Навигация
Фізіологічні основи живлення рослин и застосування добрив
110187
знаков
11
таблиц
7
изображений
Зміст
Вступ
Розділ 1. Живлення рослин: суть процесу та його особливості 1.1 Коренева система як орган поглинання та обміну речовин
1.2 Функції кореневої системи
Розділ 2. Класифікація добрив 2.1 Мінеральні добрива
Вступ
Розділ 1. Живлення рослин: суть процесу та його особливості
1.1 Коренева система як орган поглинання та обміну речовин
1.2 Функції кореневої системи
1.3 Поглинання елементів мінерального живлення рослин
1.4 Фізіологічні основи використання мінеральних добрив
Розділ 2. Класифікація добрив
2.1 Мінеральні добрива
2.2 Органічні добрива
2.3 Бактеріальні добрива
Розділ 3. Агротехнічні вимоги до внесення органічних і мінеральних добрив
Розділ 4. Екологічний стан ґрунтів
Експериментальна частина
Методика дослідження
Результати дослідження
Висновки
Список використаних джерел
Вступ
У житті рослин важливу роль відіграє живлення рослин. Нові технології вирощування рослин базуються на використанні нових видів добрив. Оптимальне живлення рослинних організмів у сполученні з раціональним підвищенням ефективності застосування добрив і зменшенням забруднення навколишнього середовища продуктами хімізації дозволить підвищити врожай і поліпшити якість сільськогосподарської продукції.
Умови мінерального живлення в значній мірі залежать від типу ґрунту. Тому при виявленні потреб рослини в мінеральному добриві необхідно виходити як з видових особливостей організму, так і зі специфічних особливостей даного типу ґрунту.
Мета роботи: з’ясувати значення живлення рослин та залежність агрохімічних показників родючості від застосування добрив на прикладі Шосткінського району Сумської області.
Предмет дослідження мінеральне живлення рослин.
Об’єкт дослідження впливу використання добрив на врожайність окремих культур.
Завдання даної роботи:
1) З`ясувати суть та процеси мінерального живлення рослин;
2) Охарактеризувати основні класи добрив;
3) Проаналізувати агротехнічні вимоги до внесення добрив;
4) Встановити залежність агрохімічних показників родючості ґрунту від використаних та внесених елементів мінерального живлення на прикладі Шосткінського району Сумської області.
5) Встановити залежність врожайності сільськогосподарських культур від внесення добрив на прикладі Шосткінського району Сумської області.
Робоча гіпотеза: внесення в грунт елементів мінерального живлення сприяє покращенню агрохімічних показників ґрунтів та зростанню врожайності сільськогосподарських культур.
Розділ 1. Живлення рослин: суть процесу та його особливості 1.1 Коренева система як орган поглинання та обміну речовин
У кореневому живленні рослин, як і в фотосинтезі, проявляється одна з найяскравіших властивостей рослинного організму — автотрофність, тобто здатність будувати своє тіло з неорганічних речовин. Крім того, саме живлення рослин забезпечує постійний кругообіг речовин і перенесення енергії, тісно поєднуючи мінеральний та живий світ.
Коренева система, будучи спеціалізованим органом поглинання води, забезпечує також і надходження до рослини мінеральних речовин. Функція кореня не обмежується лише поглинанням і транспортуванням речовин у надземні органи, адже коренева система є також органом активного і спеціалізованого обміну речовин. Корінь забезпечує також виділення в навколишнє середовище різних за природою та біологічним значенням речовин. Фізіологічні функції кореня тісно пов'язані з його анатомічною будовою [12].
Сформована коренева система — досить складний орган з добре диференційованою внутрішньою структурою.
Диференціювання клітин кореня розпочинається в зоні поділу клітин меристеми. В зоні росту розтягуванням ці процеси прискорюються — з'являється ризодерма (епіблема), перші провідні елементи прото- й метафлоеми, трициклу.
Зони поділу та росту розтягуванням кореня — найактивніші ділянки поглинання води і мінеральних речовин. На зовнішній поверхні клітин ризодерми утворюються кореневі волоски, де найактивніше відбуваються процеси метаболізму і куди постійно надходять поживні речовини. Зона поглинання, де зосереджена основна маса кореневих волосків — найактивніша всмоктувальна частин;) кореня.
В міру відмирання ризодерми з кореневими волосками на по верхні кореня з первинної кори формується нова покривна тканина — екзодерма. Клітини первинної кори активно і пасивно транспортують речовини до центрального циліндра кореня, а також виконують функцію синтезу і накопичення різних запасних речовин. На шляху речовин, що надходять із фунтового розчину, є ряд бар'єрів.
Перший із них — це ризодерма, а другий — внутрішній шар клітин первинної кори — ендодерма, бічні стінки оболонок яких мають особливі потовщення — пояски Каспарі, просякнуті лігніном і суберином, а тому непроникні для йонів. Однак пояски Каспарі є не в усіх клітин ендодерми, серед них є так звані пропускні клітини, які не лігніфіковані. Вони локалізовані в місцях утворення бічних коренів і крізь них йони легко транспортуються.
Зона ендодерми — фізіологічний бар'єр, де протопласт "контролює" потік йонів. Функція ендодерми цим не вичерпується, адже вона також затримує та регулює надходження речовин із центрального циліндра в периферійні тканини.
Ще одним бар'єром на шляху до центрального циліндра кореня є перицикл, що складається з активних меристематичних клітин. Перицикл — зовнішній шар осьового центрального циліндра кореня. В ньому накопичуються різні речовини, в тому числі фітогормони, що стимулюють утворення бічних коренів.
Завдяки таким фізіологічним бар'єрам розчин, який потрапляє в центральний циліндр кореня, значно трансформується, а тому відрізняється від ґрунтового розчину. В корені синтезуються "напівфабрикати", які з часом легко залучаються до біосинтетичних процесів уже в стеблі чи листках [8].
Центральний циліндр кореня характеризується радіальним розміщенням первинних провідних тканин — флоеми та ксилеми. Отже, особливості будови кореня багато в чому визначають його функції. Як відомо, листки та корені рослин полярно відрізняються за напругою окисно-відновного потенціалу. Листки мають низьку напругу, корені — навпаки. Утримання даного потенціалу на рівні, що відповідає нормальній життєдіяльності рослин, забезпечується відповідним обміном продуктами їх власної синтетичної функції.
Ще в 1949 р. Д. А. Сабінін обґрунтував концепцію про синтетичну функцію кореня: корінь не лише поглинає мінеральні елементи, а й перетворює їх у процесі транспортування в надземні органи. Синтетична діяльність кореня ґрунтується на базі асимілятів, які надходять від донорів органів фотосинтезу.
