Навигация
Обособление и последующее формирование материнских почек происходят в период роста побегов, процессы формирова-
40518
знаков
0
таблиц
0
изображений
2. Обособление и последующее формирование материнских почек происходят в период роста побегов, процессы формирова-
* Материнскими называются почки, формирующиеся на растущих или закончивших рост побегах. Почки возобновления, которые закладываются (образуются) в материнских почках, называют дочерними. После распускания материнской почки и вырастания из нее побега продолжения заложенные на этом побеге дочерние почки сами становятся материнскими.
** Генеративные побеги связаны только с образованием генеративных органов; закладываются они в функционально генеративных (цветковых) почках и после окончания цветения или созревания семян отмирают. Генеративно-ростовые побеги служат для образования как генеративных, так и вегетативных органов, они закладываются в генеративно-ростовых (смешанных) почках. После окончания цветения или созревания семян на побегах этого типа отмирает их генеративная часть, а вегетативная сфера сохраняется для выполнения функций вегетативных побегов.
*** У голосеменных растений фазе бутонизации соответствует фаза обособления на побегах микро- и макростробилов, а фазе цветения — фаза пыления.
ния почек возобновления обычно завершаются после окончания роста побегов. У целого ряда древесных пород (кипариса, туи, платана, белой акации, актинидии) фенологические фазы формирования почек внешне не выражены.
3, Генеративные, а у таких родов древесных растений, как рябина, боярышник, калина, и генеративно-ростовые почки весной трогаются в рост раньше вегетативных.
4. Большим фенологическим разнообразием отличаются древесные растения по соотношению во времени прохождения циклов вегетативного и генеративного развития. Так, по периодам цветения различают древесные растения, цветущие до распускания листьев (осина, виды ольхи, лещины, вяза; до распускания молодой хвои пылит сосна), в начальный период облиствения (виды березы), вскоре после массового облиствения (дуб черешчатый, бук лесной, виды ореха), после окончания роста побегов — в середине лета (виды липы) или только в конце лета—осенью (аралия маньчжурская, целебник).
Особую биологическую группу составляют виды с ремонтантным цветением — очень продолжительным, циклически многократным. Такой тип цветения присущ растениям тропиков, но может наблюдаться и у ряда видов древесных растений умеренных широт (роза морщинистая, курильский чай). Иногда у древесных растений происходит вторичное цветение — преждевременное зацветание в конце лета или осенью цветков, заложенных в почках для будущего года (ива козья, конский каштан, слива, вишня, яблоня).
Значительно различаются между собой древесные растения разных родов и видов по времени созревания плодов и семян. Например, у осины и многих видов ивы семена созревают в конце весны: у вяза — в начале лета, у вишни, смородины, березы повислой — в середине, у рябины —- в конце лета. Плоды дуба черешчатого и бука, семена сосен обыкновенной и кедровой сибирской, пихты, ели созревают в начале осени, плоды кленов остролистного и ложноплатанового, клюквы — в середине осени, а липы и ольхи — в конце. Еще более различны у древесных растений разных фенологических групп периоды распространения зрелых плодов и семян.
Далеко не одинаковы у древесных растений и циклы формирования плодов и семян. У ивы, тополя и вяза их созревание происходит через 3—6 недель после зацветания, у дуба черешчатого и скального — через 3—4 месяца, у сосны, кипариса и дуба из секции красных дубов (дубы красный, шарлаховый и др.) — только через 1.5 года.
В связи с изменениями климатических и других условий внешней среды у одних и тех же видов и форм древесных растений одноименные фенофазы могут наступать в совершенно различные сроки как в одном географическом пункте, так и в разных. Поэтому регулярные многолетние фенологические наблюдения очень важны для установления фактических местных периодов наступления фенофаз у изучаемых растений. Эти же фенологические наблюдения позволяют выявлять истинную длительность разного рода фенологических циклов, прежде всего циклов вегетации и покоя.
Вегетация — это такое состояние растений, при котором происходят процессы видимого роста вегетативных и генеративных органов, и осуществляется непрерывная ассимиляционная деятельность листьев.
