Навигация
Реферат
на тему:
«Зародыши и предки»
План
1. Проблема морофологии
2. Отногенез, филогенез и реапитуляция
3. Механика развития и менделевская генетика
4. Генетика развится
1. Проблема морофологии
Вероятно, мне следует пояснить, — добавил барсук, нервно опуская свои бумаги и глядя поверх них на бородавку, — что все зародыши выглядят в общем одинаково. Зародыш - это то, что вы есть прежде, чем вы родитесь на свет. И станете ли вы в будущем лягушкой или павлином, жирафом или человеком, пока вы остаетесь зародышем, вы похожи всего лишь на омерзительное и беспомощное человеческое существо. Итак, я продолжаю:
Зародыши стояли перед Господом, вежливо сложив свои слабые ручонки на животах и почтительно свесив вниз тяжелые головы, и Господь обратился к ним. Он сказал: «Ну так вот, зародыши, все вы пока выглядите совершенно одинаково. Но Мы дадим вам возможность самим решать, кем вы хотите быть. Когда вы станете взрослыми, вы так или иначе вырастите, но Нам приятно наделить вас еще одной способностью. Вы можете заменять любые свои части такими, какие, по вашему мнению, окажутся вам полезными в будущей жизни».
Т. Уайт «Бывший и будущий король»
Для всех организмов характерно то или иное строение, и все они обладают определенными типами поведения и физиологических адаптации. В перспективе долгих эр геологического времени эти характеристики проявляют способность к видоизменению, почти достойную Протея: лопастной плавник кистеперых становится конечностью амфибии, крылом птицы, рукой и кистью человека. Это зримое достижение эволюции. Каковы же механизмы, при помощи которых совершаются эволюционные изменения морфологии?
Ответ на этот вопрос нам, в сущности, уже известен, во всяком случае в формальном смысле. Гарстанг (Garstang) дал его еще в 1922 г., обратив внимание на то, что эволюционный ряд, или филогения, это не просто последовательность взрослых форм. Каждое поколение взрослых особей возникало в результате последовательных процессов развития - онтогенеза - от, казалось бы, лишенного структуры яйца до сложной морфологии взрослого организма. Таким образом, для того чтобы некое эволюционное изменение проявилось в виде изменения структуры взрослого организма, некая новая морфология, некое измерение должно возникнуть в онтогенезе.
Можно было бы ожидать, что роль процессов развития в эволюции составляет один из главных компонентов современных эволюционных исследований; однако это не так. Эмбриональное развитие, составлявшее столь важную часть эволюционной теории в конце XIX в., в XX в. стало рассматриваться как не очень существенное. Позже в этой главе мы обсудим причины такого странного отчуждения. Конечно, значение зависимости между развитием и эволюцией никогда не было целиком предано забвению. Гарстанг, Гексли, де Бер и Гольдшмидт (Goldschmidt) определенно уделяли серьезное внимание этой зависимости в период 1920-1950-х годов. А сравнительно недавний выход книги Гулда (Gould) «Онтогенез и филогенез» показывает, что интерес к этой теме остается не только живым, но и острым.
Наше собственное увлечение этой проблемой разгорелось некоторое время назад под влиянием книги де Бера (de Beer) «Зародыши и предки», в которой так убедительно доказывается, что изменения сроков наступления различных процессов развития могут иметь глубочайшие эволюционные последствия. К сожалению, де Бер (de Beer) ограничился лишь кратким общим рассмотрением генов, регулирующих скорости процессов развития, уделив мало внимания роли генетической регуляции в развитии или эволюции. В то время когда де Бер (de Beer) писал свою книгу, первое издание которой вышло в 1930 г., о генетике развития было просто слишком мало известно, чтобы он мог излагать ее достаточно глубоко. К 1958 г., когда вышло третье и последнее издание «Зародышей и предков», о ней стало известно гораздо больше, но де Бер привел очень немногие из результатов, достигнутых генетикой развития после 30-х годов. Его основные интересы лежали в другой плоскости.
Фактически эмбриогенетические основы эволюционных изменений никогда подробно не разбирались. Именно ими мы и хотим заняться в этой книге. Наша исходная позиция состоит в том, что процессы развития находятся под генетическим контролем и что эволюцию следует рассматривать как результат изменений в генах, регулирующих онтогенез.