Кругообіг речовин у рослині — важлива ланка кореневого живлення. Він зумовлений специфічними вимогами рослини до поживних елементів та джерелами живлення, які для неї доступні. Цей кругообіг тісно пов'язаний як із поглинальною, так і з видільною функцією кореня. Доведено, що через кореневу систему виділяються майже всі типи водорозчинних сполук. Логічно виникає запитання щодо екологічної доцільності таких виділень. Вони можуть бути пристосувальною реакцією на зміну умов середовища, поживними елементами для ризосферної та ґрунтової мікрофлори, частина їх реутилізується тим самим або поряд розміщеним рослинним організмом ценозу. Кореневі виділення зумовлюють явище алелопатії (взаємного впливу) рослин, що входять до складу фітоценозу. Активними алелопатичними агентами ґрунту є тритерпени: еритрородіол і сератіол. Зазначимо, що такі токсичні виділення рослин, як халкони та дигідрохалкони (флоридзин), у ґрунті швидко руйнуються, втрачаючи свою активність. Кількість і склад таких виділень (алкалоїди, глікозиди, сапоніни, кумарини, терпени, флавоноїди тощо) залежать від виду рослин [12].
Розглядається навіть концепція алелохімікатів, тобто продуктів, що утворюються в разі взаємодії рослин і ґрунту, та можуть бути використані як форма природних гербіцидів. Створення такої системи природних гербіцидів може стати однією з біотехнологічних моделей, яка дасть змогу уникнути використання в рослинництві синтетичних гербіцидів із підвищеною токсичністю.
Доведено, що у різних рослин спостерігається специфічність кореневих виділень, із якими пов'язують так звану ґрунтовтому.
Це явище є типовою ознакою антропогенних змін в екосистемі ґрунту. Монокультура як екологічний фактор порушує природні процеси в масштабі екосистеми ґрунту. Втомлений ґрунт характеризується надмірним розвитком шкідливих для рослин мікроорганізмів. Встановлено, що 15...45 % виділених із таких ґрунтів мікроорганізмів продукують фітотоксини. В таких ґрунтах надмірно розвиваються бактеріофаги, що знищують бульбочкові бактерії, знижуючи продуктивність азотфіксації.
Ґрунтовтома — одна з найбільших проблем світового землеробства, яка перебуває під контролем продовольчої комісії Організації Об'єднаних Націй (ФАО ООН). За даними цієї комісії внаслідок ґрунтовтоми щороку втрачається 25 % загальних біологічних втрат світового врожаю. Таке явище як ґрунтовтома у природних ценозах не спостерігається.
Токсичність речовин, що виділяються коренем, найчутливіше проявляється в зоні ризосфери — шарі ґрунту (2—3 мм), що безпосередньо оточує кореневу систему рослин. Кореневі виділення, відмерлі кореневі волоски сприяють розвиткові в ризосфері різноманітних мікроорганізмів. Деякі з них забезпечують рослини азотом, синтезують біологічно активні речовини, які стимулюють ріст рослин, мінералізують органічні сполуки. Інші, навпаки, пригнічують рослини, виділяючи певні токсини. Взаємодія кореневих систем і мікрофлори проявляється у формі ризосферного ефекту, величина якого визначається відношенням кількості мікроорганізмів у ризосфері до вмісту їх у ґрунті за її межами.
Кореневі виділення здатні змінити водний режим ґрунту й рослини, інтенсивність дихання, транспірації, засвоєння вуглеводів тощо.
1.2 Функції кореневої системи
Питання про функції кореневої системи представляє цілий ряд проблем, пов'язаних з живленням рослин, і тому необхідно познайомитися з основами виділення і синтезу речовин у коренях, транспортом іонів у клітину, тканини й органи.
Роль кореня в житті всієї рослини багатогранна. У першу чергу, корінь - це спеціалізований орган поглинання води і мінеральних елементів з ґрунту. Друга сторона діяльності кореневої системи - часткова чи повна переробка поглинених іонів, їх відновлення, включення в різні органічні сполуки і транспортування в наземні органи для синтезу складних метаболітів і фізіологічно активних речовин. Ця сторона синтезуючої функції коренів пов'язана з процесом поглинання. Третя функція - виділення в навколишнє середовище речовин, різних за хімічною природою і біологічним значенням [9].
У роботах Д Н. Прянишникова вивчення питання про порівняльну цінність різних неорганічних сполук азоту, як джерел азотного живлення, послужило частиною цікавих досліджень з перетворення сполук азоту в кореневій системі. Спочатку зацікавило запитання про походження і роль аспарагіну. Відомо, що ця речовина нагромаджується при проростанні насіння у темряві в дуже значних кількостях, що азот аспарагіну складає біля половини всього азоту проростків. Значного поширення набуло уявлення про аспарагін, як продукт розпаду білків, що виникає при мобілізації білкових запасів насіння. Експериментально Д. Н. Прянишников (1945) [9] довів, що аспарагін не може бути безпосереднім продуктом розпаду білків при проростанні насіння, а виникає в результаті вторинного синтезу. Це було зроблено шляхом проведення дослідів по вивченню здатності рослин засвоювати азот аміачних солей при різних умовах використання. Проростки гороху, ячменя і люпину жовтого тим краще здатні утворювати аспарагін за рахунок амонійних солей, які надходять ззовні, чим краще вони забезпечені вуглеводами. У темряві проростки в процесі дихання витрачають свої вуглеводні запаси і перетворюються в організм, позбавлений можливості використовувати аміак для синтезу аспарагіну.
Д Н. Прянишников уявляв, що рослини мають можливість знешкодження аміаку шляхом його проміжного зв'язування з органічними молекулами, що виникають у процесі метаболізму. У результаті вивчення дикарбонових амінокислот і амідів у рослин у період інтенсивного білкового обміну він знайшов, що ці з'єднання грають важливу проміжну і разом з тим захисну роль у системі перетворень, що супроводжуються вивільненням і використанням аміаку. Подальші дослідження розкрили біохімічну картину первинної асиміляції коренями іонів амонію і показали провідну роль у цьому процесі циклу ди- і трикарбонових кислот. Азот, що поглинається коренями, зазнає амінування й амідування.
Д А. Сабінін в 1949 р. обґрунтувати концепцію про синтетичну роль коренів. Основні положення цієї концепції наступні:
· корінь здатний не тільки поглинати мінеральні елементи, але і перетворювати їх (частково або цілком) і подавати в наземні органи в зміненому вигляді;
· корінь впливає на надземні органи не тільки за рахунок забезпечення їх водою і мінеральними елементами, але і за рахунок продуктів специфічних реакцій обміну речовин, що відбуваються в коренях.
Практично підтвердилося положення про синтез у коренях фізіологічно активних речовин гормональної природи неауксинового типу. Серед речовин, які потрібні для росту, у першу чергу необхідно назвати цитокінини. Вони синтезуються в коренях і пересуваються в надземні органи.
