоторых гормонов. То есть роль циклазной системы заключается в том, что они являются вторыми посредниками в механизме действия гормонов.
Система «аденилатциклаза - цАМФ». Мембраны фермент аденилатциклаза может находиться в двух формах - активированной и неактивированной. Активация аденилатциклазы происходит под влиянием гормон-рецепторного комплекса, образование которого приводит к связыванию гуанилового нуклеотида (ГТФ) с особым регуляторным стимулирующим белком (GS-белок), после чего GS-белок вызывает присоединение магния к аденилатциклазе и ее активацию. Так действуют активизирующие аденилатциклазу гормоны глюкагон, тиреотропин, паратирин, вазопрессин, гонадотропин и др. Некоторые гормоны, напротив, подавляют аденилатциклазу (соматостатин, ангиотензин-П и др.).
Под влиянием аденилатциклазы из АТФ синтезируется цАМФ, вызывающий активацию протеинкиназ в цитоплазме клетки, обеспечивающих фосфорилирование многочисленных внутриклеточных белков. Это изменяет проницаемость мембран, т.е. вызывает типичные для гормона метаболические и, соответственно, функциональные сдвиги. Внутриклеточные эффекты цАМФ проявляются также во влиянии на процессы пролиферации, дифференцировки, на доступность мембранных рецепторных белков молекулам гормонов.
Система «гуанилатциклаза - цГМФ». Активация мембранной гуанилатциклазы происходит не под непосредственным влиянием гормон-рецепторного комплекса, а опосредованно через ионизированный кальций и оксидантные системы мембран. Так реализуют свои эффекты натрийуретический гормон предсердий - атриопептид, тканевой гормон сосудистой стенки. В большинстве тканей биохимические и физиологические эффекты цАМФ и цГМФ противоположны. Примерами могут служить стимуляция сокращений сердца под влиянием цАМФ и торможение их цГМФ, стимуляция сокращений гладких мышц кишечника цГМФ и подавление цАМФ.
Кроме аденилатциклазной или гуанилатциклазной систем существует также механизм передачи информации внутри клетки-мишени с участием ионов кальция и инозитолтрифосфата.
Инозитолтрифосфат - это вещество, которое является производным сложного липида - инозитфосфатида. Оно образуется в результате действия специального фермента - фосфолипазы "С", который активируется в результате конформационных изменений внутриклеточного домена мембранного белка-рецептора.
Этот фермент гидролизует фосфоэфирную связь в молекуле фосфатидил-инозитол-4,5-бисфосфата и в результате образуются диацилглицерин и инозитолтрифосфат.
Известно, что образование диацилглицерина и инозитолтрифосфата приводит к увеличению концентрации ионизированного кальция внутри клетки. Это приводит к активации многих кальций-зависимых белков внутри клетки, в том числе активируются различные протеинкиназы. И здесь, как и при активации аденилатциклазной системы, одной из стадий передачи сигнала внутри клетки является фосфорилирование белков, которое в приводит к физиологическому ответу клетки на действие гормона.
В работе фосфоинозитидного механизма передачи сигналов в клетке-мишени принимает участие специальный кальций-связывающий белок - кальмодулин. Это низкомолекулярный белок (17 кДа), на 30 % состоящий из отрицательно заряженных аминокислот (Глу, Асп) и поэтому способный активно связывать Са+2. Одна молекула кальмодулина имеет 4 кальций-связывающих участка. После взаимодействия с Са+2 происходят конформационные изменения молекулы кальмодулина и комплекс "Са+2-кальмодулин" становится способным регулировать активность (аллостерически угнетать или активировать) многие ферменты - аденилатциклазу, фосфодиэстеразу, Са+2,Мg+2-АТФазу и различные протеинкиназы.
В разных клетках при воздействии комплекса "Са+2-кальмодулин" на изоферменты одного и того же фермента (например, на аденилатциклазу разного типа) в одних случаях наблюдается активация, а в других - ингибирование реакции образования цАМФ. Такие различные эффекты происходят потому, что аллостерические центры изоферментов могут включать в себя различные радикалы аминокислот и их реакция на действие комплекса Са+2-кальмодулин будет отличаться.