Интересно напомнить, что эту точку зрения впервые выдвинул в 1940 г. Гольдшмидт в своих «Материальных основах эволюции», хотя в то время о генах и об их функциях в развитии было известно слишком мало, чтобы получить успешный синтез эмбриологических и генетических данных. Идеи Гольдшмидта на протяжении последних 35 лет игнорировались из-за его своеобразного (и ошибочного) взгляда на природу генов, но сформулированное им определение эволюции дает совершенно ясное представление о теме этой книги:
«Эволюция означает переход одной достаточно стабильной органической системы в другую, но также стабильную систему. Генетическая основа этого процесса - изменение некой стабильной генетической конституции и превращение ее в другую — лишь одна сторона проблемы. Никакая эволюция невозможна без первичного изменения в зародышевой плазме, т.е. преимущественно в хромосомах, приводящего к новой стабильной структуре. Однако у этой проблемы есть и другая сторона. Зародышевая плазма держит под контролем тип данного вида, регулируя процесс развития индивидуума... специфичность зародышевой плазмы - это ее способность обеспечивать протекание системы реакций, составляющих процесс индивидуального развития, в соответствии с некой постоянной программой, которая повторяется, ceteris paribus, с целенаправленностью и упорядоченностью автомата. Эволюция, следовательно, означает создание измененного процесса развития, регулируемого измененной зародышевой плазмой». Термин «зародышевая плазма», используемый Гольдшмидтом, означает генетический материал, т.е., пользуясь современной терминологией, ДНК генома.
Какого рода гены управляют онтогенезом и каким путем они участвуют в эволюции?
В настоящее время наиболее хорошо изучены гены, кодирующие различные специализированные виды РНК, или белки, жизненно важные для общей структуры и функции клеток; это рибосомные РНК, различные ферменты, структурные белки, как, например, тубулин или коллаген, или такие белки, как гемоглобин, служащие переносчиками других веществ. Оценки роли таких структурных генов для регуляции развития и морфогенеза колеблются в очень широких пределах. По нашему мнению, регуляторные функции структурных генов в процессах развития очень ограничены, однако высказывалась и прямо противоположная точка зрения. Примером морфогенетической гипотезы, приписывающей структурным генам и их продуктам весьма существенную роль, служит гипотеза, выдвинутая Моно (Monod) в его книге «Случайность и необходимость». По мнению Моно, структурная сложность возникает в результате того, что он назвал молекулярным эпигенезом белков. Под этим термином он понимал хорошо известную особенность белков, а именно, что аминокислотная последовательность данного белка определяет трехмерную конформацию, которую он принимает в среде данной клетки. Далее белки могут специфическим образом взаимодействовать с другими белками, образуя надмолекулярные структуры. Моно пишет: «Упорядоченность, структурная дифференцировка, приобретение функций - все это возникает из случайной смеси молекул, каждая из которых, взятая в отдельности, лишена какой бы то ни было активности или функциональной способности, за исключением способности узнавать партнеров, с которыми ей предстоит образовать определенную структуру». Далее он высказывает предположение, что этот процесс лежит в основе и служит парадигмой ряда автономных эпигенетических событий, объединяющихся и завершающихся развитием целостного организма. Доведенная до крайности эта идея вызывает в памяти эпигенетическую фантазию о том, что из смеси соответствующих макромолекул можно получить целую мышь.
Гипотеза Моно, даже не доведенная до крайности, неприемлема в качестве модели развития. И эволюцией структурных генов нельзя объяснить морфологическую эволюцию. Исследования Вилсона (Wilson А. С.) и его сотрудников показывают, что - во всяком случае применительно к таким ныне живущим группам организмов, как лягушки и млекопитающие, - эволюция структурных генов, кодирующих белки, имеет мало отношения к морфологической эволюции. Человек и шимпанзе быстро дивергировали морфологически, однако аминокислотные последовательности их белков на 99% одинаковы. В отличие от них у такой более древней группы, как лягушки, морфологическая эволюция протекает довольно медленно, но скорость эволюции их аминокислотных последовательностей сравнима с аналогичными скоростями у других организмов. На основании этих фактов Кинг (King) и Вилсон высказали предположение, что в основе морфологической эволюции, по всей вероятности, лежат изменения не структурных, а регуляторных генов.