Основними реакціями первинного включення аміаку в коренях більшості рослин є, на думку багатьох авторів, амінування a-кетоглутарової кислоти й амідування глутаминової кислоти. Оскільки в патоці часто переважає аланін, був зроблений висновок про те, що перед надходженням NН2-сполук з коренів у погони відбувається часткове переамінування з глутамінової в піровиноградну кислоту. Ця суміш частково використовується в коренях для синтезу білків, частково ж виділяється в трахеїди ксилеми і переноситься з висхідним током патоки в надземні органи, досягаючи точок росту і листків [9].
У деяких рослин як перші продукти асиміляції NН3 при нормальних умовах утворюються такі багаті NН2-групами сполуки, як цитрулін, алантоінова кислота, алантоін і деякі інші. Ці речовини також виділяються в патоку і належать до органічних транспортних форм азоту. Їх NН3-групи використовуються для переамінування з іншими акцепторами, що приводить до утворення амінокислот.
Здатність коренів синтезувати ряд сполук, у тому числі вітамінів, детально вивчалася в досвідах А. М. Смирнова (1970). Приведені дослідження показали, що найбільше інтенсивно аскорбінова кислота синтезується в коренях бобових, особливо вики і гороху.
Корені рослин (проростки) при вирощуванні їх у темряві зберегли здатність до синтезу аскорбінової кислоти. При цьому в коренях бобових синтезувалося її більше, ніж в коренях інших рослин. На підставі цих даних можна припустити, що в коренях проростків аскорбінова кислота накопичується не в результаті відтоку її з листів, а утворюється за рахунок процесів біосинтезу, що протікають безпосередньо в тканинах коренів [9].
Відомо, що синтез амінокислот локалізований у певних ділянках кореня. Перетворення амінокислот у патоці й у коренях при вегетації показує, що біосинтез амінокислот у коренях відіграє більш значну роль у початковій фази вегетації. У коренях рослин у міру їх розвитку змінюються як спрямованість біосинтезу, так і перетворення амінокислот. З віком змінюється не тільки кількісний, але і якісний вміст амінокислот [13].
Корені рослин здійснюють асиміляцію азоту переважно в зоні кори.
Кількісний склад амінокислот змінюється в процесі росту клітин кореня. Велика кількість амінокислот у зонах розтягнення і диференціації свідчить про зростання потреби амінокислот для білкового синтезу, здатність до якого збільшується в міру росту клітини.
Зміна амінокислотного складу коренів і подачі з патокою амінокислот у процесі вегетації рослин є відображенням нормального функціонування коренів, їх синтезуючої діяльності протягом онтогенезу.
Корені в рослині як би виконують роль "залози", що виробляє за рахунок асиміляторів, які надходять з листків, і азоту з ґрунту багато вторинних азотистих сполук, у тому числі речовини регуляторного характеру. Синтезовані в коренях сполуки надходять у надземні органи. Кругообіг речовин у рослині є ланкою кореневого живлення і тому чітко контролюється потребою рослини. Цей кругообіг тісно пов'язаний з поглинаючою і видільною діяльністю коренів і служить в основному для розподілу вироблених NН2 - сполук.
Однієї з функцій кореневої системи є виділення в навколишнє середовище речовин, різних за хімічною природою і біологічним значенням. Експериментально встановлено, що через кореневу систему виділяються майже всі типи водорозчинних органічних сполук. У складі кореневих виділень виявлено багато різноманітних амінокислот і органічних кислот. Неодмінними компонентами кореневих виділень є цукри. Кількість і склад кореневих виділень визначаються видовими і сортовими особливостями рослин. Наприклад, кореневі виділення в бобових рослин значно багатші на амінокислоти, ніж злакові; яблуня через корені виділяє фенольні речовини, а овес - речовини типу лактонів.
Є відомості про те, що корені здатні виділяти мінеральні речовини. Яскравим прикладом можуть служити бобові й олійні культури, корені яких виділяють фосфорну кислоту й інші мінеральні елементи [9].
Отже, кількість виділених у ґрунт речовин може істотно перевищувати рівень їх вмісту в самому корені. Цей феномен означає, що екскреторні процеси визначаються життєдіяльністю всього організму, а корінь виконує функцію органа виділення. Процес виділення речовин відбувається постійно і, мабуть, є нормальною функцією рослинного організму. Зараз визнається існування постійного круговороту поживних речовин по рослині, під яким варто розуміти пересування елементів ґрунтового живлення з коренів у надземні органи, а потім знову в корені, звідки частина речовин може мігрувати назад у ґрунт.
Одним з найбільш переконливих підтверджень інтенсивних виділень є відзначені факти негативного балансу процесів живлення на останніх етапах онтогенезу рослин. Поглинені коренями зольні елементи наприкінці вегетації в значних кількостях повертаються в ґрунт (22 % Са, 10 % Мо). Виділення фосфорної кислоти коренями люпину, гірчиці, ярового рапсу може скласти близько 14-34%, усієї фосфорної кислоти, що поглинається рослиною.
Швидкості виділення і поглинання К, Nа, Са, Со настільки великі, що рослини за період вегетації здатні поглинути і виділити в десятки разів більше цих елементів, ніж містили їх у собі в період вегетації.
Виділення метаболітів коренями властиве зоні кореневих волосків, і кореневі виділення, як правило, мають кисле середовище.
Виникає питання: яка екологічна доцільність викиду асимільованого вуглецю і раніше поглинених елементів мінерального живлення.
1. Безсумнівно, ці виділення, з одного боку, можуть бути пристосувальною реакцією до змін у зовнішньому середовищі. Відзначено, що висихання ґрунту до початку зів'янення рослин і наступного її поливу сприяють посиленому виділенню амінокислот і відновлених сполук з рослин.
З іншого боку, виділення позаклітинних гідролаз у ризосферу, очевидно, свідчить про пристосування рослинних форм у процесі еволюції до використання деяких елементів мінерального складу.
2. Виділені кореневою системою продукти життєдіяльності накопичуються в ризосфері і служать поживним субстратом для ризосферної і ґрунтової мікрофлори, тобто корені забезпечують її азотом і вуглецем у легко доступній формі. Наявність легко доступної органічної речовини сприяє розвитку клубенькових бактерій. У вигляді кореневих виділень безпосередньо на процеси азотфіксації витрачається від 25 до 37 % вуглецю, фотосинтезованого рослиною.
3. Частина кореневих виділень реутилізується. Прямий обмін метаболітами коренів сусідніх рослин між собою відіграє ведучу роль у взаєминах рослин у фітоценозах.
Як відомо, при багаторічному, беззмінному вирощуванні тієї чи іншої культури відбувається збідніння якісного складу мікрофлори. Рослинні виділення можуть пригнічувати існування деяких представників ґрунтової мікрофлори.
