Вступ
Живлення рослин відіграє важливу роль у вирішенні однієї з головних задач фітофізіології - розкритті механізмів продуктивного процесу на загальному тлі інтенсифікації рослинництва з метою теоретичного обґрунтування інтенсивних технологій вирощування основних сільськогосподарських культур. Принципово нові технології базуються також на використанні нових видів добрив, створених на полімерній основі з програмованим вивільненням живильних речовин. Оптимальне живлення рослинних організмів у сполученні з раціональним підвищенням ефективності застосування добрив і зменшенням забруднення навколишнього середовища продуктами хімізації дозволить підвищити врожай і поліпшити якість сільськогосподарської продукції.
Мета даної роботи сформувати поняття про грунт як активне поживне середовище кореневого живлення рослин та про процеси, які впливають на активність живлення.
Предмет дослідження грунт та кореневе живлення рослин.
Об’єкт дослідження процес кореневого живлення рослин.
Завдання даної роботи:
1) розглянути грунт та фактори, що впливають на його властивості;
2) виявити мінеральні та металорганічні сполуки в рослинах;
3) визначити місце кореневого живлення в житті рослин;
4) встановити основні функції кореневої системи;
Методологічною основою даної роботи, є діалектичний матеріалізм, що виходить із уявлень про життя як особливу форму руху матерії. Застосування законів діалектики як метод пізнання у фізіологічних дослідженнях живлення рослин відкрило широкі практичні перспективи.
1 Ґрунт – активне середовище живлення
1.1 Ґрунт як поживний субстрат рослин
Ґрунт є поживним субстратом рослин: у ньому міститься головний запас потенційної біогенної енергії у вигляді коренів рослин, біомаси мікроорганізмів і гумусу. Без надходження з ґрунту таких елементів як фосфор і калій, було б неможливе створення первинної рослинної продукції.
У процесі ґрунтоутворення відбувається руйнування мінералів породи і витяг елементів, що надходять потім в обмінні реакції біосинтезу. В основі розташування мінералів у ґрунтах лежать наступні процеси: розчинення сильними мінеральними кислотами, виділюваними коренями і життєдіяльними мікроорганізмами; вплив органічних кислот - продуктів шумувань і неповних окислювань вуглеводів грибами; взаємодія з позаклітинними амінокислотами, полісахаридами, фенольними сполуками. Органічні сполуки прямо чи побічно взаємодіють з мінералами, руйнуючи кристалічні ґратки, утворити комплексні сполуки, переводячи елементи з однієї форми в іншу зі зміною валентності і рухливості.
Розглянемо біохімічну деструкцію мінералів на прикладі калію, вміст якого в доступній формі в ґрунті нижче за потреби в ньому рослин. Засвоюваний калій складає всього 1-2% його загальної кількості в ґрунті. Основний запас калію знаходиться в мінералах і в складі органомінеральних комплексів. Первинні мінерали, що містять калій, - слюди (біотит і мусковіт) і польові шпати (ортоклази і мікрокліни). Калій входить і у вторинні мінерали: каолін, монтморилоніт, вермикуліт. Звільнення калію з мінералів здійснюється в процесі біологічного вивітрювання, у результаті якого відбувається або повне розчинення мінералу з утворенням аморфних продуктів розпаду, або ізоморфне заміщення іонів мінералу іонами водню і натрію без руйнування кристалічних ґрат мінералу.
Хімічні елементи, що входять до складу мінералу, витягаються необов'язково пропорційно їхньому вмісту і співвідношенню у вихідному матеріалі. Біологічне вивітрювання може привести до перетворення одного мінералу в інший завдяки застосуванню хімічного складу при виборчому витягу елементів. Наприклад, при розкладанні алюмосилікатів за участю гетеротрофних бактерій відбувається послідовний витяг спочатку лужних елементів, потім лужноземельних і в останню чергу - кремнію й алюмінію. У природі найбільш інтенсивна деструкція мінералів протікає в підзолистих ґрунтах.
Стійкість мінералів до біологічного руйнування визначається не тільки специфічністю мікрофлори, міцністю структури кристалічних ґраток, але й умовами середовища. Мікроорганізми ґрунту беруть участь не тільки в розсіюванні елементів, що містяться у мінералах, але й у мінералоутворенні. Наприклад, вони беруть участь у відкладенні сульфідних, карбонатних, фосфатних, залізистих і силікатних мінералів. Кальцити утворяться при осадженні кальцію вуглекислотою, яка виділяється при диханні, бродінні і неповному окисному розкладанні органічних речовин анаеробними сульфатвідновлюючими бактеріями, аеробними дріжджами і псевдомонадами.
Нагромадження відмерлих рослинних залишків у вигляді особливого шару підстилки, чи повсті, на поверхні ґрунту створює особливе сховище елементів харчування, що у міру розкладання поступово надходять у ґрунт. На поверхні твердих ґрунтових часток зосереджені основні запаси живильних речовин: гумус, органомінеральні колоїди, катіони кальцію, магнію й ін. Їхня концентрація тут значно вище, ніж у ґрунтовому розчині. Серед органічних речовин ґрунтового розчину є вітаміни, ферменти й інші продукти життєдіяльності коренів і мікроорганізмів. У ґрунтовому розчині підзолистих і болотних ґрунтів переважають органічні речовини, а в чорноземах приблизно рівне співвідношення органічних і мінеральних речовин. У каштанових ґрунтах і сіроземах, як правило, більше мінеральних речовин, чим органічних.
З мінеральних речовин ґрунту в мінімумі звичайно знаходяться азот і фосфор, а калій входить до складу твердої частини ґрунту.1
Органічні і мінеральні речовини ґрунту або використовуються рослинами і мікроорганізмами, або акумулюються і стабілізуються у формі гумусових речовин, що визначають ґрунтову родючість. Гумус складає до 90 % загального запасу органічних речовин у ґрунтах і представлений групою високомолекулярних сполук різної хімічної природи, головним чином високополімерних органічних кислот (гумінові і фульвокислоти), полісахаридів, білкових речовин.
До складу гумусу входить майже весь азот ґрунту, біля половини фосфору і 60-90 % сірки. Установлено, що в гумусі є кальцій, магній, залізо, а також фосфор у доступній для рослин формі.
Гумус сильно варіює як по якості, так і по кількості в ґрунтах різних типів внаслідок того, що він утворюється з рослинних залишків неоднакового хімічного складу й у різних умовах, а в його формуванні бере участь комплекс ґрунтових організмів.
Біохімічна концепція гумусоутворення зводиться до наступного. Мікробне розкладання рослинних залишків супроводжується втратою маси (до 75%) і виділенням СО2. Першоджерела структурних одиниць гумусових речовин - вуглеводи рослинних залишків, перероблені мікроорганізмами, фенольні речовини, а також продукти мікробного ресинтезу, які містять азот. У процесі формування гумусу відбувається конденсація структурних одиниць за участю мікробних (головним чином, грибних) поліфенолоксидаз. У кінцевих процесах мають місце гетерополіконденсація, стабілізація за рахунок ізомеризації і перегрупувань. Гумінові кислоти, як гетерополіконденсати, містять велике число фенольних і індольних одиниць.
На мінеральне харчування рослин впливають кореневі мікроорганізми, що розділяються на ризпланові, клубенькові і мікоризоутворювачі. На поверхні кореня в ризоплані чисельність мікроорганізмів, здатних фіксувати азот і проводити денітрифікацію, у тисячі разів більша, ніж у ґрунті. Завдяки слизуватим виділенням кореня, що отримали назву міцелію, мікроорганізми утворять на поверхні кореня мікроколонії, а в деяких місцях - майже суцільні плівки. Ризопланові бактерії, асоційовані з коренем, і клубенькові бактерії впливають на азотний баланс рослини.
Гриби утворять на коренях рослин обростання, названі мікоризою. В даний час установлено наявність мікоризи в 80 % рослин: у всіх голонасінних і в 78 % покритонасінних. Рослини з мікоризою зустрічаються у всіх природних зонах, за винятком полярних пустель і високогір'їв. Завдяки мікоризі корені краще поглинають із ґрунту вологу і мінеральні елементи. Особливо важлива роль мікоризи в постачанні рослин доступними формами фосфору. Мікроорганізми не тільки вступають у безпосередній контакт із коренем, але і, оббита в зоні його дії, можуть так чи інакше впливати на життєздатність рослинного організму. Та частина ґрунтового середовища, що примикає до кореня і випробує вплив кореневих виділень, називається ризосферою.
Отже, при наявності гумусу і життєздатних мікроорганізмів у ґрунті рослини можуть більше використовувати мінеральних елементів живлення. Значна частина поживних речовин ґрунту, що іде з врожаєм, випадає з малого біологічного кругообігу.
Тому забезпечення бездефіцитного балансу живильних речовин у ґрунті - важлива задача регулювання живильного режиму рослин.
1.2 Вміст мінеральних елементів у рослинах
Усереднені результати аналітичних досліджень показують, що близько 1 % мінеральних солей приходиться на 75 - 80 % води. Ці величини в перерахуванні на молекулярні співвідношення можна представити так: на 105 молекул води - 500 - 600 молекул неорганічних речовин. Метали, як складова частина золи, розподіляються в клітках рослин нерівномірно. Нижче вказуються зведення про наявність металів у клітинних структурах (по Е. А. Бойченко, 1977): клітинні стінки – Si, Са , іноді Мg, Аl; ядра- Са, Мg, Nа, K, Fe, Zn, Cu; хлоропласти - Мg, Са, К, Nа, , Fe, Zn, Cu; Мо; мітохондрії - Са, Мg, К, Nа, , Fe, Zn, Cu; рибосоми - Мg, Са, Мn; вакуолярный сік - Nа, Мg, K, Са.
Виходячи з кількісного змісту мінеральних елементів у тканинах рослин, їх прийнято поділяти на макроелементи (С, Н, 0, N, S, Р, K, Мg, Са, Fе) і мікроелементи (B, Мn, Си, Zn, Мо, Co, Li). Залізо знаходиться на границі між макро- і мікроелементами. Макроелементи поєднують у групу елементи, вміст яких виражається 101- 10-2, а мікроелементи - зміст яких коливається в межах 10-3 – 10-5.1
Якісний склад золи залежить від вмісту мінеральних речовин у ґрунті й умов зовнішнього середовища. Як правило, чим багатший ґрунт і чим сухіше клімат, тим вище вміст золи в рослинах. Будь - який хімічний елемент, наявний у даному місцеперебуванні, може бути виявлений і в рослині.