Поскольку существует целая иерархия взаимодействующих контрольных механизмов, управляющих экспрессией генов и онтогенезом, регуляторные гены распадаются на ряд категорий, и дать им общее определение, как некой единой группе, труднее, чем определить структурные гены. Можно сказать, что в основном структурные гены обеспечивают поставку материалов, необходимых для развития, а регуляторные гены поставляют и расшифровывают рабочие чертежи. Структурные гены относительно легко исследовать, так как продукты, синтез которых они кодируют, нетрудно выделить, исследовать и определить их функции. Не удивительно, что найти подход к изучению регуляторных генов оказалось сложнее. Некоторые регуляторные гены или элементы не образуют никаких продуктов; другие образуют их, но лишь в чрезвычайно малых количествах. Наиболее хорошо известный пример - белок lac-репрессора (Е. coli); этот продукт одного из регуляторных генов контролирует экспрессию генов, определяющих метаболизм лактозы. В одной бактериальной клетке содержится всего 10 молекул репрессора.
Регуляторные гены функционируют на протяжении всего процесса развития, управляя онтогенезом тремя различными способами: во-первых, регулируя время наступления тех или иных событий; во-вторых, делая выбор из двух возможностей и тем самым определяя судьбу клеток или частей зародыша; в-третьих, интегрируя экспрессию структурных генов, с тем чтобы обеспечить создание стабильных дифференцированных тканей. Все эти три способа регуляции играют большую роль в эволюции.
Роль изменений в сроках наступления различных событий в процессе развития как важного и гибкого механизма для достижения существенной морфологической эволюции рассматривали де Бер (de Beer) в своем ценном труде «Зародыши и предки», а позднее Гулд (Gould) в книге «Онтогенез и филогенез». Эти авторы уделяли внимание не столько механизмам, осуществляющим генетическую регуляцию процессов развития, сколько определению типов возможных изменений в сроках событий, происходящих в онтогенезе, и демонстрации их эволюционных последствий. Различные эволюционные изменения рассматривались ими как последствия изменения этих сроков. Чаще всего в качестве таких примеров приводятся случаи неотении - возникновение новых планов строения взрослого организма в результате достижения личиночными стадиями половозрелости и утраты предковой взрослой стадии.
Генетическая регуляция онтогенеза не ограничена, однако, воздействием на продолжительность процессов развития. Недавними работами, в особенности на плодовой мушке Drosophila melanogaster, ставшей для исследователей структуры и функции генов за это десятилетие чем-то вроде эукариотической Е. coli, установлено, что организация развивающегося зародыша контролируется целой иерархией регуляторных генов. Эти гены действуют как переключатели, от которых зависит, по какому из двух альтернативных путей развития пойдет данная клетка или группа клеток. После того как решение принято, возможности клеток в смысле дальнейшего выбора оказываются ограниченными, и их судьба в процессе развития становится все более и более определенной. Регуляторные гены такого типа доступны изучению благодаря очень ярко выраженным эффектам, которыми сопровождаются мутации этих генов, лишающие их функции двоичных переключателей или изменяющие эту функцию. У дрозофилы эти так называемые гомеозисные мутации вызывают трансформации, которые изменяют характер морфогенеза и приводят к замене одной структуры другой, например к возникновению ног вместо антенн или добавочных крыльев вместо жужжалец. Изменение наборов регуляторных генов этого класса или возникновение новых таких наборов создает значительные потенциальные возможности для радикальных эволюционных модификаций или возникновения новых морфологических структур. Подобно изменениям регуляторных генов, влияющих на сроки или структурную интеграцию, изменения регуляторных генов, контролирующих тканевую дифференцировку, также обладают большим эволюционным потенциалом. Если изменения регуляторных генов двух первых типов вызывают изменения формы органов, то изменения генов этого третьего типа приводят к образованию новых тканей. Все это сыграло чрезвычайно важную роль в эволюции размножения млекопитающих и