Негативний вплив токсичних кореневих виділень деякою мірою пов'язаний з наявністю в ґрунті органічної речовини. Чим нижчий вміст органічної речовини в ґрунті, тим швидше настає стомлюваність ґрунту.
Результати визначення активності ферментів ризосфери показали, що ферментативна активність ґрунту знаходиться в тісному зв'язку з діяльністю кореневих систем. Про здатність тонких закінчень коренів виділяти в навколишнє середовище активну протеазу вказував В.Д.Купревич (1954). Виявляється також певна залежність між чисельністю мікроорганізмів у ґрунті й активністю протеаз.
Кореневі виділення здатні змінювати водяний режим у ґрунті і рослині, порушувати інтенсивність транспірації, дихання і засвоєння вуглеводів [12].
1.3 Поглинання елементів мінерального живлення рослинПоглинання мінеральних елементів - саморегульований процес. В основі його лежить здатність рослин підтримувати збалансованість рівнозначних потоків іонів мінеральних солей, що забезпечує необхідний для життєдіяльності рівень нагромадження мінеральних елементів. Будь-яке відхилення від цього рівня викликає ряд процесів, спрямованих на відновлення втраченої рівноваги, тобто на включення гомеостатичних механізмів. При цьому можливі не тільки повернення до вихідного рівня нагромадження мінеральних елементів у клітках, але і перехід (відповідно до їхнього змісту в зовнішнім середовищі) на новий рівень, що лежить у межах фізіологічних норм відхилень.
Під іонним гомеостазом клітини варто розуміти систему, що забезпечує підтримку і розподіл внутрішньоклітинних концентрацій і активностей іонів і води. Підтримка іонного гомеостазу в клітинах рослин може досягатися різними шляхами. Один зі шляхів регуляції іонного гомеостазу рослин полягає в тому, що поглинання одного іона може відбуватися за рахунок виділення іншого. Особливо у великих розмірах таке заміщення може відбуватися в галофітів. У них повнота заміщення поживних іонів баластовими (Nа +,Cl-) досягає 70 %.
Іншим шляхом регуляції іонного гомеостазу рослин є посилення видільної діяльності кореневої системи, сольових залоз. А. М.Смирнов (1970), вивчаючи ріст і метаболізм ізольованих коренів у стерильній культурі, установив, що в процесі росту в ізольованій культурі корені виділяють у середовище крім органічних сполук катіони й аніони.
Третій шлях підтримки іонного гомеостазу - це перерозподіл вмісту іонів в органах рослин. Надлишок іонів солей може виділятися в старі листки, затримується в коренях [8].
Оскільки для нормального функціонування клітин рослин важлива не тільки концентрація тих чи інших іонів солей, але і їх співвідношення, то в клітині відбуваються процеси, що забезпечують необхідне співвідношення різнозарядних іонів. Тут набирає сили закон сталості відношення суми катіонів до суми аніонів.
Рослина поглинає катіони в еквівалентно більшій кількості, ніж аніони, тому одержана частка небагато більша 1.
Прикладом цього можуть бути численні дослідження і кількісний аналіз поживних речовин, що витрачаються деревом на створення деревини, листків і плодів. Підсумувавши показники виносу, установлені для різних умов оброблення плодових культур, можна одержати наступні середні значення виносу чистих поживних елементів (у кг/га): азот - 76; фосфор - 33; калій - 110; кальцій - 70.
Якщо взяти за основу вищенаведені середні значення виносу поживних речовин, то співвідношення N: Р : К : Са буде відповідати 1:0,4:1,4:0,9. Навіть при однобічному внесенні мінеральних речовин його величина майже не змінюється, тому що інтенсивне поглинання калію потім буде вирівняно, наприклад, незначним поглинанням магнію і кальцію, що є антагоністами, чи сильне поглинання NО3- буде урівноважено не менш інтенсивним поглинанням катіонів унаслідок їх синергізму. Посилений синтез органічних кислот є одним з діючих способів зв'язування катіонів і нейтралізації сполук основного характеру. Велику роль в утриманні кислотно-лужної рівноваги в клітинах відіграють органічні полікатіони і поліаніони (амінокислоти, нуклеїнові кислоти, фосфоліпіди. полісахариди).
Основним, і найчастіше єдиним, джерелом мінеральних речовин для рослин служить ґрунт.
Вивчення питання транспорту елементів є одним з основних у мінеральному живленні. Це визначено наступним: по-перше, сутність живлення рослин полягає в поглинанні і включенні в метаболізм мінеральних елементів у результаті обміну між організмом і середовищем.
По-друге, з'ясування питань, пов'язаних із транспортом, наближає нас до керування продуктивністю сільськогосподарських рослин на більш високій теоретичній основі. Знаючи умови поглинання і пересування того чи іншого елемента, можна кількісно змінити вміст його в тканинах [12].
По-третє, вивчення транспорту елементів сполучено із з'ясуванням властивостей і функцій клітинної оболонки, мембранних утворень, зв'язку між клітками і тканинами.
В остаточному підсумку іонний транспорт накладає відбиток на всі основні групи явищ: перетворення речовин, енергії, передачу інформації.
Тривалий час серед фізіологів рослин була розповсюджена думка, відповідно до якої елементи мінерального живлення надходять у рослини разом з водою на основі осмотичних і дифузійних закономірностей. Вважалося, що речовини поглинаються рослиною в тих же кількостях і співвідношеннях, у яких вони знаходяться в ґрунтовому розчині. Було встановлено, що процеси поглинання води й елементів мінерального живлення в широких межах незалежні одне від одного. Вода необхідна як розчинник речовин, у розчиненому стані окремі елементи пересуваються по рослині, але поглинання речовин рослиною відбувається вибірково, що може цілком змінювати співвідношення поглинених речовин у порівнянні з тими, які є в зовнішньому розчині. Це дуже важливе принципове питання фізіології кореневого живлення рослин.
Експерименти цілком підтверджують теоретичний висновок про незалежність процесів поглинання солей і води кореневими системами. Д А. Сабінін (1955) [8] приводить три ряди фактів, що є підставою для твердження про незалежності цих процесів.
По-перше, не існує прямого зв'язку між кількістю транспірованої рослинами води і кількістю солей, поглинених з розчину, що оточує кореневі системи. Установлено, що навіть при слабкій транспірації (11 відносних одиниць) відбувалося інтенсивне поглинання солей (50 відносних одиниць). Відзначено, що при більшій транспірації спостерігається деяке посилення поглинання солей, але воно є дуже невеликим, що ледь виходить за межі похибки методу обліку поглинання солей.