Покой — период (во временном отношении — цикл) в годичном цикле развития растений, когда видимый рост отсутствует, а листья не ассимилируют, в том числе и у вечнозеленых растений (а если ассимилируют, то только в отдельные дни и часы с достаточно высокой для осуществления фотосинтеза температурой воздуха).
У листопадных видов древесных растений фенологическим индикатором начала вегетации является распускание вегетативных почек, а окончания ее — полное осеннее расцвечивание листьев в кроне или их опадание, если листья опадают зелеными (сирень обыкновенная, жостер слабительный, ольха серая, часто ясень обыкновенный). У вечнозеленых видов фенологические признаки начала и окончания вегетации менее определенны и разными учеными понимаются неодинаково (Малышева, 1973; Елагин, 1976, и др.). Так, Н.Е. Булыгин начало вегетации вечнозеленых древесных растений устанавливает по дате массового набухания вегетативных почек (когда начинают расти корни и обнаруживаются анатомические признаки образования камбиального кольца), а окончание — по совокупности фенологических признаков у разных групп растений. Например, у видов сосны — по завершению опадания пучков отмершей хвои вместе с укороченными побегами (брахибластами), у видов туи — по прекращению веткопада, у вересковых — по окончанию осеннего расцвечивания листьев (брусника, клюква) или по свертыванию листьев вдоль центральной жилки (багульник, рододендрон). Если же четко выраженные признаки окончания вегетации отсутствуют (виды ели, пихты, лжетсуги, тсуги, можжевельника и др.), то ориентировочно считается, что вегетация заканчивается одновременно с завершением листопада у таких наиболее длительно вегетирующих зимнеголых деревьев местной флоры, как ольха черная (к югу от подзоны средней тайги) или ольха серая (средняя и северная тайга, лесотундра).
Следует различать понятия «период (цикл) вегетации» и «вегетационный период». Период вегетации — явление биологическое и означает время вегетирования растения или растительного сообщества (фитоценоза). А вегетационный период — явление географическое и может быть как метеорологическим (сообщается в официальных климатических справочниках), так и фенологическим (указывается в фенологической литературе). В метеорологическом смысле он выражает период между датами перехода весной и осенью среднесуточной температуры воздуха через пороговые значения (+5 °С), в фенологическом — заключает период между датами наступления фенофаз-индикаторов вегетационного сезона. В таежной зоне — это начало пыления ольхи серой—полное пожелтение листьев березы (повислой, плосколистной или пушистой), южнее — между датами начала пыления орешника (лещины) и завершением листопада у березы (Шульц, 1980; Булыгин, 1997).
Растения со сходными сроками начала и окончания вегетации и близкой продолжительностью циклов вегетации и покоя объединяют в определенные фенологические группы, получившие название феноритмотипа. Автор этого термина И.В. Борисова (1965, 1972) разработала стройную систему феноритмотипов сосудистых растений. Однако все многообразие фенологических групп древесных растений она свела к двум феноритмотипам — вечнозеленым и листопадным. На самом деле их, конечно, значительно больше. Так, во влажно-тропических лесах один феноритмотип составляют вечнозеленые, непрерывно вегетирующие деревья и кустарники. У них не бывает периодов покоя, а наблюдается чередование циклов роста побегов и временной приостановки его. Иной феноритмотип составляют вечнозеленые древесные растения умеренного климатического пояса: они вегетируют летом, зимой находятся в покое, весенний сезон для них является периодом перехода от состояния покоя к вегетации, а осенний — от вегетации к покою.
Свои феноритмотипы имеют листопадные растения. Одни из них характеризуются ежегодным чередованием одного цикла вегетации и одного цикла покоя (например, древесные породы мелко-и широколиственных лесов Российской Федерации), другие же могут вегетировать и впадать в покой (вынужденный) несколько раз за год. Так, по свидетельству Г.Вальтера (1968), в субтропической пустыне Соноре (Северная Америка) есть виды кустарников, вегетирующих до 10 раз в году и столько же раз вступающих в покой. Такая многоцикличность вегетации и покоя обусловлена периодичностью выпадения дождей: как только почва достаточно увлажнится, растения начинают вегетировать, а с прекращением дождей и наступлением очередного засушливого периода они сразу сбрасывают листья и впадают в покой.
