Навигация
ГИГАНТСКИЕ РАСТЕНИЯ И ЖИВОТНЫЕ
55705
знаков
0
таблиц
0
изображений
2.4 ГИГАНТСКИЕ РАСТЕНИЯ И ЖИВОТНЫЕ
Как обстоит дело с предположением о том, что в прошлом формы растительной и животной жизни должны были быть гораздо крупнее? Если экранированная Земля существовала, то в летописи окаменелостей должны быть многочисленные примеры гигантских растений и животных. Рассмотрим некоторые из таких примеров. В "Зеленом царстве" - книге, опубликованной Чайлдкрафтом, приводится ряд рисунков растений, живших в прошлом. В основе этих рисунков лежит информация, полученная из летописи окаменелостей. В книге изображены моховидные растения почти метровой длины. В наши же дни их размеры не превышают 5-8 см. Так же в книге показаны растения, похожие на гигантский аспарагус. Длинна их стеблей превышает 12 м. Корневая система этих pастений подобна волосовидным корневым системам современных луковичных растений. Очевидно, что корни этих растении не должны были углубляться далеко в почву, чтобы добывать влагу и не закреплялись в грунте с целью защиты от ветров. В летописи окаменелостей присутствуют хвощи высотой более 15 м. Хвощ растет на болотах и в наши дни, но достигает максимальной высоты от одного до полутора метров, высота папоротниковых растений в прошлом достигала более 16 м, теперешние, же папоротниковые достигают лишь высоты кустарника. В летописи окаменелостей есть также насекомые, гораздо более крупные, чем сегодняшние виды. Например, были найдены тараканы диаметром более 30 см; сохранились стрекозы с размахом крыльев более метра.
Окаменелые останки морских организмов часто гораздо более крупные, чем их сородичи сегодня. В санденском каньоне близ Нанффа (провинция Альберта в Канаде) были найдены окаменелые двустворчатые моллюски длиной более полуметра. В летописи окаменелостей есть раковины наутилоидов диаметром около 2 м. Сегодня их потомки достигают длины не более 20 см.
В разных частях Земли найдены окаменелости гигантских животных различных видов. В книге "Гиганты из прошлого", опубликованной Национальным географическим обществом, показано много таких гигантских, существ, ныне не существующих. Окаменелые останки безрогих носорогов говорят об их росте, превышающем 6 м. Свиньи достигали размеров крупного рогатого скота. Рост верблюда превышал 3,5-метровую отметку. Гигантские птицы вырастали до 3 м, а бобры достигали размеров свиньи. Ширина оленьих рогов превышала 3,5 м. Земляные ленивцы, достигающие ныне размеров средней обезьяны, в летописи окаменелостей представлены экземплярами длиной более 5,5 м.
Одной из наиболее известных особенностей летописи окаменелостей является обилие гигантских рептилий. Наиболее распространенная из них - динозавр. Рептилии вылупляются из яиц, а затем растут. Чем дольше живет рептилия, тем больших размеров она достигает. Динозавр является одним из Крупнейших существ, когда-либо живших на нашей планете. Для того, чтобы динозавр мог вырасти до таких огромных размеров, какие мы видим в летописи окаменелостей, продолжительность его жизни должна быть намного больше продолжительности жизни ныне живущих рептилий. Существование защитного экрана, несомненно, помогает объяснить то, почему животные прошлого достигали больших размеров и жили дольше.
2.5 ОСОБЕННОСТИ ЭВОЛЮЦИИ ЖИВЫХ ОРГАНИЗМОВ НА РАЗНЫХ УРОВНЯХ РАЗВИТИЯ
Особенности эволюции на клеточном уровне
Рассмотрим подробнее особенности эволюции на клеточном уровне организации жизни. Наибольшее различие существует не между растениями, грибами и животными, а между организмами, обладающими ядром (эукариоты) и не имеющими его (прокариоты). Последние представлены низшими организмами - бактериями и сине-зелеными водорослями (цианобактерии, или цианеи), все остальные организмы - эукариоты, которые сходны между собой по внутриклеточной организации, генетике, биохимии и метаболизму.