Приведемо середній елементний склад рослин у % на сиру речовину (по Д Б. Вахмистрову, 1979): кальцій - 0,4; калій -0,25; азот - 0,25; кремній - 0,2; фосфор - 0,06; сірка - 0,04; магній -0,03; натрій - 0,02; хлор - 0,02; залізо - 0,001; марганець - 0,001; бор - 0,001; цинк - 0,0005; мідь - 0,0002; молібден - 0,00001.
Таким чином, хімічний склад рослини не відбиває його потреби в живильних речовинах.
Кількість золи в різних частинах рослини, а також у різних рослинах неоднаково. З'ясовано, що більше всього золи міститься в тканинах рослин, що складаються переважно з живих кліток. Найменше золи (близько 1 %) міститься в деревині, у насіннях - 3 %, коренях, стеблах - 4-5 %. Уже ці цифри показують, що зольні елементи зосереджені в тих органах, рівень життєдіяльності яких досить високий.
Вміст золи й окремих елементів у рослинах коливається в широких межах, у залежності від виду рослин. Так, у складі листів картоплі 5-13% золи, а буряка - 11-15%. За даними А. М. Гродзинського (1979), у середньому при врожаї яблук 100 ц/га з ґрунту щорічно виноситься, г: магнію - 1022, кальцію -854, заліза - 45, міді - 3,8, марганцю - 3,2, молібдену - 0,05.
Процентний уміст зольних елементів і азоту різко падає в міру старіння рослин. Дані по виносі мінеральних елементів ґрунту свідчать про те, що озима пшениця накопичує мінеральні елементи до фази молочної спілості, при цьому особливо інтенсивно вони поглинаються в період від кущіння до молочної стиглості.
Вміст того самого елемента в тканинах і клітках рослини дуже мінливий. Наприклад, якщо в листах і стеблах у складі їхньої золи основна частка приходиться на кальцій, то в насіннях, напроти, кальцію, як правило, менше, ніж калію і фосфору.
Розподіл елементів у клітинних структурах дуже по-різному, що можна проілюструвати даними на прикладі хлоропластів (табл. 2). Таблиця показує нагромадження елементів у пластидах. Такі елементи, як калій, кальцій, магній і фосфор, знаходяться в різних співвідношеннях, причому переважаючим є калій. Магній, залізо, мідь і цинк показують визначену локалізацію в пластидах, що, безсумнівно, знаходиться в зв'язку з роллю, що вони виконують у процесі фотосинтезу.
Відомо, що магній є складовою частиною молекул хлорофілу й атом цього елемента сприяє утворенню агрегатів молекул хлорофілу, що полегшує уловлювання світла.
Залізо хоча і не входить до складу хлорофілу, однак зовсім необхідно для його утворення. За даними А. С. Вечора (1961), близько 80 % заліза всієї листової тканини шпинату і люцерни знаходиться в хлоропластах. Крім того, велику роль у функціонуванні цих пластид відіграють залізовмісні ферменти (каталаза, пероксидаза, цитохроми). Висока активність залізовмісних ферментів у пластидах служить підтвердженням біокаталітичної природи заліза, що знаходиться в пластидах.
До ферментів, що містять цинк відноситься карбоангідраза хлоропластів, яка приймає участь в асиміляції і виділенні вуглекислоти рослинами. Установлено стимулюючий вплив міді на нагромадження хлорофілу.
Відзначено таку закономірність: мінеральні речовини, що не приймають участі у реутилізаці (заліз, бор, цинк), мають базипетальний градієнт вмісту, тобто кількість їх збільшується від вершин до основи органа, а мінеральні речовин, здатні реутилізуватися (азот, фосфор, сірка), - акропетальний, тобто кількість їх зростає від основи до верхівки.1
Мобільність різних іонів широко варіює: високорухливі - калій, натрій, фосфор, сірка; іони із середньою рухливістю -магній, залізо, мідь, цинк; відносно рухливі - кальцій, літій, бор. Однак рухливість іонів проміжної групи залежить від кількості елемента в рослині. Наприклад, мідь і цинк рухливі тільки при високій концентрації в ґрунті.
Отже, величина виносу мінеральних елементів надземними органами значно коливається під впливом грунтово-кліматичних умов, агротехніки оброблення культур і залежить від сортової специфікації і фази розвитку рослин. В даний час найбільш з'ясована потребу рослин у трьох елементах мінерального харчування - азоті, фосфорі і калії.
Принципи добору природою біометалів для побудови біомаси дотепер не з'ясовані, хоча зовсім очевидно, що їх розповсудженість не була вирішальним критерієм. Так, алюміній і титан - досить розповсюджені в природі елементи, однак вони не є елементами життя. Навпроти, молібден, що рідко зустрічається в природі елемент, є вкрай необхідним в організмі для процесів, зв'язаних, у першу чергу, з переносимо електронів.
1.3 Металорганічні сполуки рослинБагато біометалів (Fе, Мп, Си, Zn, Мо і Co) у рослинах взаємодіють з макромолекулами. Такі макромолекули прийнята називати біолігандами. До природних лігандів варто віднести білки, нуклеїнові кислоти і вітаміни. Координаційні сполуки характеризуються насамперед координаційним числом, тобто числом атомів, що складають найближче оточення центрального атома - біометалу (атома комплексоутворювача). Метал і його найближче оточення складають внутрішню сферу координаційного з'єднання. Донорні атоми ліганду розміщаються в просторі у чітко визначеному порядку, утворюючи при цьому геометричну фігуру. Можливість утворення комплексів полідентатними лігандами приводить до різкого зміцнення комплексів у порівнянні з тими, у яких хелатний ефект не спостерігається. Це положення може бути проілюстроване на прикладі координації іона металу в макромолекулі нуклеїнових кислот.
Можливо монодентатне приєднання іона металу до негативно зарядженого атома кисню у фосфатній групі і до вільних атомів азоту гетероциклів, а також - до атомів чи азоту кисню, що беруть участь в утворенні водневих зв'язків. Бідентатне (хелатное) приєднання іонів металів може здійснюватися за участю різних атомів нуклеотиду з утворенням п’ятичленних циклів. Але найбільш активними координуючими групами нуклеїнових кислот є фосфатні.
Отже, електродонорні атоми азоту і кисню гетероциклічних основ, гідроксильні групи рибози нуклеозидов і фосфатні групи нуклеотидів є потенційними центрами зв'язування іонів металів.
Іони металів, приєднані до таких центрів у полінуклеотидах, роблять на них різкий вплив, викликаючи лігандні реакції, що сильно змінюють структуру макромолекул.
Участь функціональної групи в зв'язуванні металу залежить від двох факторів, а саме: наскільки успішно функціональна група конкурує з іншими сусідніми і наскільки успішно іони металів конкурують із протонами за потенційно донорні атоми.
Теоретичні досягнення в області вивчення структури і функції біокомплексів знайшли практичне застосування в рослинництві. Зокрема, розроблений метод боротьби з вапняним хлорозом рослин. Унаслідок високого значення рН вапняних ґрунтів багато елементів мінерального харчування рослин, у тому числі і залізо (III), утворять малорарозчинні карбонати і гідроксиди, недоступні для засвоєння рослинами. Природно, що за таких умов добавка з'єднань заліза в ґрунт не дає належного ефекту, оскільки вони' негайно ж виводяться з розчинної фази ґрунту внаслідок утворення гідрооксиду.
Застосування полідентатних реагентів типу комплексонів створило цілком реальну можливість для перекладу заліза в стійкий, розчинний і проникаючий крізь клітинну мембрану комплекс заліза (III). З цією метою був застосований етилендіамінтетраацетат (ЕДТА), ефект від якого спостерігався лише на помірно кислих ґрунтах. Комплекси заліза з ЕДТА в лужному середовищі виявилися недостатньо стабільними унаслідок випадання осаду гідроксиду заліза.
Початковий добір необхідних металів для життєдіяльності клітини мав місце, мабуть, на стадії утворення біосфери в гідросфері Землі. Багато елементів могли виконувати роль каталізаторів у вигляді вільних іонів. Результатом еволюції функцій з'явилося ускладнення структури за рахунок утворення комплексів з органічними сполуками, що дало початок у першу чергу ферментним системам. У зв'язку з еволюцією автотрофного способу живлення рослинних організмів збільшувалася частка участі металорганічних комплексів у внутрішньоклітинному метаболізмі. Згідно з даними Е.А.Бойченко (1977), зміни металів в еволюції функцій можна представити в такий спосіб:
Поліфункціональність неорганічних іонів виявилася, мабуть, основним критерієм добору їх у процесі хімічної еволюції. Основна роль цих іонів - окислювально-відновна.
Окисники і відновники завжди функціонують як сполучені окислювально-відновні пари (редокс-пари), подібно тому, як кислоти і основи функціонують як сполучені кислотно-основні пари.
Ланцюг переносу електронів може включати велике число переносників, які можна розглядати як функціональні ансамблі, що діють у визначеній послідовності.
Особливий інтерес представляє негеміновое і геміновое залізо. На його прикладі добре просліджується удосконалювання окислювально-відновних систем. Синьо-зелені водорості, що широко поширилися ще в період докембрію і виявилися першими фотосинтетиками, що виділяли кисень, містять найбільша кількість заліза (до 1 % на суху речовину). Як затверджує Е. А. Бойченко й ін. (1979), у ході еволюції синьо-зелених водоростей відзначається поступове зростання частки інших металів щодо заліза. Зростання окисних процесів у клітках зв'язано з нагромадженням у них кобальту, нікелю і пізніше міді. З цією тенденцією gjв'язане подальше виникнення нових ферментів і включення додатково до заліза другого металу в молекулі таких ферментів, як гідрогенази (Fе і Мо; Fе і Мn), редуктаза вуглекислоти (Fе і Мо), ксантиноксидаза (Fе і Мо), цитохромоксидаза (Fе і Си). У результаті утворення в білках двохядерних комплексів за участю металів зростають активність і ефективність каталізу. Еволюція окислювально-відновних систем відбувалася головним чином у напрямку можливості окислювання речовин з більш високим окислювально-відновним потенціалом.
