Навигация
Генетический анализ бактериофагов
23421
знак
0
таблиц
0
изображений
2. Генетический анализ бактериофагов
Генетика бактериофагов связана с генетическими особенностями бактерий-хозяев. Признаки бактериофагов, доступные генетическому анализу — это прежде всего скорость и полнота лизиса инфицированных клеток и круг бактерий-хозяев, поражаемых фагами. Широкое распространение в генетическом анализе бактериофагов получили мутанты с условным проявлением. Это мутанты, чувствительные к повышению и понижению температуры, — так называемые термочувствительные (ts) и холодочувствительные (cs). Они нормально размножаются и лизируют клетки только при пермиссивной температуре (37°С). В качестве условных используют также мутанты, чувствительные к действию определенных супрессоров, находящихся в геноме бактерий. Это класс так называемых sus (от англ. suppressor sensitive) мутантов. Sus-мутанты можно получить практически по любому гену, кодирующему белок. Генетический анализ бактериофагов основан на совместном заражении клетки генетически различающимися частицами бактериофага. Полные фаговые геномы, проникая в клетку, экспрессируют заложенную в них информацию подобно гомологичным хромосомам, и таким образом становится возможным исследование взаимодействия аллелей и неаллельных генов. Если два температурочувствительных мутанта, проникших в бактериальную клетку, несут изменения, способные взаимодействовать комплементарно, то в непермиссивных условиях клетки лизируются. В ходе репликации геномов возможна рекомбинация, в результате чего появятся рекомбинанты дикого типа. Их можно учесть, высевая фаговое потомство на чувствительные бактерии в условиях непермиссивных для родительских бактериофаговГенетический анализ бактериофагов сопряжен с рядом трудностей, определяемых особенностями их развития в клетке. Репликация геномов Т-четных фагов идет экспоненциально со временем генерации 2—3 мин. Вплоть до образования количества фаговой ДНК, эквивалентного 50 полным геномам, зрелые частицы фага в клетке не обнаруживаются. Затем из общего фонда синтезируемой ДНК фаговые геномы начинают расходоваться на образование зрелых частиц. При лизисе клетки в ней остается еще столько же ДНК, сколько включилось в головки фагов. В процессе репликации фаговые геномы претерпевают неоднократные циклы спаривания, рекомбинации и репликации. В результате наблюдаемая частота рекомбинации всегда выше ожидаемой. В подобных многократных циклах репликации фаговых геномов генетические события должны быть проанализированы с позиций популяционной генетики. Такой подход разработали Н. Висконти и М. Дельбрюк. При этом они исходили из следующих основных положений: 1) в общем фонде ДНК родительские геномы фагов полностью перемешаны; 2) каждый геном фага аналогичен целой хромосоме; 3) при репликации происходят неоднократные спаривания и рекомбинации. В соответствии с этими допущениями получается, что геномы Т-четных бактериофагов проходят около 5 циклов спаривания и рекомбинации. Однако вызывает сомнение универсальность и надежность исходных допущений. Так, множественность инфекции одной клетки (соотношение родительских частиц) может колебаться до 10 раз. Лизис индивидуальных зараженных клеток обнаруживает неравенство реципрокных классов рекомбинантов. Тем не менее все эти трудности преодолимы, если жестко контролировать соотношение вводимых в клетку геномов фага и время репликации и рекомбинации. При этом можно применять стандартную логику анализа рекомбинации в двух- и трехмаркерных скрещиваниях. На основе этого подхода построены генетические карты бактериофагов ф XI74, ф 80, X, которые представляют собой кольцо, соответствующее кольцевой структуре генома этих бактериофагов. Благодаря знанию генетики двух близких умеренных бактериофагов Я и ф 80 удалось получить их гибриды, совмещающие свойства как X, так и ф 80. Кольцевая карта рекомбинации построена и для Т-четного фага Т4, у которого геном в действительности представлен линейной молекулой ДНК. Этот парадокс объясняется тем, что геномы Т4, упакованные в частицы фага, имеют так называемую концевую избыточность, что допускает кольцевые перестановки, или кольцевые пермутации генов. Дело в том, что в головке Т4 заключено ДНК примерно на 1 % больше, чем соответствует одному геному фага. Дополнительные, концевые, участки фаговой ДНК и составляют физическую основу концевой избыточности. При заражении клетки между молекулами такой ДНК происходят рекомбинации с объединением