Различие между прокариотами и эукариотами заключается еще и в том, что первые могут жить как в бескислородной (облигатные анаэробы), так и в среде с разным содержанием кислорода (факультативные анаэробы и аэробы), в то время как для эукариотов, за немногим исключением, обязателен кислород. Все эти различия имели существенное значение для понимания ранних стадий биологической эволюции.
Сравнение прокариот и эукариот по потребности в кислороде приводит к заключению, что прокариоты возникли в период, когда содержание кислорода в среде изменилось. Ко времени же появления у эукариот концентрация кислорода была высокой и относительно постоянной.
Первые фотосинтезирующие организмы появились около 3 млрд. лет назад. Это были анаэробные бактерии, предшественники современных фотосинтезирующих бактерий. Предполагается, что именно они образовали самые древние среди известных строматолитов. Обеднение среды азотистыми органическими соединениями вызывало появление живых существ, способных использовать атмосферный азот. Такими организмами, способными существовать в среде, полностью лишенной органических углеродистых и азотистых соединений, являются фотосинтезирующие азотфиксирующие сине-зеленые водоросли. Эти организмы осуществляли аэробный фотосинтез. Они устойчивы к продуцируемому ими кислороду и могут использовать его для собственного метаболизма. Поскольку сине-зеленые водоросли возникли в период, когда концентрация кислорода в атмосфере колебалась, вполне допустимо, что они - промежуточные организмы между анаэробами и аэробами.
С уверенностью предполагается, что фотосинтез, в котором источником атомов водорода для восстановления углекислого газа является сероводород (такой фотосинтез осуществляют современные зеленые и пурпурные серные бактерии), предшествовал более сложному двустадийному фотосинтезу, при котором атомы водорода извлекаются из молекул воды. Второй тип фотосинтеза характерен для цианеи и зеленых растений.
Фотосинтезирующая деятельность первичных одноклеточных имела три последствия, оказавшие решающее влияние на всю дальнейшую эволюцию живого. Во-первых, фотосинтез освободил организмы от конкуренции за природные запасы абиогенных органических соединений, количество которых в среде значительно сократилось. Развившееся посредством фотосинтеза автотрофное питание и запасание питательных готовых веществ в растительных тканях создали затем условия для появления громадного разнообразия автотрофных и гетеротрофных организмов. Во-вторых, фотосинтез обеспечивал насыщение атмосферы достаточным количеством кислорода для возникновения и развития организмов, энергетический обмен которых основан на процессах дыхания. В-третьих, в результате фотосинтеза в верхней части атмосферы образовался озоновый экран, защищающий земную жизнь от губительного ультрафиолетового излучения космоса.
Еще одно существенное отличие прокариот и эукариот заключается в том, что у вторых центральным механизмом обмена является дыхание, у большинства же прокариот энергетический обмен осуществляется в процессах брожения. Сравнение метаболизма прокариот и эукариот приводит к выводу об эволюционной связи между ними. Вероятно, анаэробное брожение возникло на более ранних стадиях эволюции. После появления в атмосфере достаточного количества свободного кислорода аэробный метаболизм оказался намного выгоднее, так как при окислении углеводов в 8 раз увеличивается выход биологически полезной энергии в сравнении с брожением. Таким образом, к анаэробному метаболизму присоединился аэробный способ извлечения энергии одноклеточными организмами.
Когда же появились эукариотические клетки? На этот вопрос нет точного ответа, но значительное количество данных об ископаемых эукариотах позволяет сказать, что их возраст составляет около 1,5 млрд. лет. Относительно того, каким образом возникли эукариоты, существуют две гипотезы.
Одна из них (аутогенная гипотеза) предполагает, что эукариотическая клетка возникла путем дифференциации исходной прокариотической клетки. Вначале развился мембранный комплекс: образовалась наружная клеточная мембрана с впячиваниями внутрь клетки, из которой сформировались отдельные структуры, давшие начало клеточным органоидам. От какой именно группы прокариот возникли эукариоты, сказать невозможно.