Другий ряд факторів, що свідчать про відсутність зв'язку між поглинанням води і розчинених речовин із середовища, що оточує корені, це дані про одночасний рух іонів солей і води через кореневі системи в протилежних напрямках. При розгляді видільної функції коренів відзначалося, що з кореневих систем серед інших речовин виділяються в зовнішнє середовище різні іони. Наприклад, при рН < 6,0 з коренів злаків, вирощених у водяній культурі, виділяється кальцій. При визначених значеннях рН на світлі в умовах інтенсивної транспірації відбувається виділення сульфат- і фосфат-іонів.
Третій ряд факторів, що обґрунтовують представлення про незалежність поглинання іонів коренями від поглинання води, складають результати досвідів по засвоєнню кореневими системами іонів із ґрунту.
Завдяки роботам ґрунтознавців і фізіологів стало відомо, що живильні речовини з ґрунту в корені надходять переважно у формі іонів, чи присутніх у розчині, чи адсорбованих частками. Поглинання іонів здійснює головним чином молода (зростаюча) частина коренів. Існує кілька шляхів, що забезпечують сталість контактування коренів з елементами харчування. По-перше, це досягається завдяки активному пошуку необхідних іонів самою рослиною: збільшенню довжини коренів і освоєнню нової товщі ґрунту. Так здійснюється перехоплення елементів харчування. По-друге, іони надходять у корені з масовим струмом, що виникає в ґрунті в результаті транспірації води рослинами. По-третє, іони пересуваються з ґрунту убік коренів дифузно по градієнті концентрації [8].
Пайова участь кожного з перерахованих шляхів доставки іонів до коренів у залежності від умов може істотно мінятися. Так, доставка багатьох елементів-біофілів до коренів здійснюється переважно масовим потоком. Однак це можливо тільки при їхньому значному змісті в ґрунтовому розчині. Якщо ґрунтовий розчин бідний елементами харчування, то відбувається обмін іонами між клітками епідермісу і частками ґрунту. Обмін може чи відбуватися безпосередньо між поверхнями чи клітки частки, чи в результаті переходу іонів у ґрунтовий розчин.
Відповідно до сучасних представлень, на першому етапі поглинання елементів мінерального харчування значну роль грає їхня адсорбція на поверхні кліток і тканин. Перша протікає за рахунок електричних сил адсорбуючої поверхні, друга - за рахунок взаємодії з зарядами амфотерних з'єднань протоплазми. Торкаючись питання про значення адсорбції в процесах харчування рослин, слід зазначити роботи Д А. Сабініна (1940), И. И. Колосова (1962) [12]. Автори вказували, що розгляд процесу надходження речовин у клітину повинен починатися з ефекту взаємодії оболонок рослинних клітин з іонами зовнішнього середовища, і досить чітко сформулювали уявлення про клітинну стінку як про іонообмінну фазу.
Значна інформація про роль оболонки в первинному поглинанні іонів була отримана в роботах, присвячених питанню локалізації так званого вільного простору. Д Б. Вахмістров (1969) довів, що вільний простір локалізований у клітинній оболонці і не поширюється на цитоплазму. Цей висновок прийшов на зміну раніше пануючим уявленням, згідно яким основним місцем адсорбції іонів була протоплазма.
Ряд досвідів, виконаних на ізольованих клітинних оболонках позбавлених слідів цитоплазми, показав їх високу адсорбційну ємність.
Таким чином, сорбційні властивості клітинної оболонки повинні значно впливати на процес поглинання іонів рослинною клітиною. Припускають, зокрема, що в результаті адсорбції при дуже низьких іонних концентраціях відбувається значне концентрування речовин на фазовій границі клітина/зовнішній розчин. Показано, що клітинна стінка, як перший бар'єр на шляху проникнення елементів мінерального живлення, є слабкокисла катіонообмінна мембрана, матриця якої утворена целюлозою і несе певну кількість карбоксильних груп, що зв'язують катіони. Притягування до клітинної стінки позитивно заряджених часток збільшує концентрацію розчинних речовин, що приводить до підтримки в клітинній стінці більш високого осмотичного тиску, ніж у розчині.
Ключову роль у здійсненні контролю за рухом іонів із клітини в клітину відіграє плазмалема. Для елементів мінерального живлення вона служить просто бар'єром, що обмежує їх рух по градієнту концентрацій. Однак для деяких іонів роль плазмалеми більш специфічна. У цих випадках у мембранах включаються механізми, що полегшують рух іонів через мембрану по градієнту їх концентрацій. Це відбувається тоді, коли клітини активно накопичують які-небудь іони, концентрація яких повинна підтримуватися на низькому рівні. При такому активному транспорті іонів через мембрану витрачається енергія, накопичена у формі АТФ.
Отже, по своєму відношенню до енергетичних процесів транспортні системи рослин поділяють на пасивні й активні механізми. Пасивні механізми приводять лише до прискорення вирівнювання концентрації речовин у зовнішнім середовищі і у внутрішньому об`ємі клітин. Вважається, що рушійною силою такого процесу є концентраційний градієнт, і тому передбачається, що перенос, що є власне кажучи полегшеною дифузією, відбувається без додаткової витрати енергії.
1.4 Фізіологічні основи використання мінеральних добривВикористання мінеральних добрив — основний засіб підвищення врожайності сільськогосподарських культур. За рахунок азоту, фосфору, калію та інших дефіцитних у землеробстві елементів, які містяться у відповідних типах мінеральних добрив, культурні рослини краще використовують енергію Сонця і ґрунтово-кліматичні фактори, що дає змогу одержувати додаткові врожаї. Нестачу цих елементів неможливо замінити ніякими іншими агротехнічними заходами.
Ефективність добрив залежить від потреб рослин у поживних елементах і здатності ґрунту задовольняти ці потреби. Таку взаємозалежність образно виразив Д. М. Прянишников [9] у вигляді трикутника, де у вершинах кутів розташовуються рослина — грунт — добрива, причому рослина розміщена у верхньому куті. Цим підкреслюється, що рослина і її врожайність — основа всіх проблем економічних і біологічних наук у практиці сільського господарства. Існують три загальноприйняті правила діагностики живлення.
• Впродовж вегетаційного періоду контролюють ступінь забезпечення посіву майбутнього врожаю основними елементами живлення. Основою діагностики живлення є хімічний склад рослин протягом вегетації. Одночасно з хімічним аналізом рослин слід враховувати етапи їхнього росту і розвитку.
• Визначають кілька поживних елементів, причому не менше трьох основних — азот, фосфор, калій.
• Зіставлення даних хімічної діагностики з урахуванням особливостей росту рослин та кліматичними умовами й агротехнікою.
Рослинна діагностика не замінює ґрунтових аналізів, а слугує для глибшого розуміння забезпечення рослин поживними речовинами в конкретних умовах їх вирощування.