нескольких геномов в конкатемеры. Длинные молекулы ДНК, состоящие из нескольких геномов, далее реплицируются и рекомбинируют вновь. При созревании бактериофага Т4 работает принцип «наполнения головки». При этом нарезаются молекулы ДНК, в которых наблюдаются концевая избыточность и кольцевые пермутации генов. В молекулах ДНК, полученных из разных источников, встречаются модифицированные основания, например, 5-метилцитозин, 6-метиламинопурин и др. Они не включаются в ДНК при репликации как таковые, а появляются в результате ферментативной модификации синтезируемых молекул, начиная со стадии фрагментов Оказаки. При этом модифицируются основания, занимающие определенное положение в молекуле. Эти реакции осуществляют ферменты, представляющие собой часть единой системы рестрикции — модификации. Модификация определенных оснований предохраняет ДНК от разрушения рестриктирующими эндонуклеазами, которые имеют сродство к тем же последовательностям нуклеотидов, что и ферменты модификации. Ферменты модификации защищают ДНК хозяина от действия эндонуклеаз рестрикции. При этом одна и та же молекула может претерпевать разные типы модификации ДНК. Методом центрифугирования в градиенте плотности в лизатах таких клеток можно обнаружить частицы бактериофага, несущие одну из родительских цепей ДНК и совмещающие типы модификации, характерные для штамма К и для штамма В. Структура многих сайтов рестрикции — модификации в настоящее время расшифрована. Эти сайты представлены короткими нуклеотидными последовательностями, характерная черта которых — их симметричность. Система рестрикции — модификации — это своеобразный барьер, охраняющий клетку от включения в ее генетический материал чужеродных молекул ДНК. Возникновение мутантов, лишенных способности к рестрикции, открывает дополнительные возможности для изменчивости за счет ассимиляции клеткой чужеродной генетической информации. Кроме того, некоторые рестриктазы (например, Eco RI) могут осуществлять сайтспецифическую рекомбинацию плазмид в клетках Е. coli. Успехи генетического анализа у микроорганизмов, особенно у бактерий и бактериофагов, сыграли революционизирующую роль в методах изучения структуры и функций генетического материала. Организация геномов бактерий и пути, ведущие к их рекомбинации, оказались, на первый взгляд, совершенно отличными от того, к чему привыкли генетики, работавшие с эукариотами. У бактерий были открыты дополнительные (к хромосоме) генетические элементы: плазмиды и эписомы. Некоторые эписомы существуют в свободной форме. Это бактериофаги, вся структура которых приспособлена к переносу генома между клетками. Другие плазмиды способны только к репликации в бактериальной клетке. Между этими крайними формами есть промежуточные варианты. Само существование таких дополнительных элементов генома поставило вопрос о возможности их использования для переноса генетического материала и не только между клетками бактерий.
Заключение
Перспективы практического применения рекомбинационной изменчивости во многом связаны с получением живых рекомбинантных вакцин. Весьма актуальным считается получение рекомбинантных вакцин против гриппа, возбудитель которого отличается большой изменчивостью. Принцип рекомбинации современных эпидемических вирусов с хорошо изученным донором аттенуации обеспечивает быстроту формирования клонов с такими полезными свойствами, как термочувствительность и безвредность при сохранении иммунизирующей способности эпидемических штаммов.
Но основании всех вышеперечисленных сведений можно сделать вывод, что генетическая рекомбинация является наиболее удобным методом преобразования биологических свойств вирусов, в частности бактериофагов. Но в настоящее время рекомбинация с успехом используется лишь для передачи признака ослабленной патогенности. На этом основано новое направление применения ts-мутантов для конструирования живых вакцин. Закономерности же передачи и наследования других «полезных» свойств изучены еще недостаточно. Существуют неоднократные указания на большие затруднения в использовании рекомбинации для быстрого выведения высокоурожайных штаммов, пригодных для производства гриппозных вакцин и вирусных диагностикумов. Пока нет какой-либо коллекции мутантов, несущих признак высокой репродуктивной активности.
Список литературы
1. Айала Ф., Кайгер Дж. Современная генетика / Ф. Айала, Дж. Кайгер.- М.: Мир, 1987.- 368 с.
2. Бил Д., Ноулз Д. Внеядерная наследственность / Д. Бил, Д. Ноулз.- М.: Мир,1981.- 138 с.
