Навигация
Загальна характеристика вірусів
55889
знаков
0
таблиц
1
изображение
2. Загальна характеристика вірусів
Основними формами життя на Землі є організми клітинної будови. Цей тип організації характерний для всіх видів живих істот, за винятком вірусів, які розглядають як неклітинні форми життя.
Віруси настільки малі, що лише в кілька разів перевищують розміри великих молекул білків. Віруси мають розміри 10—275 нм. їх можна побачити лише під електронним мікроскопом. Вони легко проходять крізь пори спеціальних фільтрів, що затримують усі бактерії і клітини багатоклітинних організмів.
Віруси були відкриті у 1892 р. російським фізіологом рослин і мікробіологом Д. І. Івановським під час вивчення хвороби тютюну.
Віруси — збудники багатьох хвороб рослин і тварин. Вірусними хворобами людини є кір, грип, гепатит А (хвороба Боткіна), поліомієліт (дитячий параліч), сказ, віспа тощо.
Під електронним мікроскопом різні види вірусів мають форму паличок або кульок. Окрема вірусна часточка складається з молекули нуклеїнової кислоти (ДНК або РНК), скрученої в клубок, і молекул білка, розміщених у вигляді своєрідної оболонки навколо молекули кислоти (капсид).
Віруси не здатні самостійно синтезувати нуклеїнові кислоти і білки, з яких вони складаються. Розмноження вірусів можливе лише в разі використання ферментних систем клітин. Потрапивши у клітину, віруси змінюють і перебудовують обмін речовин у ній, внаслідок чого клітина починає синтезувати молекули нових вірусних часточок. Поза клітинами віруси переходять у кристалічний стан, що сприяє їх збереженню.
У житті вірусів можна виділити такі етапи: прикріплення вірусу до клітини, вторгнення вірусу в клітину, латентну стадію, утворення нового покоління вірусів, вихід віріонів. У латентну стадію вірус ніби зникає. Його не вдається виявити або виділити з клітини, але в цей період уся клітина синтезує необхідні для вірусу білки і нуклеїнові кислоти, в результаті чого утворюється нове покоління віріонів.
Проникнення вірусу в клітину організму хазяїна розпочинається із взаємодії вірусної часточки з поверхнею клітини, на якій є особливі рецепторні ділянки. Оболонка часточки вірусу має відповідні прикріпні білки, які "впізнають" ці ділянки.
Саме цим зумовлена висока специфічність вірусів стосовно клітинхазяїв: часто віруси уражують лише певний тип клітин якогось виду організмів.
Так, вірус поліомієліту уражує лише нервові клітини людини, а вірус тютюнової мозаїки — клітини листків тютюну.
Якщо часточка вірусу прикріплюється не до рецепторних ділянок, а до інших місць на поверхні клітинихазяїна, то зараження останньої може і не відбутися. Отже, наявність рецепторних ділянок на поверхні клітини визначає її чутливість до того чи іншого виду вірусів.
Усередину клітинихазяїна вірус може проникнути різними шляхами. Часом оболонки вірусних часточок зливаються з клітинною мембраною (як у вірусу грипу), і ДНК виявляється у цитоплазмі клітини, іноді вірусна часточка потрапляє в клітину шляхом піноцитозу, після чого ферменти клітинихазяїна розщеплюють її оболонку і вивільняють нуклеїнову кислоту (вірус поліомієліту тварин).
У рослинні клітини віруси можуть проникати крізь пошкоджені ділянки клітинної стінки.
У 1917 р. французький вчений Ф. д'Ерелл відкрив віруси бактерій — бактеріофаги (або фаги). Під електронним мікроскопом вони мають форму коми або тенісної ракетки розміром близько 5 нм.
Коли часточка фага прикріплюється своїм тонким відростком до бактеріальної клітини, його ДНК проникає в клітину і викликає синтез нових молекул ДНК і білка бактеріофага. Через 30—60 хв. бактеріальна клітина руйнується і з неї виходять сотні нових часточок фага, здатних спричинити зараження інших бактеріальних клітин.
Спочатку вважали, що бактеріофаги можна використовувати для боротьби з хвороботворними бактеріями. Проте згодом виявилося, що фаги швидко руйнують бактерії в пробірці, але неефективні в живому організмі. У зв'язку з цим їх використовують в основному для діагностики захворювань, виявлення бактерій. [1,4]
3. Механізми імунологічної захисту організму
Початок розвитку імунології відноситься до кінця XVIII століття і пов'язане з ім'ям Е. Дженнера, вперше застосував на підставі лише практичних спостережень згодом обгрунтований теоретично метод вакцинації проти натуральної віспи.
Відкритий Е. Дженнером факт ліг в основу подальших експериментів Л. Пастера, що завершилися формулюванням принципу профілактики від інфекційних захворювань - принцип імунізації ослабленими або вбитими збудниками.