Другую гипотезу (симбиотическую) предложила недавно американский ученый Маргулис. В ее обоснование она положила новые открытия, в частности обнаружение у пластид и митохондрий внеядерной ДНК и способности этих органелл к самостоятельному делению. Л. Маргулис предполагает, что эукариотическая клетка возникла вследствие нескольких актов симбиогенеза. Вначале произошло объединение крупной амебовидной прокариотной клетки с мелкими аэробными бактериями, которые превратились в митохондрии. Затем эта симбиотическая прокариотная клетка включила в себя спирохетоподобные бактерии, из которых сформировались кинетосомы, центросомы и жгутики. После обособления ядра в цитоплазме (признак эукариот) клетка с этим набором органелл оказалась исходной для образования царств грибов и животных. Объединение прокариотной клетки с цианеями привело к образованию пластидной клетки, что дало начало формированию царства растений. Гипотеза Маргулис разделяется не всеми и подвергается критике. Большинство авторов придерживается аутогенной гипотезы, более соответствующей дарвиновским принципам монофилии, дифференциации и усложнения организации в ходе прогрессивной эволюции.
В эволюции одноклеточной организации выделяются промежуточные ступени, связанные с усложнением строения организма, совершенствованием генетического аппарата и способов размножения.
Самая примитивная стадия - агатная прокариотная - представлена цианеями и бактериями. Морфология этих организмов наиболее проста в сравнении с другими одноклеточными (простейшими). Однако уже на этой стадии появляется дифференциация на цитоплазму, ядерные элементы, базальные зерна, цитоплазматическую мембрану. У бактерий известен обмен генетическим материалом посредством конъюгации. Большое разнообразие видов бактерий, способность существовать в самых разных условиях среды свидетельствуют о высокой адаптивности их организации.
Следующая стадия - агамная эукариотпая - характеризуется дальнейшей дифференциацией внутреннего строения с формированием высокоспециализированных органоидов (мембраны, ядро, цитоплазма, рибосомы, митохондрии и др.). Особо существенной здесь была эволюция ядерного аппарата - образование настоящих хромосом в сравнении с прокариотами, у которых наследственное вещество диффузно распределено по всей клетке. Эта стадия характерна для простейших, прогрессивная эволюция которых шла по пути увеличения числа одинаковых органоидов (полимеризация), увеличения числа хромосом в ядре (полиплоидизация), появления генеративных и вегетативных ядер - макронуклеуса и микронуклеуса (ядерный дуализм). Среди одноклеточных эукариотных организмов имеется много видов с агамным размножением (голые амебы, раковинные корненожки, жгутиконосцы).
Прогрессивным явлением в филогенезе простейших было возникновение у них полового размножения (гамогонии), которое отличается от обычной конъюгации. У простейших имеется мейоз с двумя делениями и кроссинговером на уровне хроматид, и образуются гаметы с гаплоидным набором хромосом. У некоторых жгутиковых гаметы почти неотличимы от бесполых особей и нет еще разделения на мужские и женские гаметы, т.е. наблюдается изогамия. Постепенно в ходе прогрессивной эволюции происходит переход от изогамии к анизогамии, или разделению генеративных клеток на женские и мужские, и к анизогамной копуляции. При слиянии гамет образуется диплоидная зигота. Следовательно, у простейших наметился переход от агамной эукариотной стадии к зиготной - начальной стадии ксеногамии (размножение путем перекрестного оплодотворения). Последующее развитие уже многоклеточных организмов шло по пути совершенствования способов ксеногамного размножения.
Возникновение и развитие многоклеточной организации
Следующая после возникновения одноклеточных ступень эволюции заключалась в образовании и прогрессивном развитии многоклеточного организма. Эта ступень отличается большой усложненностью переходных стадий, из которых выделяются колониальная одноклеточная, первично - дифференцированная, централизованно - дифференцированная.
Колониальная одноклеточная стадия считается переходной от одноклеточного организма к многоклеточному и является наиболее простой из всех стадий в эволюции многоклеточной организации.
Недавно обнаружены самые примитивные формы колониальных одноклеточных, стоящих как бы на полпути между одноклеточными организмами и низшими многоклеточными (губками и кишечнополостными). Их выделили в подцарство Mesozoa, однако в эволюции на многоклеточную организацию представителей этого полцарства считают тупиковыми линиями.