Обобщая фенологические особенности древесных растений различных биологических групп, Н.Е. Булыгин (1998) предложил следующую иерархическую систему дендрофенологических единиц (таксонов): класс — подкласс — порядок — фенологическая группа—дендроритмотип—субдендроритмотип — фенологическая вариация. Основным таксоном здесь является дендроритмотип — дендрологический аналог феноритмотипа. Это — совокупность видов и форм древесных (и полудревесных) растений со статистически сходными сроками (т.е. обоснованными сроками с применением методов математической статистики) начала и окончания вегетации. Высший таксон — класс. Их всего два: вечнозеленые и листопадные (сезонноголые или летне-зеленые) растения. Подклассов три: непрерывно вегетирующие растения (во влажных тропиках), чередующие циклы вегетации и вынужденного покоя (в сухих тропиках и субтропиках) и чередующие циклы вегетации с глубоким (органическим) покоем (в умеренном и холодном климатических поясах). Порядки выделяют по числу циклов вегетации за астрономический год. Для климатических условий России типичен один годовой цикл вегетации, в более низких широтах, с иными климатическими условиями годовых циклов вегетации у древесных растений их может быть два и более (см. примеры выше). Фенологические группы устанавливают по среднемноголетним датам начала и окончания вегетации, выделяя растения с ранними, средними, поздними и очень поздними сроками, так как в различных условиях местопроизрастания календарные даты начала и окончания вегетации могут существенно различаться; при выделении феногрупп применяют определенные фенологические индикаторы. Например, в лесной части европейской территории России к группе рановегетирующих растений (Р) относят те, которые начинают вегетировать до зацветания ивы козьей или осины, к группе средневегетирующих (С) — до распускания листьев калины красной, к поздневегетирующим (П) — до зацветания черемухи обыкновенной, а к начинающим вегетировать очень поздно (ОП) — позже зацветания черемухи.
По периодам завершения вегетации к группе «ранних» относят те растения, у которых эта фаза наступает раньше или одновременно с ивами трехтычинковой и ушастой, с березой пушистой наиболее ранних биотипов; к группе «поздних» — оканчивающие вегетацию после прекращения листопада у березы повислой. По показателям этих двух феногрупп и определяют соответствующий дендроритмотип.
Среди вечнозеленых растений естественной дендрофлоры Российской Федерации имеется всего два дендроритмотипа: ранне-поздний (Р.П) и средне-поздний (СП — у позднораспускающихся форм елей европейской и сибирской, клюквы болотной и др.). У зимнеголых древесных видов состав дендроритмотипов значительно многообразнее. Здесь, например, имеются такие дендроритмотипы, как Р.Р (ранний-ранний) — голубика; П.С (поздне-средний) — береза повислая; С.С (средне-средний), П.С (поздне-средний) и ОП.П (очень поздний-поздний) — дуб черешчатый и другие виды. Из фенологических вариаций дендроритмотипов можно отметить две: выделяемые по соотношению периодов роста побегов и листораспускания (у ели, пихты, березы и подавляющего большинства других древесных пород эти процессы идут одновременно, а у видов сосны — вначале растут побеги, а распускание хвои происходит непосредственно перед окончанием их роста), а также по числу циклов роста побегов за период вегетации: для деревьев и кустарников нашей страны типичен один цикл роста, но у ряда видов рост многоцикличный, как у тропических древесных растений. Такой рост, например, наблюдается у дуба, образующего второй и третий приросты (так называемые «ивановы побеги»), у роз морщинистой и иглистой, у вереска и дрока.