Координація органічних молекул з іонами металів істотно змінила їхню реакційну здатність. У результаті приєднання їх до позитивно зарядженого іона металу електронна щільність зміщається в напрямку цього іона. Поляризація ліганду, як правило, збільшує кислотність атомів водню при донорному атомі, полегшує атаку ліганду нуклеофілами й утрудняє атаку електрофілами (відзначимо, що атоми чи групи атомів, що оточують центральний атом металу, називаються лігандами, а атоми, безпосередньо приєднані до металу, називаються донорними атомами).1
Центральним питанням мінерального живлення рослин є азотне живлення, що, як відомо, є найбільш лімітуючим продуктивність рослин. Азот разом з вуглецем, киснем і воднем утворить групу так званих елементів-органогенів. Кількість азоту в складі сухої речовини рослин невелика - вона звичайно коливається від 1 до 3 %. У середовищі, що оточує рослину, азот знаходиться в двох формах: у вигляді газоподібного азоту атмосфери, що складає 78,2 % повітря (за обсягом), і у виді різних органічних і неорганічних сполук, пригнічуюча частина яких зосереджена в ґрунті, морях і океанах. Понад 99% азоту ґрунту знаходиться в недоступній для рослин органічній формі.
Рослинам недоступний величезний запас газоподібного азоту, що знаходиться в повітрі, а з великої розманітності сполук азоту, що зустрічаються в ґрунті, вони можуть використовувати для харчування в основному тільки мінеральні форми цього елемента. Тому питання про перетворення з'єднань азоту в ґрунті під впливом мікроорганізмів залучає до себе велика увага. Більшість таких перетворень здійснюється шляхом нітрифікації, денітрифікації, амоніфікації, іммобілізації й азотфіксації.
2 Родучість ґрунту та фактори, що на неї впливають
Основним джерелом зольного живлення рослин є мінеральні елементи, що містяться в ґрунті, і азот. Багатство ґрунту мінеральними речовинами визначається специфічними особливостями материнської гірської породи і діяльністю мікроорганізмів.
Велику роль в утворенні самого ґрунту і нагромадженні у верхніх її обріях мінеральних елементів виконують у процесі своєї життєдіяльності і самі рослини. Поглинаючи своїми коренями мінеральні речовини, рослини піднімають їх у верхні шари ґрунту і тим самим збагачують останні.
Умови мінерального живлення в значній мірі залежать від типу ґрунту. Тому при виявленні потреб рослини в мінеральному добриві необхідно виходити як з видових особливостей організму, так і зі специфічних особливостей даного типу ґрунту.
У багаторічних дослідженнях вітчизняних ґрунтознавців постійно підкреслювалася думка, що правильне уявлення про ґрунтове живлення рослин можна одержати лише розглядаючи ці процеси як одне з ланок загальбіологічного круговороту речовин у природі.
В. Р. Вільямі, серед факторів, що забезпечують високий рівень родючості ґрунтів, висував на перше місце міцну мілкокомкову структуру.:
Характер взаємодії рослини і ґрунту в значної степені пов'язаний із властивим ґрунтам так званої поглинаючої здатності, під якою розуміють властивість ґрунту поглинати, зв'язувати різні хімічні сполуки. Основний внесок у навчання про поглинальну здатність вніс один з видатних представників вітчизняної науки про хімію ґрунту Костянтин Каетанович Гедройц. Для різних типів ґрунтів їм розроблена детальна характеристика поглинаючою здатністю, рівня і ємності поглинання, складу поглинених катіонів. Ці дослідження показують існування зв'язку між агрономічними властивостями ґрунту, рівнем її родючості і складом поглинених катіонів.
К. К. Гедройц розрізняв наступні види поглинання:
а) механічне, б) фізичне, в) фізико-хімічне, або обмінне, г) хімічне, д) біологічне.
Усі перераховані види поглинання здійснюються комплексом, що складається з цеолітної (вірніше — цеолітоподібної, чи неорганічної) і гуматної (органічної) складових частин ґрунту. Весь комплекс перерахованих складових частин ґрунту Гедройц назвав ґрунтовим поглинаючим комплексом.
Найбільше значення має фізико-хімічне поглинання, сутність якого полягає в тому, що частина катіонів ґрунту, що містяться в її твердих частках, може бути виміняна на еквівалентну кількість катіонів, що знаходяться в ґрунтовому розчині або внесених у грунт у вигляді добрив. Ґрунт здатний поглинати не тільки катіони, але і деякі аніони, наприклад аніон фосфорної кислоти.1
Поглинаюча здатність різних ґрунтів неоднакова. Вона, зокрема, залежить від ступеня роздробленості (дисперсності) органічних і мінеральних сполук, що містяться в ґрунті. Особливо велике значення має багатство ґрунту органічною речовиною, що володіє досить високою здатністю до поглинання (адсорбції) катіонів. Поглинаюча здатність ґрунтових часток стосовно того чи іншого катіона залежить також від ступеня насиченості цим катіоном колоїдів ґрунту; Чим нижче зміст катіона в поглинаючому комплексі ґрунту, тим з більшою швидкістю «силою цей катіон адсорбується ґрунтовими колоїдами.
Склад поглинених іонів у різних ґрунтів різний. У число їх входить і водень, витіснення якого може спричинити підвищення кислотності 'ґрунту. У поглиненому стані може знаходитися й алюміній, що, будучи витиснений, має на рослину токсичний вплив і т.д.
Звідси видно, що фізико-хімічна, чи обмінна, поглинаюча здатність ґрунту є одним з активних регуляторів взаємодії між добривом і ґрунтом. У силу цього поглинальна здатність ґрунту робить безсумнівний вплив на здатність рослин використовувати внесене в даний ґрунт добриво. Поглинаючий комплекс грунту відіграє важливу роль як регулятор реакції ґрунту. Від особливостей поглинаючого комплексу в значній ступені залежить і буферність ґрунту, тобто її здатність протистояти зміщенням реакції.
Було б неправильно однак, зв'язувати всю сукупність властивостей ґрунту, і першу чергу її родючість, із властивостями й особливостями її поглинаючого комплексу.
Одним з факторів, від яких залежить рівень родючості ґрунту і, отже, весь комплекс умов мінерального живлення рослин, є ґрунтова мікрофлора.
Виняткове значення для з'ясування ролі мікроорганізмів у створенні родючості ґрунту мають класичні дослідження засновника ґрунтової мікробіології Сергія Миколайовича Виноградського. Особливу роль зіграли роботи Виноградского по азотфіксуючих і бактеріях, що нітрифікують, "а також по сіро- і залізобактеріям. Ці дослідження є основою сучасних уявлень про кругообіг речовин у природі.
В даний час можна вважати встановленим, що в процесі еволюції відбувся добір визначених видів і асоціацій мікроорганізмів, у взаємодії з який здійснюється кореневе живлення вищих рослина. У цю группу мікроорганізмів входять різні гриби, бактерії, водорості.
Мікроорганізми - симбіонти концентруються в основному в зоні розташування кореневих систем вищих рослини (так називана ризосфера). Взаємодія між ними і рослинами здійснюється або шляхом «роздільного симбіотрофізму», тобто під час відсутності безпосереднього контакту між тканинами коренів і мікроорганізмів, або на основі «щирого симбіотрофізму» — в умовах безпосередньої, інтимної асоціації партнерів.
Симбіози коренів вищих рослин із грибами звуться мікориз, симбіози з бактеріями — бактеріориз. У залежності від форми симбіотрофізму розрізняють мікоризи екто - і ендотрофні. Перші розвиваються на поверхні коренів, другі проникають усередину тканини. Класичним прикладом бактеріотрофізму є взаємодія бобових рослин із клубеньковими бактеріями.
Дослідження мікробіологів, проведені в останні роки, значно розширили представлення про значення симбіотрофізму в процесах кореневого живлення рослин.
Важливе значення серед продуктів життєдіяльності бактерій мають вітаміни — речовини, що володіють високою біологічної активністю. В даний час достовірно встановлено наявність у ґрунті вітамінів групи В (В1 В2, В6, В12), а також біотину, пантотенової, нікотинової, фолевої, параамінобензойної кислот і інших органічних кислот, мезоінозиту. Поряд з цими виявлені різні ферменти, а також ряд антибіотичних і токсичних речовин.
Кількість вітамінів і інших біологічно активних сполук залежить від загального .рівня родючості ґрунту, воно тим вище, чим вище вміст у ґрунті органічних речовин.
Отже, роль ґрунтових мікроорганізмів у житті. вищих рослин визначається не тільки їхньою здатністю мінералізовувати органічну речовину, але і здатністю синтезувати різноманітні органічні сполуки.
За характером їхні відношення до кисню ґрунтові мікроорганізми поділяються на дві групи:
а) анаеробні організми, життєдіяльність яких протікає без використання вільного кисню;
б) аеробна, здійснююча своя діяльність лише при обов'язковій участі молекулярного кисню. Ця група представлена численними видами бактерій, грибів і актиноміцетів.
Розкладання органічних речовин, результатом якого є утворення більш простих сполук, займає одне з центральних місць в життєдіяльності ґрунтової мікрофлори. Не менш суттєва роль належить цим процесам у створенні структури ґрунту.
Питання про хімічні шляхи, яким ці процеси здійснюються, складний, і не може вважатися дозволеним. Характер цих процесів залежить від наявних у ґрунті умов, хімічного складу рослинних залишків, фізіологічних властивостей мікроорганізмів і інших факторів.
У результаті всієї сукупності біологічних і хімічних процесів у ґрунті створюється складний комплекс органічних речовин, що поєднується терміном гумус.1
Кінцевий склад гумусу в значній мірі залежить від хімічного складу підлягаючих розпаду органічних сполук, а також від фізіологічної природи мікроорганізмів, що беруть участь у перетворенні цих речовин.
Саме тому виявилося настільки складним рішення питання про хімічний склад тимінових речовин. В. Р. Вільямі відводив основну увагу, ульмійовій і гуминовій кислотам, що він розглядав як фактор, що стабілізує, що консервує органічну речовину ґрунту.
Таким чином, в утворенні ґрунтового гумусу беруть участь сполуки типу лігніну і дубильні речовини, а також білкові речовини і вуглеводи. Безперечно беруть участь в утворенні структурних одиниць гумусу і продукти життєдіяльності мікроорганізмів. Останні, як відомо, не тільки розкладають різні органічні речовини, але й забезпечують різноманітні синтези.
Значно поповнилися відомості щодо загальної ролі гумусу й органічних речовин ґрунту. Продукти розкладу органічних залишків не тільки збагачують ґрунт доступними сполуками фосфору й азоту і поповнюють запаси СО2 у самих нижніх шарах атмосфери, але і проявляють різноманітну стимулююче дію на рослину (активування процесу надходження речовин у клітину внаслідок підвищення її прониклості, посилення дихання і т.п.