Первично-дифференцированная стадия в эволюции многоклеточной организации характеризуется началом специализации по принцип "разделения труда" у членов колонии. Элементы первичной специализации наблюдаются у колоний Pandorina morum (16 клеток), Eudorina elegans (32 клетки), Volvox globator (тысячи клеток). Специализация у названных организмов сводится к разделению клеток на соматические, осуществляющие функции питания и движения (жгутики), и генеративные (гонидии), служащие для размножения. Здесь наблюдается и выраженная анизогамия. На первично-дифференцированной стадии происходит специализация функций на тканевом, органном и системно-органном уровне. Так, у кишечнополостных уже сформировалась простая нервная система, которая, распространяя импульсы, координирует деятельность двигательных, железистых, стрекательных, репродуктивных клеток. Нервного центра как такового еще нет, но центр координации имеется.
С кишечнополостных начинается развитие централизованно-дифференцированной стадии в эволюции многоклеточной организации. На этой стадии усложнение морфофизиологической структуры идет через усиление тканевой специализации, начиная с возникновения зародышевых листков, детерминирующих морфогенез пищевом, выделительной, генеративной и других систем органов. Возникает хорошо выраженная централизованная нервная система: у беспозвоночных - ганглиолярная, у позвоночных - с центральным и периферическим отделами. Одновременно совершенствуются способы полового размножения от наружного оплодотворения к внутреннему, от свободной инкубации яиц вне материнского организма к живорождению.
Финалом в эволюции многоклеточной организации животных было появление организмов с поведением "разумного типа". Сюда относятся животные с высокоразвитой условно-рефлекторной деятельностью, способные передавать информацию следующему поколению не только через наследственность, но и надгаметным способом (например, передача опыта молодняку посредством обучения). Заключительным этапом в эволюции централизованно-дифференцированной стадии стало возникновение человека.
Рассмотрим основные этапы эволюции многоклеточных организмов в той последовательности, как она происходила в геологической истории Земли. Всех многоклеточных разделяют на три царства: грибы (Fungi), растения (Metaphyta) и животные (Metazoa). Относительно эволюции грибов известно очень мало, так как палеонтологицеская летопись их остается скудной. Два других царства намного богаче представлены ископаемыми остатками, дающими возможность довольно подробно восстановить ход их истории.
Эволюция животного мира
История животных изучена наиболее полно в связи с тем, что они обладают скелетом и поэтому лучше закрепляются в окаменелых остатках. Самые ранние следы животных обнаруживаются в конце докембрия (700 млн. лет). Предполагается, что первые животные произошли либо от общего ствола всех эукариот, либо от одной из групп древнейших водорослей. Наиболее близки к предкам простейших животных (Protozoa) одноклеточные зеленые водоросли. Неслучайно, например, эвглену и вольвокс, способных и к фотосинтезу, и к гетеротрофному питанию, ботаники относят к типу зеленых водорослей, а зоологи - к типу простейших животных. За всю историю животного мира возникло 35 типов, из которых 9 вымерло, а 26 существуют до сих пор.
Разнообразие и количество палеонтологических документов в истории животных резко возрастают в породах, датируемых менее 570 млн. лет. В течение примерно 50 млн. лет довольно быстро появляются почти все типы вторичнополостных животных с прочным скелетом. Широко были распространены в морях силура трилобиты. Возникновение типа хордовых (Chordata) относится ко времени менее 500 млн. лет. Комплексы хорошо сохранившихся ископаемых найдены в сланцах Бергеса (Колумбия), содержащих остатки беспозвоночных, в частности, мягкотелых организмов типа Annelida, к которому принадлежат современные дождевые черви.
Начало палеозоя отмечено образованием многих типов животных, из которых примерно треть существует в настоящее время. Причины такой активной эволюции остаются неясными. В позднекембрийское время появляются первые рыбы, представленные бесчелюстными - Agnata. В дальнейшем они почти все вымерли, из современных потомков сохранились миноги. В девоне возникают челюстные рыбы в результате таких крупных эволюционных преобразований, как превращение передней пары жаберных дуг в челюсти и формирование парных плавников. Первых челюстноротых представляли две группы: лучеперые и лопастеперые. Почти все ныне живущие рыбы - потомки лучеперых. Лопастеперые представлены сейчас только двоякодышащими и небольшим числом реликтовых морских форм. Лопастеперые имели в плавниках костные опорные элементы, из которых развились конечности первых обитателей суши. Ранее из группы лопастеперых возникли амфибии, следовательно, все четвероногие позвоночные имеют своим далеким предком эту исчезнувшую группу рыб.