Субдендроритмотипы отражают специфику генеративного цикла развития древесных растений. Их выделяют по соотношению периодов листораспускания и цветения (у голосеменных — пыления) и по продолжительности периода формирования урожая от зацветания до созревания плодов и семян. Древесные растения, цветущие до распускания листьев, составляют вариацию (группу) 1Ц (сосна, ольха, лещина), одновременно с началом распускания листьев — 2Ц (ель, пихта, береза), после массового их распускания — ЗЦ (дуб, орех), цветущие ремонтантно — ЗЦр (вследствие многоцикличного роста побегов у розы морщинистой, снежноягодника, вереска и др.), цветущие после листораспускания и окончания роста побегов — 4Ц (аралия, диморфант, липа). По длительности периода формирования урожая выделяют фенологические вариации с коротким (К) периодом (в 52 дня и менее: вяз, ива, тополь, малина), средним (С — от 53 до 80 дней: березы повислая и пушистая, брусника, черника), длительным (Д — от 81 до 105 дней: березы даурская и ребристая, клюква болотная, роза собачья), очень длительным (ОД — от 106 до 130 дней: ель, пихта, бук, дуб) и сверхдлительным (СД — свыше 130 дней: кипарисовик, граб, клен остролистный, лещина). Дополнительную группу составляют древесные растения, у которых плоды и семена вызревают во второй (2с: виды сосны, кипариса, можжевельника, дуба из секции красных дубов) или даже третий сезон вегетации (Зс: в крайне суровых климатических условиях — у некоторых видов сосны).
Пользуясь приведенными критериями и условными обозначениями дендроритмотипов и субдендроритмотипов, можно простой фенологической формулой отразить наиболее существенные особенности сезонного развития любого древесного вида или его формы. Так, фенологическая формула для ели аянской будет такой: Р.П/2Ц.ОД; для елей обыкновенной и сибирской, пихты сибирской — Р.П,С.П/2Ц.ОД, для сосен обыкновенной, кедровой сибирской и кедровой стланиковой — Р.П/Щ.2с, для осины — П.Р,П.П,ОП.С/1Ц.К, для ясеня обыкновенного — П.С,ОП.П/1Ц.ОД, для розы морщинистой — С.П/ЗЦр.С, для винограда амурского — С.С,П.С/ЗЦ.Д и т. д.
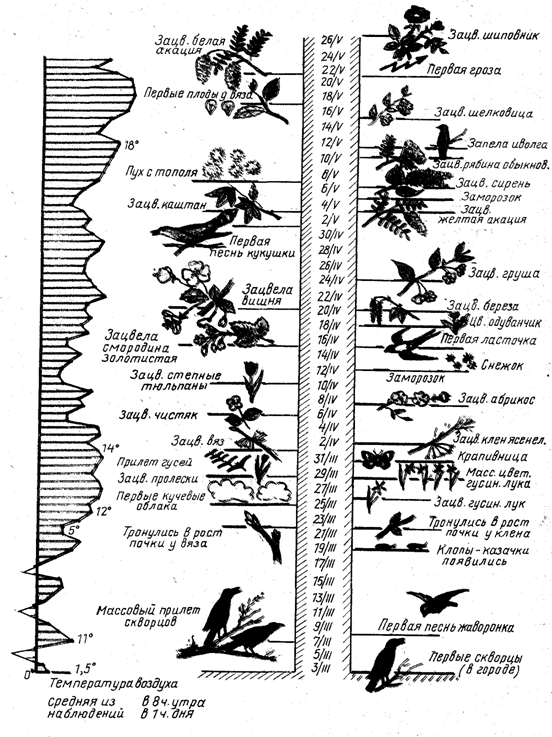
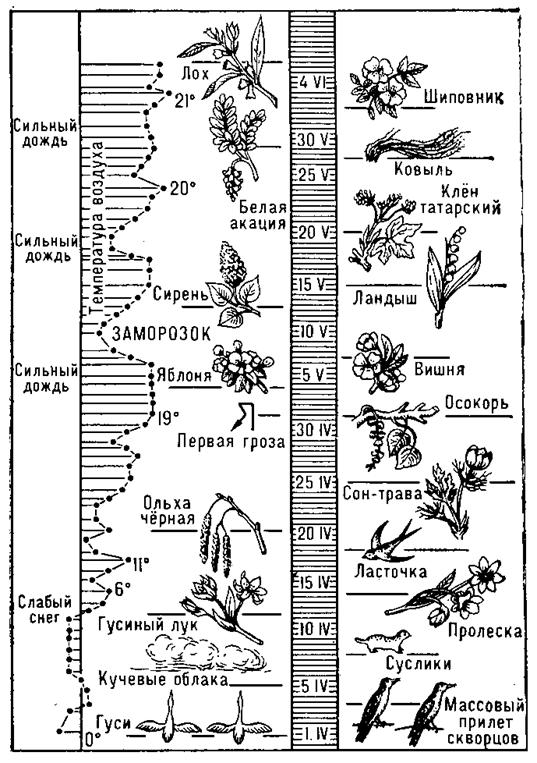
Характерные особенности сезонного развития различных видов и фенологических форм хвойных и лиственных древесных растений России наглядно показаны на помещенных в Приложениях 9.1 и 9.2 феноспектрах — графических изображениях последовательности наступления и прохождения растением фенологических фаз и циклов сезонного развития.