Ці спостереження дозволять пояснити високу ефективність органомінеральних добрив. Вони відкривають також можливість практичного! використання в якості «добрива» гумінових речовин, які вилучаються з торфу, бурого вугілля, сланців і інших джерел. Перші досвіди показали, що малі дози гумінових речовин дають позитивний ефект навіть при застосуванні на багатих органічними речовинами чорноземних ґрунтах.
Розкладання азотовмісних сполук рослинної тканиною і гумусу проходить через ряд етапів. У результаті цих процесів у ґрунті нагромаджується аміак у виді солей різних органічних і неорганічних кислот. Амоніфікація азотистих сполук здійснюється різними мікроорганізмами, які належать як до аеробних, так і анаеробних форм.
Аміак, що утворився, піддається окислюванню, яке також здійснюється східчасто. На першому етапі окислення утвориться азотиста кислота, окислювання якої приводить до утворення азотної кислоти. Кожний з етапів цього процесу, що носить назву нітрифікація, відбувається за допомогою специфічних бактерій. У різних ґрунтах нітрифікація відбувається з дуже розрізною інтенсивністю; на хід процесу впливають умови температури, аерація, вологість і багато інших факторів. Утворившись нітрати служать одним з основних джерел азотного живлення зелених рослин.
Поряд з цим у ґрунті йдуть і процеси денитрифікації, що здійснюються спеціальною групою анаеробних мікроорганізмів, так званих денітрифікаторів Діяльність останніх акцентується при нестачі кисню, яка поєднується з надлишком нерозкладених рослинних залишків. Під впливом цих бактерій відбувається відновлення нітратного іона й утворення молекулярного азоту.
Крім денітрифікації, джерелом втрат нітратів являється вимивання з ґрунту, від чого більше всього страждають рослини на легких супіщаних ґрунтах.
Істотно позначаються процеси мінералізації органічних залишків на стані інших важливих елементів (сірка, фосфор та ін.)
У природі розклад органічних сполук, що містять сірку супроводжується виділенням сірководню; лише деяка частина вихідної сірки виділяється у формі меркаптанів. Утворення сірководню проходить також і під впливом специфічних анаеробних,що здатні відновлювати окислені сполуки сірки (солі сірчаної, сірнистої і сірнувастої кислот) за рахунок енергії, яка добувається ними шляхом окислювання деяких органічних речовин
Розкладання органічних речовин впливає і на баланс фосфорнокислих сполук ґрунту. Фосфорна кислота, яка відщеплюється в результаті діяльності мікроорганізмів вступає в сполуку з різними катіонами ґрунту в результаті чого утворяться малодоступні вищій рослині фосфорнокислі солі кальцію, заліза, магнію і т д.
Одним з основних факторів підвищення доступності фосфорнокислих солей є вуглекислота, що виділяється у процесі дихання коренів рослин і ґрунтових мікроорганізмів
Поряд з цим у ґрунті протікають зворотні процеси, які приводять до зменшення розчинності фосфатів.
Як показали дослідження Ф. В. Чирикова, доступність фосфорнокислих солей ґрунту різна для різних груп рослин.
Велика роль належить ґрунтовим мікроорганізмам в утворенні в ґрунті розчинних солей К, Fe, Si, Mg та ін. Цей процес здійснюється шляхом впливу продуктів життєдіяльності мікробів на каоліни, польові шпати й інші мінерали.
3 Становлення кореневого живлення
Чим харчується рослина? Це питання було, природно, одним з перших, з яким зштовхнулися фітофізіологи. Тому визначення сучасного стану кореневого живлення вимагає висвітлення його історичного становлення. При цьому інтерес представляє не послідовність дат і фактів, а зміна ідей, що з'являлися при аналізі малопомітних фактів і відкриттів.
Джерела зародження науки про кореневе живлення рослин просліджуються в багатовіковому досвіді вирощування сільськогосподарських культур, практичної діяльності людей.
У середині XVI в. Б. Палісі (1510-1590) установив, що в золі спаленої соломи містяться солі, що рослини беруть із ґрунту. Він стверджував, що зола служить добривом, тому що ґрунту повертаються ті речовини, що були в неї узяті.
Голландський учений Я. ван Гельмонт (1579-1644) уперше висловив думку, що в рослині відбувається переробка речовин, які сприймаються, що рослини харчуються за допомогою коренів.
Наприкінці XVIII - початку XIX вв. у харчуванні рослин домінувала так називана гумусова теорія А. Теера (1752-1828). Автор, правильно оцінюючи позитивне значення органічної речовини ґрунту, заперечував значення мінеральних елементів для рослин.
У середині XIX в. німецький хімік Ю. Либих (1803-1873) уперше сформулював мінеральну теорію живлення рослин. Цьому сприяла розробка методів аналітичної хімії, що відкрили широкі можливості пізнання суті будови речовини. З ім'ям Лібіха пов'язаний початок теоретичних і практичних досліджень по мінеральному живленню. Учений вирішив проблеми землеробства чисто хімічним шляхом. Зробивши сотні аналізів органічної і зольної частини різних рослин, він визначив, що в рослинах містяться 10 основних елементів: вуглець, кисень, водень, сірка, залізо, кальцій, магній, азот, калій і фосфор. Перші три елементи надходять з повітря і води; вони складають основну масу рослини. Інші складові частини (мінеральні) дає земля.
Ю. Лібіх наполягав на тому, що господарювання традиційними методами безперестану і невблаганно виснажує ґрунт. Зміною рослин на полях можна тільки вповільнити виснаження, але не запобігти його.
Учений вважав за необхідне повернення в ґрунт мінеральних речовин, вбачаючи в цьому основний закон агрохімії. Він думав, що в ґрунт повинні бути в першу чергу повернуті ті речовини, запаси яких найбільш виснажені. На думку Лібіха, найбільша увага приділялася фосфорним добривам, що споживаються рослиною для утворення насіння. Пропозиція Лібіха про застосування фосфорних добрив у вигляді мелених кісток худоби, особливо під зернові культури, з'явилося поштовхом до розвитку суперфосфатної промисловості на базі викопних фосфатів.
Ю. Лібіх переконливо довів неспроможність основних положень гумусової теорії. Він сформулював уявлення про те, що органічна речовина, яка дає початок гумусу, виникає з засвоюваних рослиною мінеральних речовин ґрунту. Велика заслуга Лібіха полягає в тому, що він зумів зрозуміти закони природи і намагався правильно їх застосовувати. З властивою йому образністю він писав: „природа говорить з нами на її власній мові, вона завжди відповідає на питання, і ці питання - досвіди". Книга Лібіха „Хімія в додатку до землеробства і фізіології рослин" (1840) зробила визначений позитивний вплив на розвиток агрохімії.
Однак Ю. Лібіх прийшов до неправильного висновку про те, що азот надходить у ґрунт з атмосферними опадами у виді аміаку і цієї кількості азоту досить для харчування рослин. Французький агрохімік і фізіолог Ж. Буссенго (1802-1887) і німецький учений Г. Гельригель (1831-1895) спростували помилкове уявлення Лібіха про азотне харчування рослин. Використовуючи вегетаційні досвіди, автори знайшли здатність бобових рослин засвоювати молекулярний азот атмосфери. У дослідженнях з рослиною соняшника було доведено, що весь необхідний азот рослина добуває з ґрунту. Іншою помилкою Лібіха було твердження, що добриво потрібно вводити в ґрунт у нерозчиненому чи малорозчинному вигляді. Йому здавалося, що в протилежному випадку внесені солі вимиються першим же дощем. Про могутність утримуючої здатності ґрунту він тоді ще не здогадувався, а тим часом рослини засвоюють тільки розчинні сполуки.1
Російський ботанік М. С. Воронін (1838-1903) довів, що на коренях бобових з паренхімних тканин утворяться клубеньки, у клітках яких знаходяться клубенькові бактерії. Він уперше ретельно досліджував зрізи клубеньків на коренях люпину і знайшов у клітках тканини численних бактерій. П. А. Костичев (1845-1895) детально розробив питання про взаємодію між ґрунтом, рослинами й іншими її організмами. Дослідження Б. А. Келлера (1874-1945) з питань екології солончакових рослин у природі і шляхів їхніх пристосувань заслужено вважаються класичними. Крім галофітів, що накопичують солі у своїх органах, він установив тип галофітів, що виділяють надлишок солей. В. И. Вернадський (1863-1945) розробив основи біогеохімії. Він відзначав велике біогеохімічне значення ґрунту і вважав, що ґрунтові організми, будучи невід'ємною складовою частиною ґрунту, обумовлюють біохімічні процеси, що протікають у ній. В. И. Вернадський також заклав основи навчання про рідкі і розсіяні елементи, чи мікроелементах, у ґрунтах.
Значний інтерес до питань живлення рослин з'явився лише після того, як росіянин учений С. Н. Виноградський (1856-1953) встановив біологічну природу утворення в ґрунті нітратів, виділивши при цьому мікроорганізми-нітрифікатори. Йому належать відкриття анаеробної фіксації азоту і з'ясування ролі мікроорганізмів ґрунту в перетворенні гумусових речовин.
Важливими в теоретичному і практичному відношеннях для мінерального живлення рослин з'явилися роботи основоположника радянської школи агрохімії Д Н. Прянишникова (1865-1948). Він установив, що правильне використання мінеральних добрив є могутнім чинником регулювання фізіологічних процесів у рослин і формування врожаю. Він усебічно вивчав азотне живлення, довівши, що в слабкокислому середовищі нітрати поглинаються більш інтенсивно, ніж аміачні солі, а останні, навпаки, у нейтральному середовищі поглинаються більш енергійно. Дослідження Д Н. Прянишникова були покладені в основу заходів щодо хімізації сільського господарства.
Основні висновки про значення форм азоту в азотному харчуванні рослин, отримані Д Н. Прянишниковим і його учнями, зводяться до наступного.
1. При одночасній присутності в зовнішньому розчині нітратної й амонійної форм остання поглинається і споживається швидше.
2. Зовнішні і внутрішні оптимальні умови для харчування рослин аміаком і нітратами різні.
3. При порівнянні нітратів і амонію в оптимальних для кожного умовах вони фізіологічно рівноцінні.
4. Перевага нітратної чи амонійної форми залежить від реакції зовнішнього середовища, співвідношення катіонів, особливо Са2+ і К+, і змісту запасних вуглеводів у насінні і проростках.