Це дає змогу уточнити необхідний і ефективніший склад виду добрив. Облік винесення основних поживних елементів (азоту, фосфору, калію) сільськогосподарськими культурами — один з найважливіших показників, необхідних для встановлення раціональних доз добрив.
Показники винесення поживних речовин польовими культурами відображають зональний характер, що зумовлюється сортовими особливостями культур, зміною умов вирощування та зоною поширення окремих культур. Кількість доступних для рослин поживних речовин у ґрунті залежить також і від кількості внесених добрив. Тому в інтенсивних технологіях землеробства саме добривам належить основна функція створення в ґрунті оптимальних гармонійних співвідношень між елементами живлення [9].
Для визначення доз треба враховувати неврівноважене співвідношення між поживними речовинами ґрунту і вносити поправки в дози застосовуваних добрив, щоб привести кількісне співвідношення N : Р: К у ґрунті у відповідність до потреб кожної культури. У практиці землеробства найчастіше доводиться мати справу з азотними, фосфорними та калійними добривами.
Азотні добрива характеризуються високою ефективністю в усіх ґрунтово-кліматичних зонах. За підрахунком відомого агрохіміка О. В. Петербурзького [9], 1 кг азоту міндобрив за високої агротехніки забезпечує приріст урожаю зерна 20 кг/га. Залежно від форми азоту азотні мінеральні добрива поділяють на чотири групи:
· аміачні (азот міститься у формі аміаку — амоній сульфат, амоній хлорид, аміачна вода);
· нітратні (аніон нітратної кислоти — натрієва і кальцієва селітра);
· аміачно-нітратні (аміачна і вапняно-аміачна селітра, сульфат і нітрат амонію);
· амідні (азот в органічній амідній формі — сечовина, кальцій ціанід).
Нітратний азот не вбирається ґрунтом, легко розчиняється у воді, проникає у глибші шари і швидко поглинається рослинами. Тому нітратні, а також аміачно-нітратні солі слід використовувати для підживлення рослин під час вегетації, а також для внесення у невеликих дозах у рядки та гнізда під час сівби. Нітратний азот може легко вимиватися з ґрунту в разі достатнього зволоження, особливо в умовах зрошення, а також на легких дерново-підзолистих ґрунтах, унаслідок чого в таких умовах можливі значні його витрати.
Аміачний азот вбирається ґрунтом і локалізується в місцях внесення добрива й вимивається тільки після переведення його в нітратну форму під впливом нітрифікації. Ці добрива слід вносити восени.
У виробничих умовах для успішного формування бобово-ризобіального симбіозу важливим моментом, поряд із внесенням мінеральних добрив, є зараження кореневої системи рослин бульбочковими бактеріями. Досягається це шляхом передпосівного обробітку насіння бобових культур бульбочковими бактеріями або через внесення їх у ґрунт як спеціального препарату — нітрагіну. Передпосівна інокуляція зерна нітрагіном — досить ефективний агроприйом, який забезпечує підвищення врожаю бобових культур на 5...30 %. Успіх нітрагінізації значною мірою залежить від штаму бактерій, внесення в ґрунт фосфорно-калійних добрив, а також сірки, кальцію і молібдену. Використовується також азотобактерин, який збагачує ґрунт азотфіксаторами, що вільно існують.
Ефективність дії фосфорних добрив залежить від властивостей ґрунту і особливостей культур, у разі правильного застосування яких приріст урожаю на 1 кг Р2О5 становить для зернових у середньому 5... 10 кг/га. Ці добрива не тільки підвищують урожай, а й прискорюють достигання сільськогосподарських культур. Особливо помітний вплив їх у більш ранні фази розвитку рослин. З фосфорних добрив найбільше використовують суперфосфат (звичайний порошкоподібний або гранульований і марганізований, а також подвійний), фосфатшлак мартенівський, фосфоритне борошно, знефторений фосфат.
Суперфосфат — найпоширеніше фосфорне добриво. Найкращі результати від його внесення на солонцюватих ґрунтах, бо на кислих за ефективністю він поступається нейтральним фосфорним добривам.
Фосфатшлак (фосфоритне борошно) слід вносити на кислих дерново-підзолистих і опідзолених ґрунтах, а знефторений фосфат — На дерново-підзолистих і опідзолених та вилугуваних чорноземних Ґрунтах.
За даними багатьох дослідів, приріст товарної продукції від 1 кг К2О (діючої речовини калійного добрива) для зернових культур становить 3,8 кг/га. До концентрованих калійних добрив належать калій хлорид, калійна селітра, калій сульфат, калімагнезія. Остання — особливо цінне добриво на легких дерново-підзолистих ґрунтах для культур, які негативно реагують на хлор [9].
До складних мінеральних добрив належать азотно-фосфорні: амофос, діамофос, нітрофос, амонізований суперфосфат; азотно-калійні: калійна селітра; азотно-фосфорно-калійні: нітроамофоски, діамонітрофоски, нітрофоски (останні найпоширеніші з усіх складних мінеральних добрив).
Нормами (дозами) добрив називають розраховану чи фактичну кількість добрив, яку вносять на 1 га. Як правило, норму виражають у кілограмах діючої речовини (N, Р2О5 і К2О), яку вказують за відповідного поживного елемента (наприклад, N60P80K90).
Щоб виразити норму внесення мінеральних добрив у їхній фізичній масі, слід задану норму в кілограмах поживних речовин розділити на відсотковий вміст поживної речовини у відповідному добриві.
Для того щоб окремі типи мінеральних добрив перерахувати на умовні одиниці поживних елементів (азотні добрива в перерахунку на амоній сульфат із вмістом 20,5 % N, фосфорні — на простий суперфосфат із вмістом 18,7 % Р2О5, калійні добрива — на калійну сіль із вмістом 41,6 % К2О) та фізичну масу, використовують такі коефіцієнти: азотні добрива — 4,88; фосфорні — 5,35; калійні добрива — 2,40.
Для переводу мінеральних добрив у 100 %-й вміст поживних речовин фізичну масу даного добрива множать на величину відсоткового вмісту в них поживних речовин і ділять на 100.
У складних добривах кількість поживної речовини визначають так само окремо за азотом, фосфором, калієм. Останнім часом усі поживні речовини в добривах, ґрунті та рослині виражають в елементарній формі, тобто не в формі оксидів, а у вигляді елементів. Наприклад, коефіцієнти перерахунку з оксидів на елементи пожив них речовин і, навпаки, для фосфору і калію такі: Р2О5 — 0,4364 Р; Р — 1,2911 Р2О5; К2О — 0,8301 К; К — 1,2046 К2О.
