Навигация
Перенос генетического материала с помощью рекомбинации
16948
знаков
0
таблиц
0
изображений
1.2 Перенос генетического материала с помощью рекомбинации
Явление рекомбинации у актиномицет напоминает гибридизацию у высших организмов. Установлено, что при контакте клеток (чаще дефектных) двух разных штаммов бактерий или актиномицетов свойства одного штамма переходят к другому. В результате получаются смешанные формы с признаками двух исходных культур. Такой процесс происходит между двумя родственными организмами.
Явление генетической рекомбинации было продемонстрировано многими исследователями у ряда представителей рода Streptomyces, в том числе продуцирующих антибиотики, но только у одного из штаммов Str. coelicolor генетические исследования проводились планомерно в течение многих лет . Однако, хотя основные сведения о генетической системе актиномицетов получены для Str. coelicolor, результаты исследований других видов актиномицетов согласуются с ними, что дает возможность говорить об общих особенностях генетической рекомбинации в пределах рода Streptomyces (Actinomyces, по классификации Н. А. Красильникова).
Методические подходы при изучении генетики актиномицетов принципиально те же, что и для других микроорганизмов. Скрещивания производят между штаммами, маркированными различными генетическими факторами (биохимическая недостаточность, устойчивость к антибиотикам и др.), а отбор рекомбинантов ведут на специально подобранных селективных средах. И то и другое необходимо, так как генетическая рекомбинация — явление редкое и генетические рекомбинанты составляют лишь незначительную долю в популяции исходных штаммов.
В настоящее время можно считать установленным, что процесс генетической рекомбинации у актиномицетов в основном сходен с процессом конъюгации у бактерий, детально изученным у Е. coli, Str. coelicolor, подобно бактериям, имеет единую кольцевую группу сцепления, на которой определено местоположение около 40 различных генетических локусов. Характерная особенность генетической карты Str. coelicolor состоит в неслучайном расположении генетических локусов, сосредоточенных преимущественно в двух областях карты, тогда как две другие области являются почти «пустыми». Такое разобщение двух групп локусов в пространстве (возможно, лишь кажущееся) и послужило причиной первоначального представления о наличии у актиномицетов двух независимых групп сцепления.
Явление генетической рекомбинации описано у большинства изученных видов актиномицетов. Однако возникновение генетических рекомбинантов при внутривидовых скрещиваниях наблюдается далеко не во всех комбинациях мутантов, даже если они и происходят из одного и того же штамма. Вопрос о половой полярности штаммов внутри одного вида до сих пор остается не решенным, хотя и имеются некоторые данные в пользу ее существования. Лучше изучен вопрос об особенностях самого процесса генетической рекомбинации у актиномицетов. Этот процесс состоит из нескольких этапов, причем, как установлено недавними исследованиями, оба родительских штамма принимают неодинаковое участие в скрещивании: один — играет роль донора, другой — реципиента генетического материала, напоминая в этом отношении бактерии.
Как и у бактерий, в результате переноса генетического материала от одного штамма к другому происходит образование неполных зигот (мерозигот), содержащих полный геном реципиентного штамма и часть генома донорного. При этом диплоидный участок мерозиготы может варьировать как по составу, так и по протяженности, а процесс возникновения частичного диплоидного ядра происходит во времени. В отличие от бактерий, для актиномицетов характерно длительное существование стадии мерозиготы, сохраняющейся в течение ряда поколений, постепенно сменяющейся стадией образования гаплоидных рекомбинантов. В соответствии с этим у актиномицетов описаны клоны, являющиеся по своей генетической структуре мерозиготами. Они характеризуются нестабильностью и в процессе размножения выщепляют различные клоны гаплоидных рекомбинантов. Открытие таких клонов, названных гетероклонами, дало возможность разработать простой метод генетического анализа у актиномицетов, основанный на учете различных типов гаплоидных рекомбинантов в потомстве гетероклонов.