Великий внесок у пізнання закономірностей кореневого живлення внесли роботи Д А. Сабініна. У його працях роль коренів у процесах росту рослин на всіх етапах онтогенезу освітлена в декількох напрямках.
1. Проникність плазми і поглинання кореневими системами води і розчинених у ній елементів мінерального живлення.
2. Роль коренів у водяному харчуванні.
3. Роль коренів у процесах мінерального харчування і їх метаболічна активність.
4. Кругообіг елементів мінерального живлення і їхній вплив на ріст і формоутворення рослин.
Метод збору й аналізу пасоки (кореневого соку), розроблений Д. А. Сабінін, є, по суті, модельною системою, що імітує хід синтетичних процесів у коренях цілих рослин. В основі цього лежить уявлення про те, що, збираючи пасоку, ми вивчаємо речовини, подавані коренем у надземні органи. Автор установив, що збір пасоки протягом 24-36 ч з моменту зрізання стебла дає стійкі для рослини цифри концентрації. Цей термін навіть для молодих проростків, що виділяють ще мало пасоки, є зовсім достатнім, щоб зібрати потрібне для аналізу кількість кореневого соку. Другою необхідною передумовою при використанні досліджень пасоки для судження про постачання надземних органів рослини тим чи іншій речовині зовнішнього розчину є знання долі даної речовини на шляху від поверхні кореневого волоска до судин кореня.1
Д А. Сабінін запропонував способи виділення й аналізу пасоки як критерій забезпеченості рослин живильними елементами. Він показав, що поглинені мінеральні елементи вступають у хімічну взаємодію з цитоплазмою клітин кореня і велика частина їхній передається в надземні органи в органічній формі. Учений зробив висновок про те, що поглинання і пересування речовин є активним процесом, зв'язаним з життєдіяльністю всієї рослини. Він довів, що першим етапом поглинання електролітів є їхня адсорбція кореневими системами. Активне поглинання іонів поділяється на два етапи: перший - адсорбція іонів на поверхні клітин, процес фізико-хімічний і оборотний; другий - пересування іонів усередину кліток, процес зв'язування адсорбованих іонів протоплазмою кліток і залучення їх у синтетичні реакції. Подальшими етапами є пересування по живих клітках і виділення в судини ксилеми. Таке представлення лежить в основі нашого сучасного розуміння процесів поглинання речовин кореневими системами.
Розглядаючи роль кореневих систем у водопостачанні рослин, Д. А. Сабінін вважав, що процеси поглинання рослинами води й елементів мінерального харчування різні і не знаходяться в прямої залежності між собою. Ці розуміння знайшли подальше підтвердження в багатьох роботах і тепер вважаються загальновизнаними.
Д А. Сабінін завжди розглядав корінь як частина цілої рослини. Адже вся діяльність кореня зв'язана з одним з основних життєвих процесів - подихом, а тому' і поглинальна здатність кореня залежить від пересування асиміляторів від листів до кореня. Діалектика дослідження тут полягала в тім, що, відрізаючи корінь від рослини і вивчаючи специфічність процесів, що відбуваються в ньому, удавалося краще осягати закономірності цілої рослини. Безпосереднє відображення це знайшло в експериментальних і теоретичних вишукуваннях проблеми круговороту елементів мінерального харчування і їхнього впливу на ріст і формоутворення рослин.
У круговороті елементів мінерального живлення ведучу роль Д А. Сабінін додавав кореневій системі, вважаючи, що процес поглинання речовин найтіснішим образом зв'язаний з життєдіяльністю всієї рослини. Разом з тим він підкреслював необхідність обліку процесів розвитку рослин. Різна здатність до зв'язування елементів мінерального живлення в органів і частин рослин у залежності від їхнього віку робить, на думку Д. А. Сабініна, що вирішує вплив на їхній розподіл. На цьому ґрунтувалися його узагальнюючі представлення про реутилізацію і круговорот елементів мінерального харчування і різних градієнтів змісту: базипетальному для рухливих елементів - азоту, фосфору, сірки і калію й акропетальному для менш рухливих - кальцію, заліза, бора, марганцю і цинку.
У з'ясуванні питань про пересування і розподіл мінеральних речовин він надавав великого значення досвідам з кільцюванням різних ділянок стебла і начертав загальну картину пересування елементів мінерального харчування по великому і малому колах, виділивши три групи елементів: а) типу азоту, що рухаються по великому колу; б) типу калію, що пересуваються по малих колах; в) типу кальцію, не приймаючої участі в круговому русі мінеральних речовин.
Уся широта поглядів Д. А. Сабініна на проблеми кореневого живлення, оригінальність його підходів і новизна висловлюваних положень розкриваються повною мірою в його наукових працях
Дотримуючи поглядів класиків російського ґрунтознавства й агрономії, що розглядали ґрунт як самостійне природно-історичне тіло і надавали значення біологічним процесам, що протікають у ґрунті, і поглядів росіян фітофізіологів, ми можемо визнати цілком закономірним виділення: самостійного напрямку - кореневого живлення рослин.
4 Кореневе живлення в житті рослин
4.1 Значення кореневого живлення для життя рослин
Кореневе живлення рослин важливий фізіологічний процес в житті рослин. Важливість цього процесу визначається двома обставинами. З одного боку, цей процес в фізіології рослин є теоретичною основою раціонального використання мінеральних речовин - ефективного засобу керування продуктивністю сільськогосподарських рослин; з іншого боку - кореневе живлення розкриває багатобічний взаємозв'язок між рослиною і середовищем за допомогою харчування мінеральними елементами. Від подальших успіхів наукових досліджень кореневого харчування рослин залежить рішення деяких назрілих питань продовольчої проблеми.
Сутність кореневого живлення складається в поглинанні і включенні в метаболізм мінеральних елементів у результаті обміну речовин між рослиною і навколишнім середовищем.
Виникший інтерес до кореневого живлення рослин пов'язаний з його теоретичними успіхами біохімічного, онтогенетичного й екологічного напрямків. Біохімічний напрямок розглядає функціональне значення макро- і мікроелементів для рослинного організму, виявляє шляхи біосинтезу органічних сполук з елементів мінерального живлення, визначає роль мінеральних речовин як регуляторів стану колоїдів клітки. Онтогенетичний напрямок досліджує можливі шляхи керування ростом і розвитком рослин за допомогою елементів мінерального харчування. Екологічний напрямок виявляє залежність внутрішніх процесів рослинного організму від наявності хімічних елементів у зовнішнім середовищі. Найбільш актуальними питаннями в науці про кореневе живлення є наступні:
розробка біохімічної теорії харчування рослин для більш ефективного використання довгостроково діючих мінеральних добрив і використання елементів мінерального харчування як фактора стійкості рослин;
вивчення можливості регулювання співвідношення між вегетативними й органами, що запасають, за допомогою мінерального харчування;
з'ясування поглинаючих і метаболичних функцій кореневих систем під впливом фізіологічно активних речовин;
вивчення ролі елементів мінерального харчування у формуванні енергетичних і окисних систем рослин;
розкриття механізму біологічної фіксації азоту атмосфери і використання його вищими рослинами;
вивчення питань проникнення іонів через клітинні мембрани і їх компартментация в клітках, транспорту іонів у тканини й органи;
з'ясування ролі мінерального харчування при культурі тканин, зв'язаного з розробкою сучасних біотехнологій в одержанні високопродуктивних форм рослин з меристем;
вивчення питань кореневого харчування при керуванні вегетативним ростом і генеративним розвитком рослин.
З'ясування цих питань наблизить нас до керування продуктивністю рослин на більш високій теоретичній основі мінерального живлення; розгляд іонного транспорту як системи процесів, що складають основу матеріального й енергетичного обмінів у рослині; обґрунтування процесів засвоєння рослиною мінеральних речовин як активних фізіологічних явищ.
4.2 Функції кореневої системиПитання про функції кореневої системи представляє вузол проблем, зв'язаних з живленням рослин, і тому необхідно познайомитися з основами виділення і синтезу речовин у коренях, транспорту іонів у клітку, тканини й органи.
Роль кореня в житті всієї рослини багатогранна. У першу чергу, корінь - це спеціалізований орган поглинання води і мінеральних елементів із ґрунту. Друга сторона діяльності кореневої системи - часткова чи повна переробка поглинених іонів, їхнє відновлення, включення в різні органічні сполуки і транспортування в наземні органи для синтезу складних метаболітів і фізіологічно активних речовин. Ця сторона синтезуючої функції коренів зв'язана з процесом поглинання. Третя функція - виділення в навколишнє середовище речовин, різних по хімічній природі і біологічному значенні. Фізіологічні функції кореня знаходяться в тісному зв'язку з його анатомічною будовою.
У роботах Д Н. Прянишникова вивчення питання про порівняльну цінність різних неорганічних сполук азоту, як джерел азотного живлення, послужило частиною чудових досліджень по перетворенню сполук азоту в кореневій системі. Спочатку перед дослідниками виникло приватне запитання про походження і роль аспарагіну. Відомо, що ця речовина нагромаджується при проростанні насіння у темряві в дуже значних кількостях, що його азот складає біля половини всього азоту проростків. Широким поширенням користалося уявлення про аспарагін, як продукт розпаду білків, що виникає при мобілізації білкових запасів насіння. Експериментально Д Н. Прянишников (1945) довів, що аспарагін не може бути безпосереднім продуктом розпаду білків при проростанні насіння, а виникає в результаті вторинного синтезу. Це було зроблено шляхом проведення досвідів по вивченню здатності рослин засвоювати азот аміачних солей при різних умовах виростання. Проростки гороху, ячменя і люпину жовтого тим краще здатні утворювати аспарагін за рахунок амонійних солей, які даються ними ззовні, чим краще вони забезпечені вуглеводами. У темряві проростки в процесі подиху розтрачують свої вуглеводні запаси і перетворюються в організми, позбавлений можливості використовувати аміак для синтезу аспарагіну.1
Д Н. Прянишников ясно уявляв, що рослини мають у своєму розпорядженні можливість знешкодження аміаку шляхом його проміжного зв'язування з органічними молекулами, що виникають у метаболізмі. У результаті вивчення дикарбонових амінокислот і амідів у рослин у період інтенсивного білкового обміну він знайшов, що ці з'єднання грають важливу проміжну і разом з тим захисну роль у системі перетворень, що супроводжуються звільненням і використанням аміаку. Подальші дослідження розкрили біохімічну картину первинної асиміляції коренями іонів амонію і показали центральну роль у цьому процесі циклу ди- і трикарбонових кислот. Джерела азоту, що поглинається коренями, зазнають амінування й амідування.