Фенологические наблюдения над древесными растениями (дендрофенонаблюдения) требуют четкого знания методики их проведения и особенно диагностических (морфолого-биологических) признаков наступления регистрируемых фенофаз. С этими признаками, условными обозначениями фенофаз, методикой проведения наблюдений и оформления их результатов рекомендуется знакомиться по специальным фенологическим руководствам (Фенологические наблюдения..., 1973; Плотникова, 1973; Булыгин, 1974, 1976а, 1979; Методика фенологических..., 1975; Елагин, Лобанов, 1979; Зайцев, 1981; Булыгин и др., 1982). Образец отчетного бланка учебных ден-дрофенологических наблюдений приведен в Приложении 12.
Современная дендрофенология изучает не только динамику внешне выраженных фенологических (макрофенологических) процессов, но и ритмику формирования зачаточных побегов в почках, образования годичного кольца древесины, заложения и формирования зачаточных органов семени. Весь этот круг исследований, выполняемых с использованием бинокулярных луп и микроскопов, относят к микрофенологии (Фенологические наблюдения..., 1973; Булыгин, 1982), непосредственно смыкающейся с задачами изучения морфогенеза растений (морфогенез — процесс формообразования, т.е. заложения, роста и развития клеток, тканей, органов).
Например, с помощью таких глубоких микрофенологических (анатомо-морфологических) исследований было выявлено, что цикл формирования побегов древесных растений от заложения их в почках до окончания роста у одних видов может охватывать 3 сезона вегетации (ель, пихта, сосна, дуб, ясень), у других — 2 (липа), а у лиственницы — 2 и 3. Было также установлено, что по соотношению вегетационных периодов, в которые у древесных растений закладываются в почках цветочные зачатки и в которые растения впоследствии цветут, можно выделить 4 основные биологические группы видов. Самую многочисленную из них составляют древесные растения, у которых зачаточные цветки образуются в почках в год, предшествующий цветению (все виды ели, пихты, лиственницы, сосны, ясеня, березы, яблони, груши и др.). Ко второй группе относятся растения, образующие зачаточные цветки в год цветения (аралия, диморфант, целебник, липы войлочная и крымская, вереск). Третья группа объединяет древесные растения, образующие зачаточные цветки как во второй половине лета и осенью в год, предшествующий цветению, так и весной в год цветения (липы крупнолистная и мелколистная, кизильник блестящий, клен татарский, белая акация). Четвертую группу составляют однодомные деревья с раздельнополыми цветками, тычиночные из которых образуются в почках в середине лета, предшествующего году цветения, а пестичные могут образовываться как осенью предыдущего года, так и весной в год цветения: виды дуба, а в определенных климатических условиях — и виды ореха (Булыгин, 1963, 1964).
Древесные растения 1-й группы цветут весной до распускания листьев или в начальный период их распускания, 2-й группы — во второй половине сезона вегетации, после завершения роста побегов. Представители 3-й и 4-й групп цветут в конце весны—в начале лета, после массового распускания листьев. Следует заметить, что видам третьей группы присуще ремонтантное цветение.
Существуют также специальные программы и методики, связанные с изучением динамики сезонного роста корней древесных растений. С этой специфической областью дендрофенологии можно ознакомиться по руководствам В. А. Колесникова (1972), Г. Д. Ярославцева (Фенологические наблюдения..., 1973) и П. К. Красильникова (1983).