Наиболее древние представители амфибий - ихтиостеги - обнаружены в верхнедевонских отложениях (Гренландия). Эти животные обладали пятипалыми конечностями, с помощью которых они могли переползать по суше. Все же ряд признаков (настоящий хвостовой плавник, покрытое мелкими чешуйками тело) свидетельствует о том, что ихтиостеги обитали преимущественно в водоемах. Конкуренция с кистеперыми рыбами заставляла этих первых земноводных занимать промежуточные между водой и сушей местообитания.
Расцвет древних амфибий приурочен к карбону, где они были представлены большим разнообразием форм, объединяемых под названием "стегоцефалы". Среди них наиболее выделяются лабиринтодонты и крокодилообразные. Два отряда современных амфибий - хвостатые и безногие (или червяги) - произошли, вероятно, от других ветвей стегоцефалов.
От примитивных амфибий ведут свое начало рептилии, широко расселившиеся на суше к концу пермского периода благодаря приобретению легочного дыхания и оболочек яиц, защищающих от высыхания. Среди первых рептилий особенно выделяются котилозавры - небольшие насекомоядные животные и активные хищники - терапсиды, уступившие в триасе место гигантским рептилиям, динозаврам, появившимся 150 млн. лет назад. Вполне вероятно, что последние были теплокровными животными. В связи с теплокровностью динозавры вели активный образ жизни, чем можно объяснить их длительное господство и сосуществование с млекопитающими. Причины вымирания динозавров (примерно 65 млн. лет назад) неизвестны. Предполагают, в частности, что это могло быть следствием массового уничтожения яиц динозавров примитивными млекопитающими. Более правдоподобной кажется гипотеза, согласно которой вымирание динозавров связано с резкими колебаниями климата и уменьшением растительной пищи в меловом периоде. Уже в период господства динозавров существовала предковая группа млекопитающих - небольших по размеру с шерстным покровом животных, возникших от одной из линий хищных терапсид. Млекопитающие выходят на передний край эволюции благодаря таким прогрессивным адаптациям, как плацента, вскармливание потомства молоком, более развитый мозг и связанная с этим большая активность, теплокровность. Значительного разнообразия млекопитающие достигли в кайнозое, появились приматы. Третичный период был временем расцвета млекопитающих, но многие из них вскоре вымерли (например, ирландский олень, саблезубый тигр, пещерный медведь).
Прогрессивная эволюция приматов оказалась уникальным явлением в истории жизни, в итоге она привела к возникновению человека.
2.6 ПРОДОЛЖИТЕЛЬНОСТЬ ЖИЗНИ ЧЕЛОВЕКА
Если "экранная" модель правильна, то должны существовать и свидетельства того, что жизнь человека в прошлом была намного более продолжительной. Как можно видеть из описаний генеалогии человека, жившего на первобытной Земле, средняя продолжительность его жизни была намного больше, чем у человека наших дней.
С течением времени коротковолновое излучение продолжало вызывать дегенеративные изменения в генетических программах живых существ, переходившие к последующим поколениям. Процесс старения ускорялся. В наши дни средняя продолжительность жизни человека составляет приблизительно от 70 до 80 лет и никогда даже не приближается к 900 годам жизни людей (по предположению), защищенную водным экраном, помогает понять причины долголетия первобытного человека, и отвечает на многие вопросы, возникающие при изучении летописи окаменелостей.
ЗАКЛЮЧЕНИЕ
В данной работе были описаны теории как о возникновении и эволюции планеты, так и жизни на ней. Некоторые из этих теорий и предположений не доказаны, некоторые, как считают учёные, необоснованные. И все же, как ни была бы сомнительна любая из теорий о развитии нашей планеты и жизни на ней, каждая теория имеет право на существование, раз имеет сторонников. Но человечество не остановится на этом - оно будет искать единственно правильную теорию, даже если нужно будет разрушить то, что уже создано на протяжении многих веков.