Аналіз пасоки дозволив Д А. Сабініну в 1949 р. обґрунтувати концепцію про синтетичну роль коренів. Основні положення цієї концепції наступні:
корінь здатний не тільки поглинати мінеральні елементи, але і перетворювати їх ( чичастково цілком) і подавати в наземні органи в зміненому виді;
синтезуюча діяльність кореня розвертається на підставі асиміляторів, що надходять з фотосинтезованого органа;
корінь впливає на надземні органи не тільки за рахунок забезпечення їхньою водою і мінеральними елементами, але і за рахунок продуктів специфічних реакцій обміну речовин, що відбуваються в коренях.
Практично цілком підтвердилося положення про синтез у коренях фізіологічно активних речовин гормональної природи неауксинового типу. Серед речовин, які потрібні для росту і знаходяться в пасоці, у першу чергу необхідно назвати цитокінини. Вони синтезуються в коренях і пересуваються з пасокою в надземні органи
Центральними реакціями первинного включення аміаку в коренях більшості рослин є, на думку багатьох авторів, амінування (a-кетоглутарової кислоти й амідування глутаминової кислоти. Оскільки в пасоці часто переважає аланін, зроблений висновок про те, що перед виносом NН2-сполук з коренів у погони відбувається часткове переамінування з глутамінової на піровиноградну кислоту. Ця суміш частково використовується в коренях для синтезу власних білків, частково ж виділяється в трахеїди ксилеми і захоплюється з висхідним струмом пасоки в надземні органи, досягаючи крапок росту і листів
У деяких рослин як перші продукти асиміляції NН3 при нормальних умовах утворяться такі багаті NН2-групами сполуки, як цитрулін, алантоінова кислота, аллнтоін і деякі інші. Ці речовини також виділяються в пасоку і відносяться до органічних транспортних форм азоту. Запас їх МН,-груп використовується для переамінування з іншими акцепторами, що приводить до утворення амінокислот.
У корені рослина легка використовує аміни для синтезу алкалоїдів (атропін беладони, нікотин різних видів тютюну).
Ізольовані корені представляють дуже зручний об'єкт для вивчення синтезу ряду речовин, особливо речовин вторинного походження. Так, А. А. Прокоф'єв (1944) показав, що в коренях ізольованої культури тау-сагизу формувалася млечная система, у якій здійснювався синтез каучуку без зв'язку з надземними органами.
Здатність коренів синтезувати ряд з'єднань, у тому числі вітамінів, детально вивчалася в досвідах А. М. Смирнова (1970). Для цих цілей використовувалися корені багатьох рослин кленового походження, що до проведення досвідів тривалий час знаходилися в ізольованій культурі, позбавивши, отже, цілком впливу на них надземних органів. У коренях, що виросли з кінчиків бічних коренів довжиною 25-30 мм за семиденний період, у темряві завжди утворювалася аскорбінова кислота. Порівняльний зміст аскорбінової кислоти в ізольованих коренях приведено нижче (мг % на сиру масу): томати - 6,65; морква - 8,01; люцерна - 22,20; вика - 35,80; горох - 36,74.
Приведені дані показують, що найбільше інтенсивно аскорбінова кислота синтезувалася в коренях бобових, особливо вики і гороху.
Корені цілих рослин (проростків) при вирощуванні їх у темряві зберегли здатність до синтезу аскорбінової кислоти. При цьому корені бобових синтезували її в більшій кількості, чим корені небобових. На підставі цих даних можна допустити, що в коренях проростків аскорбінова кислота накопичується не в результаті відтоку її з листів, а утвориться за рахунок процесів біосинтезу, що протікають безпосередньо в тканинах коренів.
Відомо, що синтез амінокислот локалізований у визначених ділянках кореня. Так, найбільша кількість амінокислот виявлена в частині кореня кукурудзи, що відстоїть на 1-5 див від його кінчика в зоні живих кореневих волосків. Середина і підстава кореня містять мало амінокислот і виконують роль їхнього провідника в надземну частину рослини. Аналіз кореневих розгалужень (корені першого і другого порядків) показав таке ж розташування зони синтезу, тобто вони повторюють будівлю і функції основного кореня.
Закономірності розподілу амінокислот у різних частинах кореня не є загальними для всіх рослин, що підтверджують дані по змісту їх у різних частинах томатів. Головний корінь томатів містить менша кількість всіх амінокислот, чим бічні корені першого і другого порядків. Можливо, це свідчить про різне розташування більш-менш активних частин у корені в різних рослин. Дослідження основних коренів і коренів першого порядку кукурудзи показали, що в більшості ділянок основних коренів міститься менше амінокислот, чим у тих же ділянках коренів першого порядку, що може свідчити про більш активну і продуктивну роботу молодих коренів. Слід зазначити, що найбільша кількість амінокислот, знайдена в односантиметрових кінчиках коренів кукурудзи, виявлялося протягом усієї вегетації.
Перетворення амінокислот у пасоці й у коренях при вегетації показує, що біосинтез амінокислот у коренях грає більш значну роль у початкові фази вегетації. У коренях рослин у міру їхнього розвитку змінюються як спрямованість біосинтезу, так і перетворення амінокислот. З віком змінюється не тільки кількісне, але і якісний зміст амінокислот. .
Установлено дзеркальний характер зміни змісту амінокислот у коренях і пасоці в онтогенезі. Так, у кукурудзі до 36 днів вирощування зміст амінокислот у пасоці зменшувалося з одночасним підвищенням їхньої кількості в коренях. Потім спостерігалися протилежні зміни. При вивченні амінокислотного складу пасоки виноградної лози на початку руху соку виявлені валин, пролін, аланін, глутамінова кислота, серин, гліцин, глутамін, аспарагін, лізин. У період руху соку в пасоці з'являються фенілаланін, лейцин, треонін, аспарагінова й аміномасляна кислоти.
Корені рослин здійснюють асиміляцію азоту переважно в зоні кори. У досвідах С. Ф. Ізмайлова (1986) показано, що основною тихорєцькою зоною, де відбуваються перетворення амінокислот і їхнє нагромадження, є паренхімні клітини кори. При асиміляції азоту в тканинах коренів переважно використовується сахароза невеликого по величині метаболічного фонду. Цей фонд має тісний метаболічний зв'язок з екзогенними цукрами, унаслідок чого при асиміляції азоту досягається швидке використання вуглецю цукрів, що надійшов у клітку.
Кількісний склад амінокислот перетерплює істотні зміни в процесі росту клітки кореня. Велика кількість амінокислот у зонах розтягання і диференціації свідчить про зростання потреби в них для білкового синтезу, здатність до якого збільшується в міру росту клітки.
Зміна амінокислотного складу коренів і подачі з пасокою амінокислот у процесі вегетації рослин є відображенням нормального функціонування коренів, їхній синтезуючий діяльності протягом онтогенезу.
Корені виконують у цілому рослині як би роль “залози", що виробляє за рахунок асиміляторів, що надходять з листів, і азоту ґрунту багато вторинних азотистих сполук, у тому числі речовини регуляторного характеру. Синтезовані в коренях сполуки виносяться потім у надземні органи. Кругообіг речовин у рослині є ланкою кореневого живлення і тому чітко контролюється потребою рослини і тими джерелами живлення, якимип воно володіє. Цей кругообіг тісно пов'язаний з поглинаючою і видільною діяльністю коренів і служить в основному для розподілу вироблених ними NН2 - сполук.
Однієї з функцій кореневої системи є виділення в навколишнє середовище речовин, різних за хімічною природою і біологічному значенню. Експериментально встановлено, що через кореневу систему виділяються майже всі типи водорозчинних органічних сполук. У складі кореневих виділень виявлено багато різноманітних амінокислот і органічних кислот. Неодмінними компонентами кореневих виділень є цукри. Кількість і склад кореневих виділень визначаються видовими і сортовими особливостями рослин. Наприклад, кореневі виділення в бобових рослин набагато багатше амінокислотами, чим у злакових; яблуня через корені виділяє фенольні речовини, а овес - речовини типу лактонів.
Є відомості про те, що корені здатні виділяти мінеральні речовини. Яскравим прикладом можуть служити бобові й олійні культури, корені яких виділяють фосфорну кислоту й інші мінеральні елементи.
Отже, кількість виділюваних у ґрунт речовин може істотно перевершувати рівень їхнього змісту в самому корені. Цей феномен означає, що екскреторні процеси визначаються життєдіяльністю всього організму, а корінь виконує функцію органа виділення. Процес виділення речовин постійно відбувається в звичайних умовах середовища і, мабуть, є нормальною функцією рослинного організму. Зараз визнається існування постійного круговороту живильних речовин по рослині, під яким варто розуміти пересування елементів ґрунтового харчування спочатку з коренів у надземні органи, а потім знову в корені, відкіля частина речовин може мігрувати назад у ґрунт.1
Одним з найбільш переконливих підтверджень інтенсивних виділень є відзначені факти негативного балансу процесів харчування на останніх етапах онтогенезу рослин. Поглинені коренями зольні елементи наприкінці вегетації в значних кількостях повертаються в ґрунт (38 % ДО, 22 % Са, 10%Мо). Виділення фосфорної кислоти коренями люпину, гірчиці, ярового рапсу може скласти близько 14-34%, усієї фосфорної кислоти, що поглинається рослиною.
Швидкості виділення і поглинання К, Nа, Са, Со настільки великі, що рослини за період вегетації здатні поглинути і виділити в десятки разів більше цих елементів, чим містити їх у собі в якийсь даний період вегетації.
Виділення метаболітів коренями властиве зоні кореневих волосків, і кореневі виділення, як правило, мають кисле середовище.
Виникає питання: яка екологічна доцільність викиду асимільованого вуглецю і раніше поглинених елементів мінерального живлення
1. Безсумнівно, ці виділення, з одного боку, можуть бути пристосувальною реакцією на умови зовнішнього середовища, що змінюються. Відзначено, що висихання ґрунту до початку зів'янення рослин і наступний її поливши сприяють посиленому виділенню амінокислот і відновлених з'єднань з рослин. Результати досвіду Н. В. Мєшкова (1971) показали значне збільшення кореневих виділень в умовах зміни живильних розчинів у порівнянні з незмінюваними. Якщо ж зміст вуглецю в органічних речовинах кореневих виділень виразити у відсотках від загального вуглецю в рослинах, тоді цей показник для гороху буде дорівнює 4,04 % у незмінюваних розчинах і 10,96 % - у змінюваних; для кукурудзи відповідно 1,0 і 2,26 %.