Таким образом, в роли "вторых посредников" для передачи сигналов от гормонов в клетках-мишенях могут быть:
циклические нуклеотиды (ц-АМФ и ц-ГМФ);
ионы Са;
комплекс "Са-кальмодулин";
диацилглицерин;
инозитолтрифосфат.
Механизмы передачи информации от гормонов внутри клеток-мишеней с помощью перечисленных посредников имеют общие черты:
одним из этапов передачи сигнала является фосфорилирование белков;
прекращение активации происходит в результате специальных механизмов, инициируемых самими участниками процессов, - существуют механизмы отрицательной обратной связи.
Гормоны являются основными гуморальными регуляторами физиологических функций организма, и в настоящее время хорошо известны их свойства, процессы биосинтеза и механизмы действия. Гормоны являются высокоспецифичными веществами по отношению к клеткам-мишеням и обладают очень высокой биологической активностью.
Похожие работы
... за пределами ЦНС в ряде эндокринных желез, а также в других органах и тканях. В онтогенезе регуляторные пептиды появились значительно раньше «классических» гормонов, т.е. до обособления специализированных эндокринных желез. Это позволяет считать, что в геноме запрограммировано раздельное образование названных групп веществ, а следовательно они являются самостоятельными. Источниками регуляторных ...
... обратного Т3 у новорождённых резко повышено, но в течение первых недель уровень трийодтиронинов достигает пропорции, характерной для взрослых. Пониженное содержание гормонов щитовидной железы у плода и новорожденного приводит к развитию кретинизма - заболевания, которое характеризуется множественными нарушениями и тяжелой необратимой задержкой умственного развития. При возникновении гипотиреоза ...
... продукт, мы тем самым изъявляем свою волю потребителя, с которой промышленности приходится считаться. Именно воля потребителей заставила европейских политиков принять законы о запрете применения гормонов в животноводстве. Покупая, мы одобряем и поддерживаем, а отказываясь покупать, — осуждаем. Сейчас в Америке растет число людей, которые стали вегетарианцами не только ради своего здоровья, а в ...
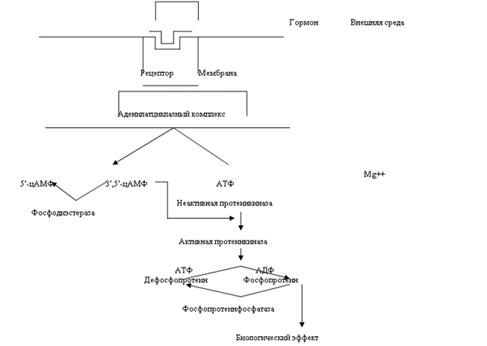
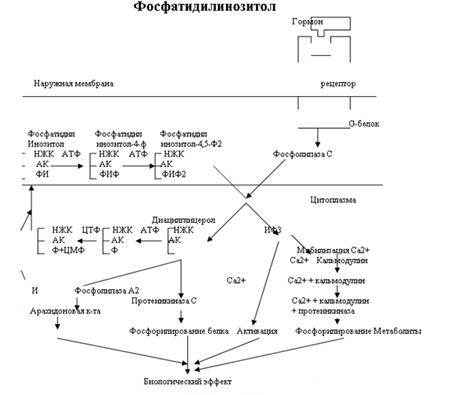
... эффект опосредуется связыванием ГР-РФ с рецепторами плазматической мембраны соматотрофов и активацией трех систем вторичных посредников. С их помощью происходит усиленный синтез мРНК СТГ. Этот эффект действия водорастворимого гормона на ядерный аппарат клетки объясняют фосфорилированием и дефосфорилированием определенной группы протеинкиназ, которые действуют на генетический аппарат клетки. На ...
0 комментариев