СПИСОК ИСПОЛЬЗОВАННОЙ ЛИТЕРАТУРЫ
1. Куликовский П. Г., Справочник любителя АСТРОНОМИИ // М. 1971 г. 2. Воронцов-Вельяминов Б.А., Очерки о Вселенной // М. "Наука" 1976 г. 3. Новиков И.Д., Эволюция Вселенной // М. 1983 г. 4. Левитан С.П., Астрономия // М. "Просвещение" 1994 г.
ИНТЕРНЕТ – РЕСУРСЫ
2. http://studzona.co.ua/referats/view/222
3. http://galspace.spb.ru/phpBB2/viewtopic.php?t=444&view=previous
Похожие работы
... В.Г. Рождение звезд. М.: УРСС, 1997. 207 с. 7. Шаткин Г.А. Наша планета – Земля // Наука и жизнь. -1999. -№ 5. Технология семинара На тему «Земля как планета солнечной системы. Проблемы целостного освоения Земли» нами был проведен семинар. В ходе семинара использовался теоретический материал представленный в курсовой работе, а перед его началом каждому студенту был предоставлен буклет ...
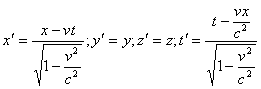
... инерциальных системах отсчета. Пространственно-временной континуум – неразрывная связь пространства и времени и их зависимость от системы отсчета. Тема 11. Основные концепции химии 1. Химия как наука, ее предмет и проблемы Важнейшим разделом современного естествознания является химия. Она играет большую роль в решении наиболее актуальных и перспективных проблем современного общества. К ...
... мы учтем эти факторы, тем точнее сможем предсказать развитие той или иной потребности и обусловленного ею поведения потребителей. 