З іншого боку, виділення позаклітинних гідролаз у ризосферу, очевидно, свідчить про пристосування рослинних форм у процесі еволюції до використання деяких елементів мінерального складу.
2. Виділені кореневою системою продукти життєдіяльності накопичуються в ризосфері і служать живильним субстратом для ризосферної і ґрунтової мікрофлори, тобто корені забезпечують її азотом і вуглецем у легко доступній формі. Наявність легко доступної органічної речовини сприяє розвитку клубенькових бактерій. У вигляді кореневих виділень безпосередньо на процеси азотфіксації витрачається від 25 до 37 % вуглецю, фотосинтезованого рослиною. .
3. Частина кореневих виділень реутилізується тим же чи суміжно розташованою рослиною ценозу. Прямий обмін метаболітами коренів сусідніх рослин між собою відіграє ведучу роль у взаєминах рослин у фітоценозах.
Як відомо, при багаторічному, беззмінному вирощуванні тієї чи іншої культури відбувається збідніння якісного складу мікрофлори. Рослинні виділення можуть придушувати деяких представників ґрунтової мікрофлори.
Негативний вплив токсичних кореневих виділень деякою мірою пов'язаний з наявністю в ґрунті органічної речовини. Чим нижче вміст органічної речовини в ґрунті, тим швидше настає стомлюваність ґрунту.
Результати визначення активності ферментів ризосфери показали, що ферментативна активність ґрунту знаходиться в тісному зв'язку з діяльністю кореневих систем. Про здатність тонких закінчень коренів виділяти в навколишнє середовище активну протеазу указував В.Д.Купревич (1954). Виявляється також визначена залежність між чисельністю мікроорганізмів у ґрунті й активністю протеаз.
Кореневі виділення здатні змінювати водяний режим у ґрунті і рослині, порушувати інтенсивність транспірації і подиху, засвоєння вуглеводів.
4.3 Поглинання елементів мінерального живлення рослин
Споживання мінеральних елементів - саморегульований процес. В основі його лежить здатність рослин підтримувати збалансованість рівнозначних потоків іонів мінеральних солей, що забезпечує необхідний для життєдіяльності рівень нагромадження мінеральних елементів. Будь-яке відхилення від цього рівня викликає ряд процесів, спрямованих на відновлення втраченої рівноваги, тобто на включення гомеостатичних механізмів. При цьому можливі не тільки повернення до вихідного рівня нагромадження мінеральних елементів у клітках, але і перехід (відповідно до їхнього змісту в зовнішнім середовищі) на новий рівень, що лежить у межах фізіологічних норм відхилень.
Під іонним гомеостазом клітини варто розуміти систему, що забезпечує підтримку і розподіл внутрішньоклітинних концентрацій і активностей іонів і води. Підтримка іонного гомеостазу в клітках рослин може досягатися різними шляхами. Один зі шляхів регуляції іонного гомеостазу рослин полягає в тому, що поглинання одного іона може відбуватися за рахунок виділення іншого. Особливо у великих розмірах таке заміщення може відбуватися в галофітів. У них повнота заміщення живильних іонів баластовими (Nа +,Cl-) досягає 70 %.
Іншим шляхом регуляції іонного гомеостазу рослин є посилення видільної діяльності кореневої системи, сольових залоз. А. М.Смирнов (1970), вивчаючи ріст і метаболізм ізольованих коренів у стерильній культурі, установив, що в процесі росту в ізольованій культурі корені виділяють у середовище крім органічних сполук катіони й аніони.
Третій шлях підтримки іонного гомеостазу - це перерозподіл змісту іонів в органах рослин. Надлишок іонів солей може приділятися в старі листи, затримуватися в коренях.
Оскільки для нормального функціонування кліток рослин важлива не тільки концентрація тих чи інших іонів солей, але і їхнє співвідношення, то в клітці відбуваються процеси, що забезпечують необхідне співвідношення різно зарядних іонів. Тут набирає сили закон сталості відносини суми катіонів до суми аніонів:
Рослина поглинає катіони в еквівалентно більшій кількості, чим аніони, тому одержуване частка небагато більше 1.
Прикладом цього можуть бути численні дослідження і кількісний аналіз живильних речовин, що витрачаються деревом на створення деревини, листів і плодів. Підсумувавши дослідні дані, отримані в різних країнах, і показники виносу, установлені для різних умов оброблення плодових культур, можна одержати наступні середні значення виносу чистих живильних елементів (у кг/га): азот - 76; фосфор - 33; калій - 110; кальцій - 70.
Якщо взяти за основу вищенаведені середні значення виносу живильних речовин, то співвідношення N: Р:К:Са буде відповідати 1:0,4:1,4:0,9. Навіть при однобічному внесенні мінеральних речовин його величина майже не міняється, тому що інтенсивне поглинання калію потім буде вирівняно, наприклад, незначним поглинанням магнію і кальцію, що є антагоністами, чи сильне поглинання N0^ буде урівноважено не менш інтенсивним поглинанням катіонів унаслідок їх синергізму. Посилений синтез органічних кислот є одним з діючих способів зв'язування катіонів і нейтралізації з'єднань основного характеру. Велику роль в утриманні кислотно-лужної рівноваги в клітках грають органічні полікатіони і поліаніони (амінокислоти, нуклеїнові кислоти, фосфоліпіди. полісахариди).
Основним, найчастіше єдиним, джерелом мінеральних речовин для рослин служить ґрунт. Генетично контрольовані потреби рослини визначають лінію його поводження: при дефіциті елементів харчування підсилюється розчинююча діяльність чи коренів з їх допомогою освоюється новий обсяг ґрунту, при надлишку - затримується надходження живильних речовин.1
Вивчення питання транспорту елементів є одним з основних у мінеральному харчуванні. Це визначено наступним: по-перше, сутність харчування рослин складається в поглинанні і включенні в метаболізм мінеральних елементів у результаті обміну між організмом і середовищем.
По-друге, з'ясування питань, зв'язаних із транспортом, наближає нас до керування продуктивністю сільськогосподарських рослин на більш високій теоретичній основі. Знаючи умови поглинання і пересування того чи іншого елемента, можна кількісно змінити зміст його в тканинах.
По-третє, вивчення транспорту елементів сполучено з з'ясуванням властивостей і функцій клітинної оболонки, мембранних утворень, зв'язку між клітками і тканинами.
В остаточному підсумку іонний транспорт накладає відбиток на всі основні групи явищ: перетворення речовин, енергії, передачу інформації і форми.
Тривалий час серед фізіологів рослин був розповсюджений погляд, відповідно до якого елементи мінерального харчування надходять у рослини разом з водою на основі осмотичних і дифузійних закономірностей. Вважалося, що речовини поглинаються рослиною в тих же кількостях і співвідношеннях, у яких вони знаходяться в ґрунтовому розчині. Експериментальна перевірка і логічний аналіз цих представлень показали, що вони зовсім неспроможні. Було встановлено, що процеси поглинання води й елементів мінерального харчування в широких межах незалежний друг від друга. Вода необхідна як розчинник речовин, у розчиненому стані окремі елементи пересуваються по рослині, але поглинання речовин рослиною відбувається вибірково, що може цілком змінювати співвідношення поглинених речовин у порівнянні з тими, котрі маються в зовнішньому розчині. Це дуже важливе принципове питання фізіології кореневого харчування рослин.
Експерименти цілком підтверджують теоретичний висновок про незалежність процесів поглинання солей і води кореневими системами. Д А. Сабінін (1955) приводить три ряди фактів, що є підставою для твердження про незалежності цих процесів.
По-перше, не існує прямого зв'язку між кількістю транспірованої рослинами води і кількістю солей, поглинених з розчину, що оточує кореневі системи. Наводяться дані чеського ботаніка С. Прата по зміні електропровідності розчину в зв'язку з поглинанням солей Мкена, що знаходилися в різних умовах транспірації. Установлено, що навіть при слабкій транспірації (11 відносних одиниць) відбувалося інтенсивне поглинання солей (50 відносних одиниць). Відзначено, що при більшій транспірації спостерігається деяке посилення поглинання солей, але воно є дуже невеликим, що ледь виходить за межі погрішності методу обліку, що застосовувався автором, поглинання солей.
Другий ряд факторів, що свідчать про відсутність зв'язку між поглинанням води і розчинених речовин із середовища, що оточує корені, це дані про одночасний рух іонів солей і води через кореневі системи в протилежних напрямках. При розгляді видільної функції коренів відзначалося, що з кореневих систем серед інших речовин виділяються в зовнішнє середовище різні іони. Наприклад, при рн < 6,0 з коренів злаків, вирощених у водяній культурі, виділяється кальцій. При визначених значеннях рН на світлі в умовах інтенсивної транспірації відбувається виділення сульфат- і фосфат-іонів.
Третій ряд факторів, що обґрунтовують представлення про незалежність поглинання іонів коренями від поглинання води, складають результати досвідів по засвоєнню кореневими системами іонів із ґрунту.
Завдяки роботам ґрунтознавців і фізіологів стало відомо, що живильні речовини з ґрунту в корені надходять переважно у формі іонів, чи присутніх у розчині, чи адсорбованих частками. Поглинання іонів здійснює головним чином молода (зростаюча) частина коренів. Мається кілька шляхів, що забезпечують сталість контактування коренів з елементами харчування. По-перше, це досягається завдяки активному пошуку необхідних іонів самою рослиною: збільшенню довжини коренів і освоєнню нової товщі ґрунту. Так здійснюється перехоплення елементів харчування. По-друге, іони надходять у корені з масовим струмом, що виникає в ґрунті в результаті транспірації води рослинами. По-третє, іони пересуваються з ґрунту убік коренів дифузно по градієнті концентрації.
Пайова участь кожного з перерахованих шляхів доставки іонів до коренів у залежності від умов може істотно мінятися. Так, доставка багатьох елементів-біофілів до коренів здійснюється переважно масовим потоком. Однак це можливо тільки при їхньому значному змісті в ґрунтовому розчині. Якщо ґрунтовий розчин бідний елементами харчування, то відбувається обмін іонами між клітками епідермісу і частками ґрунту. Обмін може чи відбуватися безпосередньо між поверхнями чи клітки частки, чи в результаті переходу іонів у ґрунтовий розчин.