Таким образом, процесс генетической рекомбинации у актиномицетов состоит из нескольких этапов, последний из которых заключается в образовании гаплоидных рекомбинантов. Эти рекомбинанты, в отличие от гетероклонов, являются стабильными и служат основным объектом исследований в промышленных скрещиваниях. Однако необходимо иметь в виду, что вследствие неодинакового участия двух родительских штаммов в скрещивании, когда один из них поставляет только часть своего генетического материала другому, гаплоидные рекомбинанты наследуют большинство генетических факторов от одного родителя и только некоторые — от другого. Иными словами, по своей генетической структуре гаплоидные рекомбинанты, как правило, более напоминают одного родителя, чем другого, что неизбежно ограничивает возможности гибридизации у актиномицетов.
Наряду с генетической рекомбинацией у актиномицетов наблюдается другое широко распространенное явление — гетерокариозис, во многом сходное с аналогичным явлением у грибов.
Первоначально считали, что возникновение гетерокарионов между двумя штаммами представляет собой первый необходимый этап генетической рекомбинации. Однако в настоящее время имеются данные, что оба эти явления не связаны между собой причинно и происходят независимо друг от друга. Поскольку гетерокариотические клоны являются обычно нестабильными, расщеплясь при размножении на оба исходных родительских типа, гетерокариозис не может использоваться в качестве способа получения гибридных форм у актиномицетов.
Похожие работы
... . Генно-инженерные технологии позволяют легко увеличивать промышленное производство ферментов. Ферменты находят все более широкое применение как биокатализаторы в фармацевтическом производстве. Биокаталитические технологии. Направленная модификация с помощью методов генной инженерии открывает возможности трансформации структуры ферментов таким образом, что они приобретают качественно новые ...
... . ЛИТЕРАТУРА: 1. Курс низших растений //Под ред. М. В. Горленко – М.: Высш. школа, 1981. 2. Жизнь растений, т. 1–3, - М.: Просвещение, 1974-77. 3. Н. Грин, У. Стаут, Д. Тейлор. Биология, т. 1-3. – М.: Мир, 1990. 4. Н.П.Горбунова , Е.С.Клюшникова, Н.А.Комарницкий. и др. Малый практикум по низшим растениям. – М.: Высш. школа, 1976. 5. Русанов В. А. Низшие растения. Методические указания ...
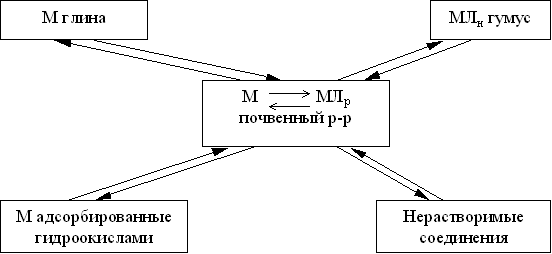
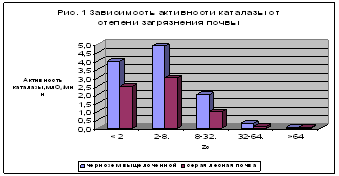
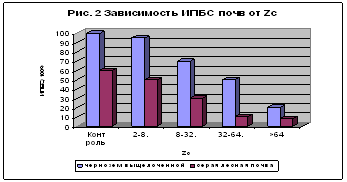
... в гумосфере Свинец 207,5 24,0 8,6 Мышьяк 739,0 12,0 61,6 Кадмий 7,4 1,2 6,2 Уран 590,4 2,4 246,0 Ртуть 0,55 0,024 27,1 Олово 295,7 19,0 15,6 Серебро 3,0 0,24 12,5 2.2 Характеристика приоритетных веществ – загрязнителей почвы Всевозрастающее внимание к охране окружающей среды вызвал особый интерес к вопросам воздействия на почву тяжелых металлов, которые в первую ...
0 комментариев