Розвиток імунології довгий час відбувалося в рамках мікробіологічної науки і стосувалося лише вивчення несприйнятливості організму до інфекційних агентів. На цьому шляху були досягнуті великі успіхи у розкритті етіології ряду інфекційних захворювань. Практичним досягненням стала розробка методів діагностики, профілактики та лікування інфекційних захворювань в основному шляхом створення різного роду вакцин і сироваток. Численні спроби з'ясування механізмів, що обумовлюють стійкість організму проти збудника, увінчалися створенням двох теорій імунітету - фагоцитарної, сформульованої в 1887 році І. І. Мечниковим, і гуморальної, висунутої в 1901 році П. Ерліхом.
Початок XX століття - час виникнення іншої гілки імунологічної науки - імунології неінфекційної. Як відправною точкою для розвитку інфекційної імунології з'явилися спостереження Е. Дженнера, так для неінфекційної - виявлення Ж. Борде і Н. Чистовича факту вироблення антитіл в організмі тварини у відповідь на введення не тільки мікроорганізмів, а взагалі чужорідних агентів. Своє твердження і розвиток неінфекційна імунологія отримала у створеному І. І. Мечниковим в 1900 р. вченні про цитотоксину - антитіла проти певних тканин організму, у відкритті К. Ландштейнером в 1901 році антигенів людських еритроцитів.
Результати робіт П. Медавара розширили рамки і привернули пильну увагу до неінфекційної імунології, пояснивши, що в основі процесу відторгнення чужорідних тканин організмом лежать теж імунологічні механізми. І саме подальше розширення досліджень в області трансплантаційного імунітету привернуло до відкриття в 1953 році явища імунологічної толерантності - неотвечаемості організму на введену чужорідну тканину.
Таким чином, навіть короткий екскурс в історію розвитку імунології дозволяє оцінити роль цієї науки у вирішенні ряду медичних і біологічних проблем. Інфекційна імунологія - прародителька загальної імунології - стала в даний час тільки її гілкою.
Стало очевидним, що організм дуже точно розрізняє "своє" і "чуже", а в основі реакцій, що виникають у ньому у відповідь на введення чужорідних агентів (незалежно від їх природи), лежать одні і ті ж механізми. Вивчення сукупності процесів і механізмів, спрямованих на збереження сталості внутрішнього середовища організму від інфекцій та інших чужорідних агентів - імунітету, лежить в основі імунологічної науки.
Друга половина ХХ століття ознаменувалася бурхливим розвитком імунології. Саме в ці роки була створена селекційно-клональна теорія імунітету, розкриті закономірності функціонування різних ланок лімфоїдної системи як єдиної і цілісної системи імунітету. Одним з найважливіших досягнень останніх років стало відкриття двох незалежних ефекторних механізмів у специфічному імунній відповіді. Один з них пов'язаний з так званими В-лімфоцитами, що здійснюють гуморальну відповідь (синтез імуноглобулінів), інший - з системою Т-лімфоцитів (тимус-залежних клітин), наслідком діяльності яких є клітинна відповідь (накопичення сенсибілізованих лімфоцитів). Особливо важливим є отримання доказів існування взаємодії цих двох видів лімфоцитів в імунній відповіді.
Результати досліджень дозволяють стверджувати, що імунологічна система - важлива ланка в складному механізмі адаптації людського організму, а його дію в першу чергу направлено на збереження антигенного гомеостазу, порушення якого може бути обумовлено проникнення в організм чужорідних антигенів (інфекція, трансплантація) або спонтанної мутації. [2]
Але, як показали дослідження останніх років, поділ імунітету на гуморальний і клітинний вельми умовний. Насправді, вплив антигену на лімфоцит і ретикулярну клітину здійснюється за допомогою мікро- та макрофагів, переробних імунологічну інформацію. У той же час реакція фагоцитозу, як правило, беруть участь гуморальні фактори, а основу гуморального імунітету складають клітини, що продукують специфічні імуноглобуліни. Механізми, спрямовані на елімінацію чужорідного агента, надзвичайно різноманітні. При цьому можна виділити два поняття ‑ "імунологічна реактивність" і "неспецифічні фактори захисту". Під першим розуміються специфічні реакції на антигени, зумовлені високо специфічною здатністю організму реагувати на чужорідні молекули. Однак захищеність організму від інфекцій залежить ще і від ступеня проникності для патогенних мікроорганізмів шкірних і слизових покривів, та наявності в їх секрети бактерицидних субстанцій, кислотності шлункового вмісту, присутності в біологічних рідинах організму таких ферментних систем, як лізоцим. Всі ці механізми відносяться до неспецифічних чинників захисту, так як немає ніякого спеціального реагування і всі вони існують незалежно від присутності або відсутності збудника. Деяке особливе становище займають фагоцити і система комплементу. Це обумовлено тим, що, незважаючи на неспецифічність фагоцитозу, макрофаги беруть участь у переробці антигену і в кооперації Т- і В-лімфоцитів при імунній відповіді, тобто беруть участь у специфічних формах реагування на чужорідні субстанції. Аналогічно вироблення комплементу не є специфічною реакцією на антиген, але сама система комплементу бере участь у специфічних реакціях антиген-антитіл.