Відповідно до сучасних представлень, на першому етапі поглинання елементів мінерального харчування значну роль грає їхня адсорбція на поверхні кліток і тканин. Перша протікає за рахунок електричних сил адсорбуючої поверхні, друга - за рахунок взаємодії з зарядами амфотерних з'єднань протоплазми. Торкаючись питання про значення адсорбції в процесах харчування рослин, слід зазначити роботи Д А. Сабініна (1940), И. И. Колосова (1962). Автори вказували, що розгляд процесу надходження речовин у клітку повинне починатися з ефекту взаємодії оболонок рослинних кліток з іонами зовнішнього середовища, і досить чітко сформулювали уявлення про клітинну стінку як про іонообмінну фазу.
Значна інформація про роль оболонки в первинному поглинанні іонів була отримана в роботах, присвячених питанню про локалізацію так називаного вільного простору. Д Б. Вахмістрів (1969) довів, що вільний простір локалізований у клітинній оболонці і не поширюється на цитоплазму. Цей висновок прийшов на зміну раніше пануючим представленням, згідно яким основним місцем адсорбції іонів є протоплазма.
Ряд досвідів, виконаних на ізольованих клітинних оболонках позбавлених слідів цитоплазми, показав їх високу адсорбційну ємність.
Таким чином, сорбційні властивості клітинної оболонки повинні значно впливати на процес поглинання іонів рослинною кліткою. Припускають, зокрема, що в результаті адсорбції при дуже низьких іонних концентраціях у навколишнім середовищі відбувається значне концентрування речовин на фазовій границі клітка/зовнішній розчин. Показано, що клітинна стінка, як перший бар'єр на шляху проникнення елементів мінерального харчування, є слабкокисла катіонообмінна мембрана, матриця якої утворена целюлозою і несе визначену кількість карбоксильних груп, що зв'язують катіони. Подібне притягання до клітинної стінки позитивно заряджених часток збільшує місцеву концентрацію розчинних речовин, що приводить до підтримки в клітинній стінці більш високого осмотичного тиску, чим у навколишньому розчині.
Ключову роль у здійсненні контролю за рухом іонів із клітки в клітку відіграє плазмалема. Для елементів мінерального живлення вона служить просто бар'єром, що обмежує їхній рух по градієнту концентрацій. Однак для деяких іонів роль плазмалеми більш специфічна. У цих випадках у мембранах включаються механізми, що полегшують рух іонів через мембрану по градієнту їхніх концентрацій. Це відбувається тоді, коли клітки активно накопичують які-небудь іони, концентрація яких повинна підтримуватися на низькому рівні. При цьому активному транспорті іонів через мембрану витрачається енергія, запасена у формі АТФ.
Отже, по своєму відношенню до енергетичних процесів транспортні системи рослин поділяють на пасивні й активні механізми. Пасивні механізми приводять лише до прискорення вирівнювання концентрації речовин у зовнішнім середовищі і у внутрішньому обсязі кліток. Вважається, що рушійною силою такого процесу є концентраційний градієнт, і тому передбачається, що перенос, що є власне кажучи полегшеною дифузією, відбувається без додаткової витрати енергії.
Висновки
Ґрунт є поживним субстратом рослин: у ньому міститься головний запас потенційної біогенної енергії у вигляді коренів рослин, біомаси мікроорганізмів і гумусу. Без надходження з ґрунту таких елементів як фосфор і калій, було б неможливе створення первинної рослинної продукції.
Якісний склад золи залежить від вмісту мінеральних речовин у ґрунті й умов зовнішнього середовища. Як правило, чим багатший ґрунт і чим сухіше клімат, тим вище вміст золи в рослинах. Будь - який хімічний елемент, наявний у даному місцеперебуванні, може бути виявлений і в рослині. Багато біометалів (Fе, Мп, Си, Zn, Мо і Co) у рослинах взаємодіють з макромолекулами. Такі макромолекули прийнята називати біолігандами. До природних лігандів варто віднести білки, нуклеїнові кислоти і вітаміни. Координаційні сполуки характеризуються насамперед координаційним числом, тобто числом атомів, що складають найближче оточення центрального атома - біометалу (атома комплексоутворювача). Метал і його найближче оточення складають внутрішню сферу координаційної сполуки.
Велику роль в утворенні самого ґрунту і нагромадженні у верхніх її обріях мінеральних елементів виконують у процесі своєї життєдіяльності і самі рослини. Поглинаючи своїми коренями мінеральні речовини, рослини піднімають їх у верхні шари ґрунту і тим самим збагачують останні.
Умови мінерального живлення в значній мірі залежать від типу ґрунту. Тому при виявленні потреб рослини в мінеральному добриві необхідно виходити як з видових особливостей організму, так і зі специфічних особливостей даного типу ґрунту.
Сутність кореневого живлення складається в поглинанні і включенні в метаболізм мінеральних елементів у результаті обміну речовин між рослиною і навколишнім середовищем.
Роль кореня в житті всієї рослини багатогранна. У першу чергу, корінь - це спеціалізований орган поглинання води і мінеральних елементів із ґрунту. Друга сторона діяльності кореневої системи - часткова чи повна переробка поглинених іонів, їхнє відновлення, включення в різні органічні сполуки і транспортування в наземні органи для синтезу складних метаболітів і фізіологічно активних речовин. Ця сторона синтезуючої функції коренів зв'язана з процесом поглинання. Третя функція - виділення в навколишнє середовище речовин, різних за хімічною природою і біологічному значенні. Фізіологічні функції кореня знаходяться в тісному зв'язку з його анатомічною будовою.
Споживання мінеральних елементів - саморегульований процес. В основі його лежить здатність рослин підтримувати збалансованість рівнозначних потоків іонів мінеральних солей, що забезпечує необхідний для життєдіяльності рівень нагромадження мінеральних елементів.
Література
1. Алексеев А.М., Гусев Н.А. Влияние корневого питания на водный режим. – М., 1957. – 220 с.
2. Вахмистров Д.Б. Питание растений. – М.: Знание, 1979. – 64 с.
3. Колосов И.И. Поглотительная деятельность корневых систем растений. – М.: АН СССР, 1962. – 388с.
4. Максимов Н.А. Краткий курс физиологии растений, 1958
5. Мусиенко Н.Н., Терневский А.И. Корневое питание растений: Учебное пособие. – К.: Высшая школа, 1989. – 203 с.
6. Нобел П. Физиология растительной клетки. – Л.: Изд-во ленингр. Ун-та, 1983. – 232 с.
7. Петербургский А.В. Корневое питание растений. – М., 1964. 340 с.
8. Потапов Н.Г. Минеральное питание // Физиология сельскохозяйственных растений. – М., 1967. – 320 с.
9. Рубин Б.А. Курс физиологии растений. – М.: Высшая школа, 1961. –583 с.
1 Максимов Н.А. Краткий курс физиологии растений, 1958
1 Максимов Н.А. Краткий курс физиологии растений, 1958
1 Мусиенко Н.Н., Терневский А.И. Корневое питание растений: Учебное пособие. – К.: Высшая школа, 1989. – 203 с.
1 Мусиенко Н.Н., Терневский А.И. Корневое питание растений: Учебное пособие. – К.: Высшая школа, 1989. – 203 с.
1 Мусиенко Н.Н., Терневский А.И. Корневое питание растений: Учебное пособие. – К.: Высшая школа, 1989. – 203 с.
1 Мусиенко Н.Н., Терневский А.И. Корневое питание растений: Учебное пособие. – К.: Высшая школа, 1989. – 203 с.
1 Мусиенко Н.Н., Терневский А.И. Корневое питание растений: Учебное пособие. – К.: Высшая школа, 1989. – 203 с.
1 Петербургский А.В. Корневое питание растений. – М., 1964. 340 с.
1 Петербургский А.В. Корневое питание растений. – М., 1964. 340 с.
1 Петербургский А.В. Корневое питание растений. – М., 1964. 340 с.
1 Петербургский А.В. Корневое питание растений. – М., 1964. 340 с.
Похожие работы
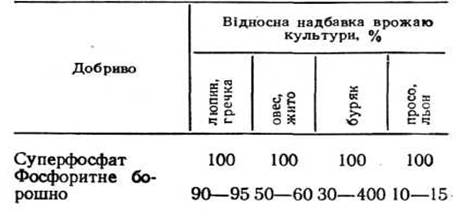

... а з другого — досягти найвищої окупності кожної одиниці добрива. В основі системи лежить створення раціонального живлення рослин з урахуванням біологічних особливостей їх і умов вирощування. Оскільки культурні рослини, як правило, вирощують у сівозміні, то й систему застосування добрив будують стосовно до конкретної сівозміни. При цьому враховують такі основні моменти: особливості живлення рослин, ...
... локалізація всіх трьох елементів живлення. Річну дозу добрив під окремі культури можна вносити в різні терміни і різними способами. Терміни і прийоми внесення добрив повинні забезпечувати якнайкращі умови живлення рослин протягом всієї вегетації і отримання найбільшої окупності живильних речовин урожаєм. Розрізняють три способи внесення добрив: допосівне (або основне), припосівне (в ряди, кубла, ...
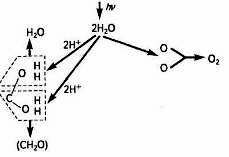
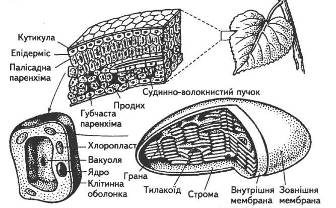
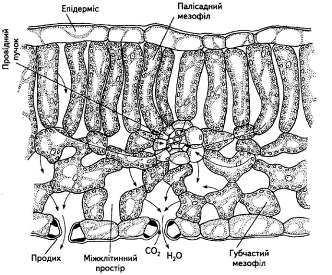
... однакова, тому його пул постійно оновлюється в хлоропласті. Нагадаємо, що в 1960 p. P. Вудворд і М. Штрель штучно синтезували молекули хлорофілу. Важливе значення для синтезу хлорофілів має мінеральне живлення рослин. Для цього передусім необхідна достатня кількість заліза — каталізатора синтезу хлорофілів. Залізо необхідне для синтезу амінолевулінової кислоти та протопорфіринів. Для синтезу ...
... при нормі 1–2 кг на 1 га. Позакореневе підживлення можна поєднувати з обприскуванням рослин отрутохімікатами для боротьби з хворобами та шкідниками. 2. Біологічні особливості найважливіших сільськогосподарських культур 2.1 Технічні культури Рослини, що дають сировину для легкої промисловості, називаються технічними. За призначенням урожаю їх поділяють на прядивні, олійні, цукро- і ...
0 комментариев