1.6. Концепция гуманистического психоанализа. Глубокая концепция человека и его положения в современном западном мире была разработана во второй половине XX в. выдающимся психоаналитиком, социологом и философом, создателем гуманистического психоанализа Эрихом ...
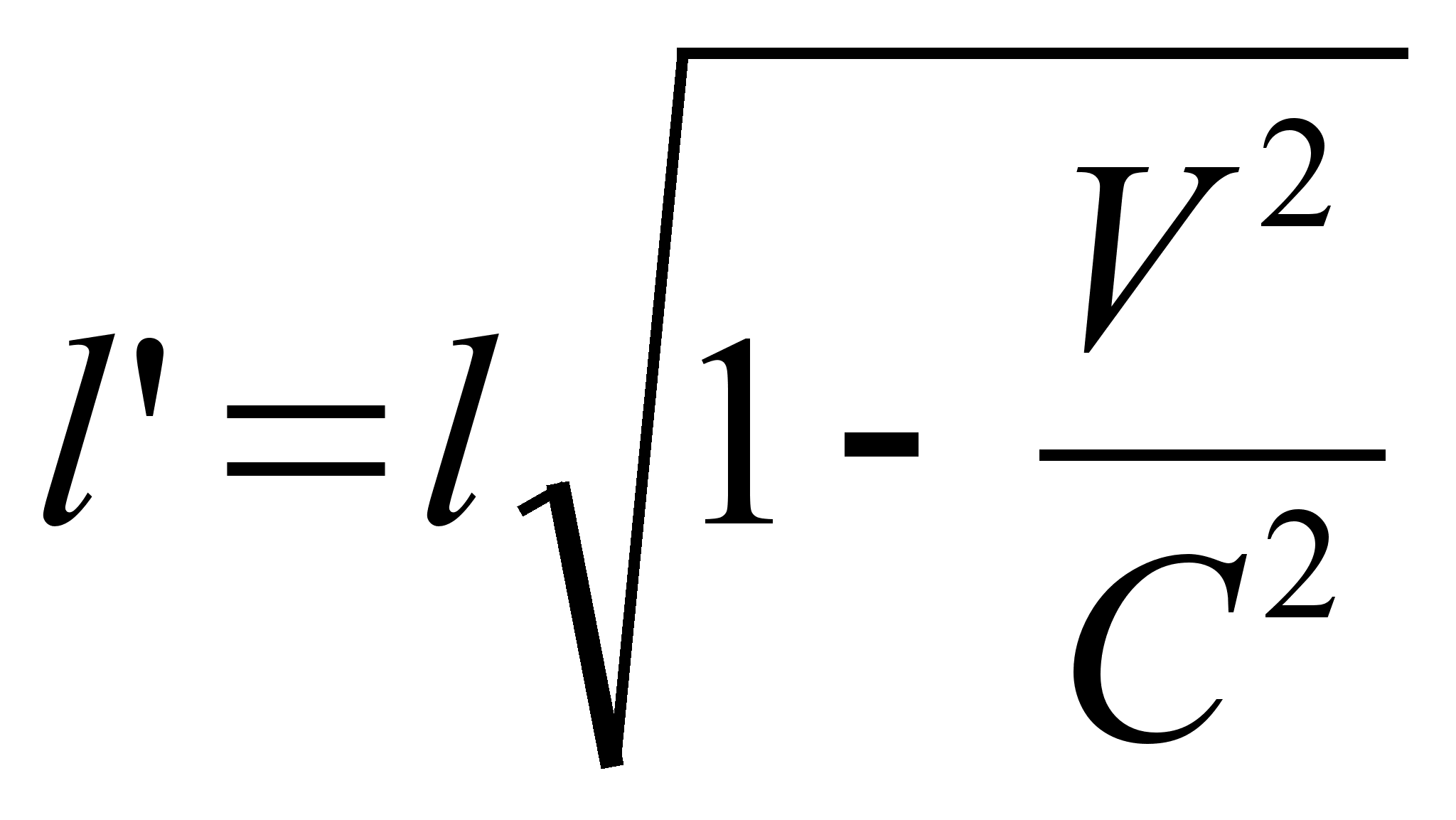
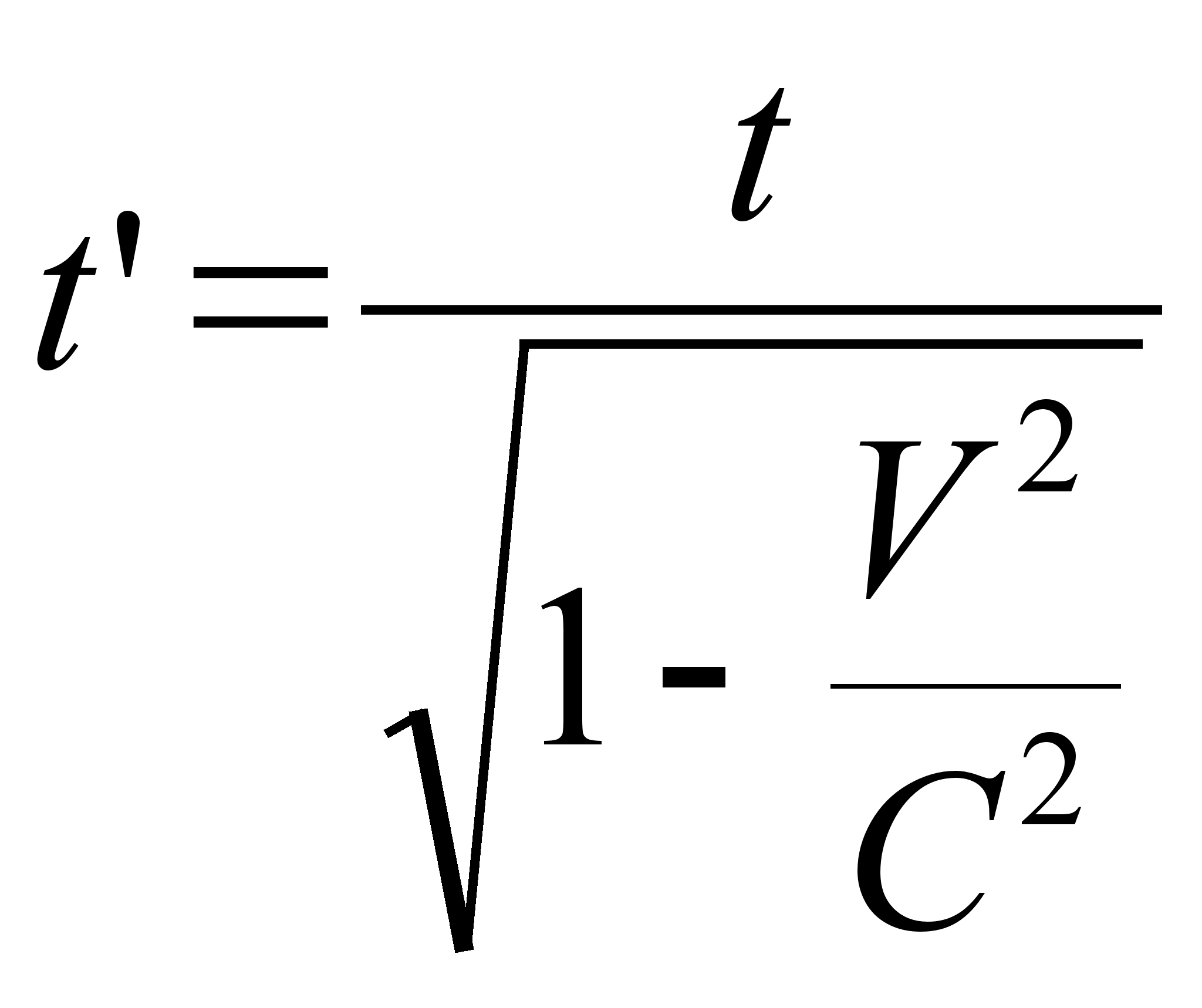
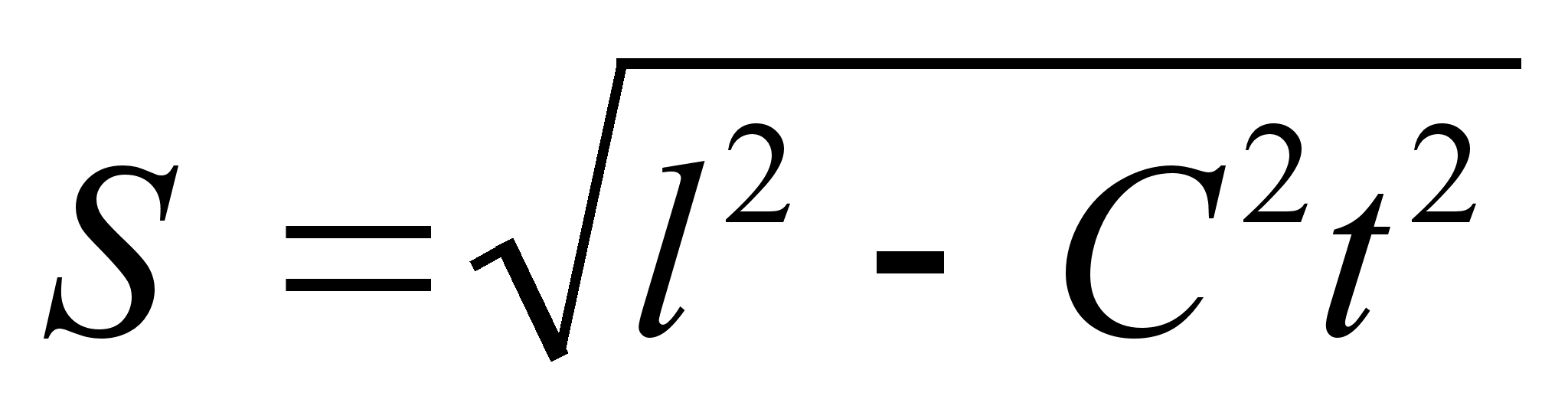
... ноосфере — сфере взаимодействия природы и общества, в пределах которой разумная человеческая деятельность становится определяющим фактором эволюции. Большой вклад в такое понимание естественно-научной картины мира и места человека в истории Земли внес П. Тейяр де Шардев. Согласно ему, в ходе развития Вселенной на Земле естественным "скачкообразным образом" совершился переход от неживого ...
0 комментариев