За клітинний імунітет відповідальні Т - лімфоцити (Т - клітини). Ці імунні клітини названі так через тимус, в якому вони піддаються основних стадіях своєї диференціації (школа Т-клітин). Активність Т-клітин спрямована проти зараженої вірусом клітини організму, а також на захист від грибів і паразитів. Т-клітини беруть активну участь в процесі відторгнення чужорідної тканини і допомагають у формуванні гуморальної імунної відповіді (див. нижче). За своєю функцією вони діляться на цитотоксичні Т-клітини ‑ Т-кілери (на схемі зеленого кольору) і клітини-помічники ‑ Т-хелпери (на схемі блакитного кольору).
У свою чергу гуморальну імунну відповідь спрямований на активацію В-лімфоцитів, які дозрівають у кістковому мозку на відміну від Т-клітин тимуса. Т-Клітини несуть на своїй поверхні антитіла і виділяють їх в плазму. Антитіла мають здатність специфічно зв'язувати відповідні антигени. Зв'язування антитіл з антигенами - вирішальне ланка в системі захисту організму від позаклітинних вірусів і бактерій. У результаті такого зв'язування останні орієнтуються як чужорідні тіла і надалі знищуються.[6]
"Пам'ять" імунної системи представлена так званими "клітинами пам'яті". Ці найбільш довгоживучі клітини існують для кожного типу імунних клітин.
На рис. 1 представлена спрощена імунна відповідь організму на антигени. Вірус, що проник в організм ендоцендоцитується макрофагами і потім частково руйнується в ЕПР (1). У результаті утворюються чужорідні фрагменти, які експонуються на клітинній поверхні макрофагів (2). Ці фрагменти «презентуються» спеціальною групою мембранних білків. Комплекс з вірусного фрагмента і білка головного комплексу гістосумісності розпізнається і зв'язується Т-клітинами за допомогою специфічних (Т-клітинних) рецепторів. Серед величезного числа Т-клітин тільки одиниці володіють відповідним рецептором (3), Зв'язування призводить до активації цих Т-клітин і появи їх селективних копій (4, "клональна селекція"). У активації Т-клітин беруть участь різні гормоноподібні сигнальні білки, інтерлейкіни. Ці білки секретуються тими клітинами імунної системи, які активуються при зв'язуванні з Т-клітинами. Так, активовані макрофаги з презентованим вірусним фрагментом секретують IL-1 (5), а Т-клітини продукують IL-2 (6), який стимулює їх власне клональне копіювання і реплікацію Т-хелперних клітин.
Клоновані та активовані Т-клітини здійснюють різні функції залежно від їх типу. Цитотоксичні Т-клітини здатні впізнавати і пов'язувати ті клітини організму, які інфіковані вірусами і на своїх рецепторах ГКГС несуть фрагменти вірусу (7). Цитотоксичні Т-клітини секретують перфорин - білок, який робить проникною мембрану пов'язаної інфікованої клітини, що і призводить до її лізису (8).
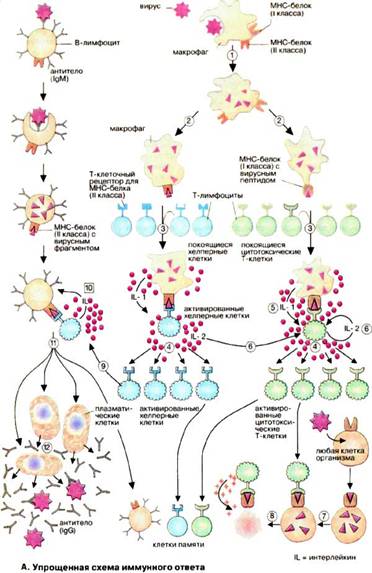
Рис. 1 Спрощена схема імунної відповіді
Т-хелпери, навпаки, зв'язуються з В-клітинами, які презентують на своїй поверхні фрагменти вірусу, пов'язані з білком ГКГС (9). Це веде до селективного клонування індивідуальних В-клітин і їх масованої проліферації. Інтерлейкін стимулює (10) дозрівання В-клітин ‑ перетворення в плазматичні клітини (11), здатні синтезувати і секретувати антитіла (12).[5]
Похожие работы

... межах господарства, заготівлею та реалізацією тваринницької продукції. Ферми і приватні господарства комплектують лише здоровими, перевіреними на туберкульоз тваринами, з благополучних щодо інфекційних захворювань господарств. У благополучних щодо туберкульозу господарствах потрібно забезпечувати нормативні умови утримання, годівлі та використання сільськогосподарських тварин, а також ізольоване ...
... епідеміологічне та клінічне обстеження хворих, статистична обробка даних, аналіз та узагальнення результатів дослідження, формулювання висновків). АНОТАЦІЯ Шкондіна О.Ф. Особливості перебігу хронічного гепатиту С в залежності від генотипу вірусу. – Рукопис. Дисертація на здобуття наукового ступеня кандидата медичних наук за спеціальністю 14.01.13 – інфекційні хвороби. ДУ ''Інститут епі ...
0 комментариев