Визначаючи дози добрив, важливо враховувати потреби різних культур в окремих елементах живлення. Розрізняють господарський, винос, тобто такий, де поживні речовини у рослинах є частиною врожаю, і біологічний, що становить максимальну кількість поживних речовин, яку винесено з ґрунту протягом вегетаційного періоду. Різні культури значно відрізняються за величиною поживних речовин. Якщо озима пшениця для створення врожаю 40 ц/га потребує в середньому 132...150 кг азоту, 43...60 кг фосфору і 85...112 кг калію, то для вирощування 400 ц/га цукрових буряків ці цифри становлять відповідно 180...220; 60...80 і 220...300.
За науково обґрунтованого програмування врожайності і визначення оптимальних доз добрив слід враховувати такі основні умови:
• загальну потребу даної культури в поживних елементах залежно від запрограмованого врожаю (основної і побічної продукції) та умов вирощування;
• можливе використання рослинами поживних речовин ґрунту;
• техніку внесення добрив;
• коефіцієнти використання рослинами поживних речовин запланованих добрив;
• економічні й організаційно-господарські умови, які визначають економічну ефективність різних доз добрив.
В Україні в усіх природних зонах у разі внесення мінеральних добрив спостерігається певна закономірність ефективності дії добрив на різних ґрунтах:
• на дерново-підзолистих і підзолистих ґрунтах високу ефективність, якщо провести вапнування, дають азотні, фосфорні та калійні добрива, на невапнованих ґрунтах — високоефективне - фосфоритне борошно;
• на сірих лісових ґрунтах, вилугуваних і опідзолених глибоких Малогумусних чорноземах найефективніші азотні добрива, а на їхньому фоні — й фосфорні та калійні;
• на звичайних, карбонатних і південних чорноземах високий ефект дають фосфорні добрива і середній — азотні; дія калійних добрив дещо слабша;
• на темно-каштанових і каштанових ґрунтах можна вносити у рядки фосфорні добрива.
Слід зауважити, що в умовах зрошення на цих ґрунтах високоефективні всі добрива. Для одержання високоякісних врожаїв сільськогосподарських культур, крім мінерального живлення, необхідно забезпечити оптимальний водний режим і найкращі умови для функціонування фотосинтетичного апарату рослинного організму.
Розділ 2. Класифікація добрив 2.1 Мінеральні добрива
Азотні добрива. Азот входить до складу найважливіших рослинних сполук, що відіграють провідну роль у всіх процесах життєдіяльності. . Він міститься в білках, нуклеїнових кислотах, фосфатидах, пептонах, поліпептидах, амінокислотах, хлорофілі, ферментах, токсинах, антитоксинах, вітамінах та ін. Нестача азоту призводить до зниження вмісту в рослині сполук, які містять азот, і в результаті — до порушення нормального перебігу життєвих процесів. Особливо сильно нестача азоту позначається на рості рослин. В результаті підсилення азотного живлення рослини розвивають міцну вегетативну масу, в них збільшується вміст білка, спостерігається загальне зростання урожайності. Регулюючи азотне живлення рослин, можна не тільки впливати на абсолютну величину врожаю, а й на його якість та структуру. Так, під час посиленого живлення рослин азотом збільшується кількість зеленої маси (наприклад, при вирощуванні силосних культур) і білка (наприклад, у зерні пшениці). Навпаки, при зниженні азотного живлення зменшується вміст азотних речовин у рослинах (наприклад, високий вміст азотних речовин в цукровому буряку знижує його якість).
Азотне живлення рослин регулюють переважно застосуванням різних мінеральних азотних добрив. Азотні добрива за своїм складом і агрономічними властивостями дуже неоднорідні. Найчастіше використовують, тверді азотні добрива, рідше — рідкі (аміачна вода, аміакати). Проте дуже перспективним і економічно вигідним добривом є рідкий аміак. За формою азотні мінеральні добрива поділяють на нітратні (в яких азот знаходиться у формі NO3-), аміачні (з азотом у формі NH4+), аміачно-нітратні (азот як у формі NO3-, так і NH 4+)та амідні добрива (форма азоту NH2-).
Аміакати — це розчини різних азотних добрив (аміачної, кальцієвої селітр або сечовини) в аміаку. При цьому досягається збільшення вмісту азоту до 35 % і більше. Проте аміакати сильно роз'їдають метали, тому зберігають їх в особливих цистернах з алюмінію, нержавіючої сталі або із спеціальними покриттями. Це утруднює транспортування їх і внесення в грунт.
В рідкому аміаку, аміачній воді і аміакатах міститься вільний аміак, який може легко втрачатися, тому ці добрива треба вносити у вологий грунт на глибину 10—12 см. Зменшення глибини загортання або поверхневе їх внесення призводять до значних втрат азоту.
Фосфорні добрива. Загальний вміст фосфору (Р2О5) в рослинах змінюється в межах 0,2 — 1,5 %. Фосфор у рослинах входить до складу молекул складних білків (нуклеопротеїдів), нуклеїнових кислот, фосфатидів, фітину, ферментів та ін. В рослинах фосфор зумовлює нормальний перебіг багатьох найважливіших процесів. При нестачі його порушується синтез вуглеводів (наприклад, знижується крохмалистість бульб картоплі, цукристість цукрових буряків тощо), уповільнюється розвиток рослин (дозрівання їх, розвиток кореневої системи), утворення репродуктивних органів, знижується зимостійкість озимих культур, погіршується якість урожаю та ін. Валовий вміст фосфору в ґрунтах змінюється в межах 0,1 — 0,25 %, проте значна частина його міститься у важкорозчинних сполуках, тому рослини часто відчувають нестачу його, яка поповнюється внесенням добрив.
Як фосфорні добрива застосовують різні сполуки — одно-, дво- і тризаміщені кальцієві солі ортофосфорної кислоти (Са(Н2РО4)2, СаНРО4 та Са3(РО4)2). Із збільшенням ступеня заміщеності розчинність цих солей зменшується, зберігаючись, проте, в достатньо високих межах при дії на них кислих розчинників. Тому особливості застосування різних фосфорних добрив багато в чому залежать від форми фосфату, розчинності його, а отже, і міри доступності рослинам фосфору. Наприклад, доступність фосфору суперфосфату, в якому він міститься у формі Са(Н2РО4)2, значно вища, ніж доступність фосфору фосфоритного борошна— Са3(РО4)2. Проте на кислому фоні, де досить рухливі навіть важкорозчинні фосфати, фосфоритне борошно в середньому не менш ефективне, ніж суперфосфат.
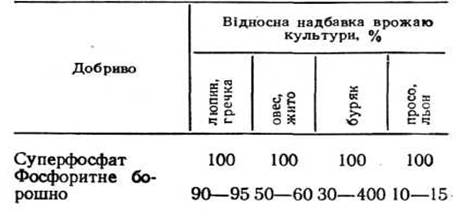
Доступність фосфору різних за ступенем розчинності фосфатів багато в чому залежить також від біологічних особливостей самих рослин (табл. 2.1).