Заключение
Исследования многих ученых показывают, что у древесных растений имеются определенная сопряженность и взаимосвязь между динамикой внешне выраженных фенологических фаз и ритмом глубинных микрофенологических процессов, включая рост и формирование корневых систем. Всестороннее изучение механизмов этих биоритмических связей на разных уровнях их проявления (от биохимического до морфологического, от клеточного до организменного) является одной из актуальнейших задач современной дендрологии.
Похожие работы

... ), определите, что из нее вам подходит и что вы можете добавить в нее своего, после чего составьте собственную программу. Оформите в тетради в клеточку. План проведения занятия. Тема занятия: Методика проведения фенологических наблюдений. Цель занятия: освоить методику организации и проведения фенологических наблюдений. Задачи занятия: 1. Научиться методически правильно организовывать ...
... биологических и химических средств защиты леса от хвое- и листогрызущих насекомых». Наставление разработано на основе обобщения результатов исследований по совершенствованию технологий изменения средств защиты леса от вредителей на базе современной авиационной техники. Данный документ рассматривает такие важные для лесозащитной практики вопросы, как определение целесообразности авиационной борьбы ...
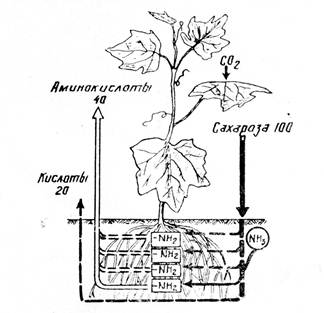

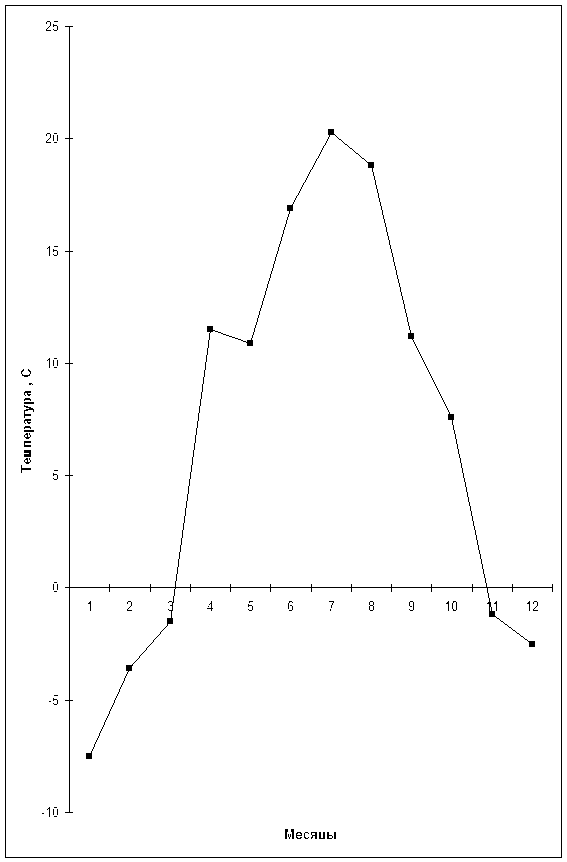
... разница по отношению к контролю (Соузга, НСР05 = 0,010) несущественна для исследуемых нами сортов и форм. Некоторую разницу в общем содержании воды в листьях калины исследуемых сортов и форм можно объяснить разными погодными условиями за период вегетации (графики 3.1, 3.2): начало вегетации (конец апреля – май) характеризовался сравнительно высокими для данного периода температурами и небольшим ...
... " г. Сочи куртина 64 23,3±0,49 34,7±1,5 6,3±0,28 2,8±0,25 4,67±0,12 4,67±0,12 Таксационные показатели были обработаны на ЭВМ методом вариационной статистики (приложение №4). 7. Опыт интродукции сосны крымской и проект ее внедрения в лесные культуры в Лооском лесхозе Впервые сосна крымская была введена в культуру в Англии в 1790 году семенами полученными от академика Полласа. В ...
0 комментариев