заботы о потомстве. Три способа регуляции развития, которые мы здесь бегло рассмотрели, неотделимы друг от друга. Все они участвовали в морфологической эволюции отдельных групп организмов. Быть может, главная трудность, с которой мы сталкиваемся в нашей попытке понять морфологическую эволюцию в контексте эмбриогенетических механизмов, заключается в том, что формообразование на молекулярном уровне изучено крайне плохо. Дело здесь не только в том, что у нас мало сведений о самих механизмах морфогенеза (перемещения клеток, их взаимодействия, возникновение структурной организации), но и в различных концептуальных подходах к оценке информации, содержащейся в морфологической структуре, и в оценке генетической информации. В качестве иллюстрации этого различия рассмотрим морфогенез не с точки зрения молекулярной генетики, а воспользуемся подходом Д'Арси Томпсона (D'Arcy Thompson), который в своей книге «О росте и форме» (ее первое издание вышло в 1917 г.) впервые применил математику к проблемам формы. Его цель была проста: «Мы хотим понять, как можно объяснить, по крайней мере в некоторых случаях, форму живых существ и частей живых существ, исходя из физических представлений, и установить, что органических форм, которые противоречили бы физическим и математическим законам, не существует». Томпсон изложил свою точку зрения в книге, которая изучалась несколькими поколениями биологов, познакомившихся с ее помощью с математическими законами, лежащими в основе формы поверхностей раздела между клетками и строения радиолярий или спирально закрученных раковин и бараньих рогов; с тем, почему скелет позвоночных и мосты построены в соответствии с одними и теми же инженерными законами, и как, используя преобразования декартовых координат, можно изображать эволюционные изменения формы таких сложных объектов, как черепа, рыбы и изоподы (равноногие рачки). Томпсон снял покров непроницаемой тайны с биологической формы и очень изящно показал, что сложные биологические объекты подчиняются физическим и математическим правилам, поддающимся проверке. Однако он уделял мало внимания событиям, происходящим на генетическом или молекулярном уровне (вероятно, это было разумно, потому что эти события и сейчас еще не вполне поняты), а вместо этого сосредоточился на действующих на организм физических силах как непосредственных факторах, определяющих его морфологию.
Похожие работы
... продемонстрировать отсутствие рекапитуляции в последовательности молекулярных процессов, то это могло бы свидетельствовать о разобщенности путей морфологической и молекулярной эволюции. Как и в случаях диссоциаций, разобщение биохимических событий от консервативной последовательности морфогенетических процессов открывает реальные возможности для эволюционной модификации онтогенеза. Главная задача ...
... с предыдущими классами животных. 10) Нервная система птиц по сравнению с нервной системой пресмыкающихся значительно усложнилась. Высокое развитие центральной нервной системы обусловлено более сложное поведение птиц. Оно проявляется в различных формах заботы о потомстве (гнездостроение, откладка и насиживание яиц, обогревание птенцов, их кормление), в сезонных перемещениях, в развитии звуковой ...
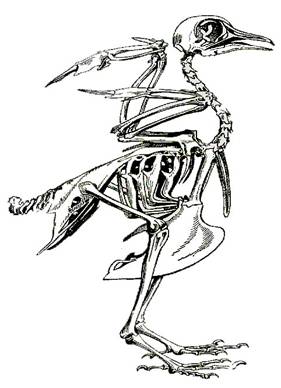
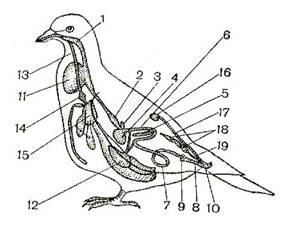
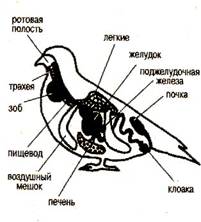
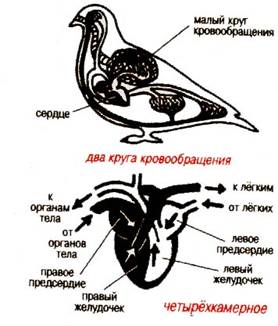
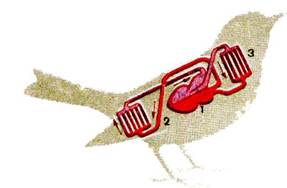
... : после таяния ледника птицы продвинулись на север и освоили экологически новые условия. Пути послеледникового расселения вида часто совпадают с путями пролета. Приспособленность птиц к различным средам обитания Экология птиц В классе птиц выделяют 28 отрядов. Основные из них: пингвины, страусы, киви, гагры, поганки, трубконосые, веслоногие, голенастые, гусеобразные, хищные птицы, ...
... сформированном состоянии, что обеспечивается существенными изменениями зародышевых оболочек, а также полового аппарата и всего материнского организма. Рис. 6. Развитие зародышевых оболочек и плаценты у млекопитающих (/—VI— последовательные стадии): /—внезародышевая вторичная полость, 2—аллантоис, 3 — амнион, 4—желточный пузырь, 5 — зародыш; а — эктодерма, б — энтодерма, в — мезодерма У ...
0 комментариев