Таблиця 2.1 Відносна ефективність різних фосфатів при внесенні їх під різні культури
Калійні добрива. Середній вміст калію в рослинах (з розрахунку на К2О) змінюється в межах 0,5 — 2 %. Особливо багато калію в молодих частинках рослин. Тут ця кількість може досягати 50 % загального вмісту зольних елементів. Проте в рослинах калій не утворює тривких органічних сполук і в значній частині перебуває в клітинному соці в рухливому вигляді. Валовий вміст калію в ґрунтах звичайно змінюється в межах 1 — 2 %, проте доступного рослинам калію значно менше.
Як калійні добрива використовують різні хімічні сполуки, які містять переважно хлорид або сульфат калію. Нерідко ці сполуки містять значну кількість баласту у вигляді NaCl, Na2SO4 та залишків сильних кислот. Це призводить до того, що при внесенні цих добрив в кислі, не насичені основами ґрунти активна кислотність ґрунтів сильно зростає. Небажаним компонентом калійних добрив є хлор. Хлор негативно впливає на деякі рослини. Так, при високому вмісті в ґрунті хлору знижується цукристість винограду і пригнічується розвиток його, погіршується якість тютюну. Чутливі до хлору льон, картопля, гречка, конюшина, ягідні рослини, коноплі і деякі інші культури. Щоб запобігти негативній дії на рослини хлору, калійні добрива, як правило, вносять у грунт восени під зяблеву оранку або рано навесні, до розмерзання ґрунту. В цьому разі в осінній і весняний періоди інтенсивного зволоження ґрунту хлор з нього вимиватиметься, оскільки він, як аніон, ґрунтовими колоїдами не вбирається. При внесенні калійних добрив під культури, які дуже страждають від наявності хлору, перевагу слід надавати сульфатам або тим добривам, в яких вміст хлору найменший. Останні доцільно використовувати і для підживлення. Раціональний вибір добрива дає можливість істотно підвищити урожайність і поліпшити якість продукції.
Мікродобрива. Мікродобрива містять мікроелементи, тобто ті елементи, які потрібні рослинам в малих кількостях (десятки грамів на гектар). До таких елементів належать бор, марганець, мідь, молібден, кобальт, цинк, залізо та ряд інших. Тепер як мікродобрива застосовують найрізноманітніші речовини.
Складні й змішані добрива. Складними називають добрива, які містять кілька (2—3 і більше) елементів живлення рослин. Нерідко їх, в свою чергу, поділяють на три групи: власне складні добрива, комбіновані (або комплексні) і змішані добрива. До складних належать добрива, в яких елементи сполучаються хімічним способом: калійна селітра — KNO3, амофос — NH4H2PO4, діамофос — (NH4)2HPO4. Комбінованими називають добрива, елементи яких сполучаються не тільки хімічним, а й фізичним способом (наприклад, сплавлюванням): амонізований суперфосфат (NH4H2PO4+ CaSO4), який дістають насиченням простого суперфосфату аміаком. Важливими представниками цієї групи є нітрофоски, які містять азот, фосфор і калій. Нітрофоски дістають сплавлюванням і дальшою обробкою суміші різних компонентів, (наприклад, сплавлюванням фосфориту, азотної кислоти, хлориду калію та ін.). Змішаними добривами називають суміші деяких простих добрив. Ці суміші готують як безпосередньо в господарствах, так і на хімічних підприємствах. Останній спосіб економічно вигідніший. Характерною особливістю змішаних добрив є те, що в них повністю зберігаються баласти простих добрив, які використовувались для виготовлення суміші. Виготовляти суміші можна тільки з деяких добрив. Іноді при змішуванні добрив утворюються токсичні для рослин сполуки (наприклад, СаС12) або знижується доступність потрібного рослині елемента(наприклад, при змішуванні суперфосфату з вапном рухливість фосфору значно зменшується), або погіршується фізичний стан добрив (наприклад, при змішуванні селітр утворюється не-сипка маса, яку важко вносити в грунт). Тому при змішуванні враховують правила змішування. Складні добрива класифікують також за їхнім складом на калійно-азотні (наприклад, KNO3), азотно-фосфорні (амофос, діамофос, амонізований суперфосфат) та ін. Крім того, є складні добрива, до яких входять як макро-, так і мікроелементи. Прикладом таких добрив є марганізований суперфосфат, марганізована аміачна селітра, молібденовий суперфосфат та ін.
Усі складні добрива містять елементи живлення в легкозасвоюваній рослинами формі.
Цінним місцевим складним добривом є зола. В ній містяться до 5 — 7 % Р2О5 до 10—13 % К2О, до 40 % і більше СаО, а також мікроелементи (табл. 2.2.).
Таблиця 2.2 Середній вміст фосфору, калію і кальцію в золі деяких рослин, %
| Рослина | Р2О5 | К2О | СаО |
| Стебла соняшнику і солома гречки Житня і пшенична солома Дрова Торф | 2—4 4—7 2—7 1—2 | 30-35 10—14 3—14 0,5—2 | 18—20 8—10 30—40 15—25 |
Похожие работы
... . Йому належать відкриття анаеробної фіксації азоту і з'ясування ролі мікроорганізмів ґрунту в перетворенні гумусових речовин. Важливими в теоретичному і практичному відношеннях для мінерального живлення рослин з'явилися роботи основоположника радянської школи агрохімії Д Н. Прянишникова (1865-1948). Він установив, що правильне використання мінеральних добрив є могутнім чинником регулювання фізі ...
... локалізація всіх трьох елементів живлення. Річну дозу добрив під окремі культури можна вносити в різні терміни і різними способами. Терміни і прийоми внесення добрив повинні забезпечувати якнайкращі умови живлення рослин протягом всієї вегетації і отримання найбільшої окупності живильних речовин урожаєм. Розрізняють три способи внесення добрив: допосівне (або основне), припосівне (в ряди, кубла, ...
... культури. Відповідно до мети досліджень у задачі досліджень входило: 1) вивчення сучасного стану інтродукованих видів рододендрон в Житомирському Поліссі; 2) розробка рекомендацій щодо інтродукції, збереження та використання в Житомирському Поліссі; 3) дослідження біологічних особливостей виду рододендронів; 4) застосування набутих знань для інтродукції рододендронів; 5) використання ...
... на забезпечення раціонального використання земельних ресурсів, охорону ґрунтів і навколишнього середовища. В умовах пересіченого рельєфу та на ерозійно небезпечних територіях необхідне освоєння ґрунтозахисних систем землеробства з використанням контурно-меліоративної організації території, яка здатна запобігти ерозії найнебезпечніших в екологічному відношенні земель. При цьому надають перевагу ...
0 комментариев