3. Захаров И.А. Курс генетики микроорганизмов / И.А. Захаров.- Минск: Высшая школа, 1978.- 192 с..
4. Крылов В.Н. Геном бактериофагов / В.Н. Крылов.- М.: Наука, 1989.- 317 с.
5. Хейс У. Генетика бактерий и бактериофагов / У. Хейс. - М.: Наука, 1965.- 254 с.
Похожие работы
... , поскольку во время репликации в системе хозяин-вектор иногда происходит клонирование случайных последовательностей, образовавшихся в результате рекомбинаций, делеций или мутаций. Основной подход состоит в использовании клонированной ДНК в качестве зонда для анализа продуктов эндонуклеазного расщепления суммарной геномной ДНК, из которой произошел данный клонированный сегмент. Такой подход ...
... методы составления генетических карт. 1. Перенос генетического материала у актиномицетов 1.1 Перенос генетического материала с помощью плазмид Это наиболее часто встречающийся способ переноса генетического материала у актиномицетов. Линейные плазмиды актиномицетов были обнаружены раньше, чем линейные хромосомы. Впервые они были описаны у одного из видов актиномицетов в конце 70-х – начале ...
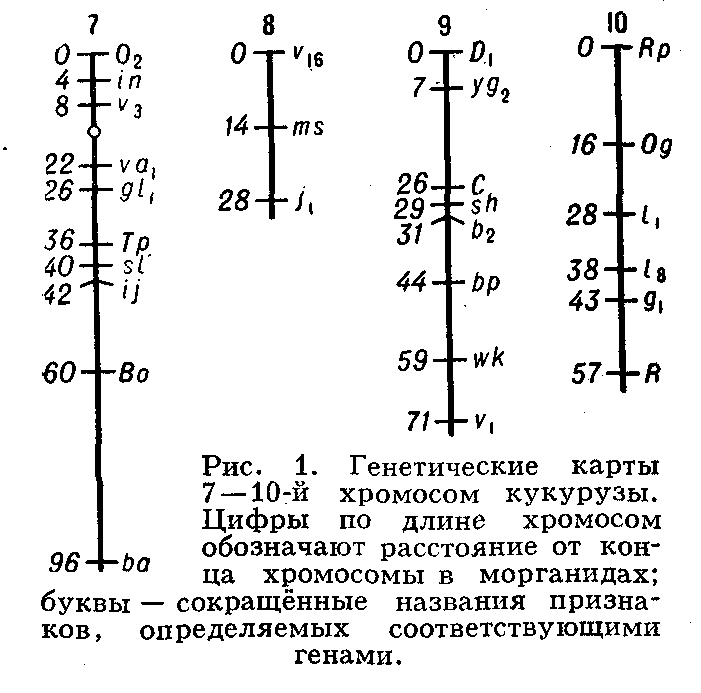
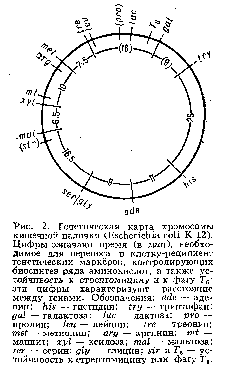
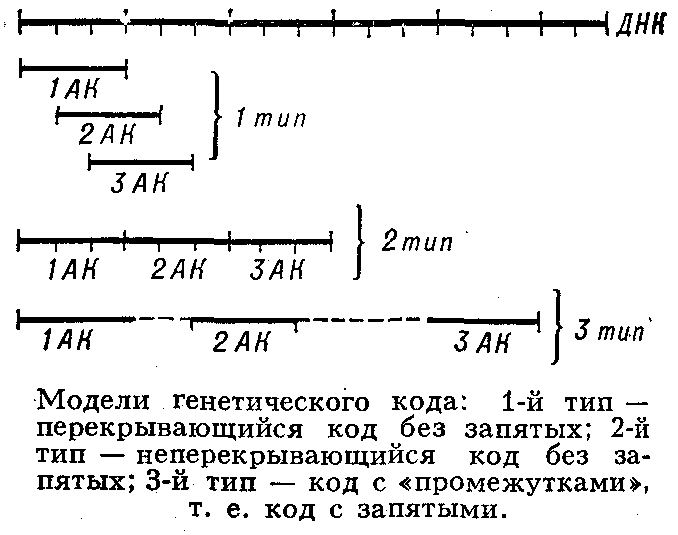
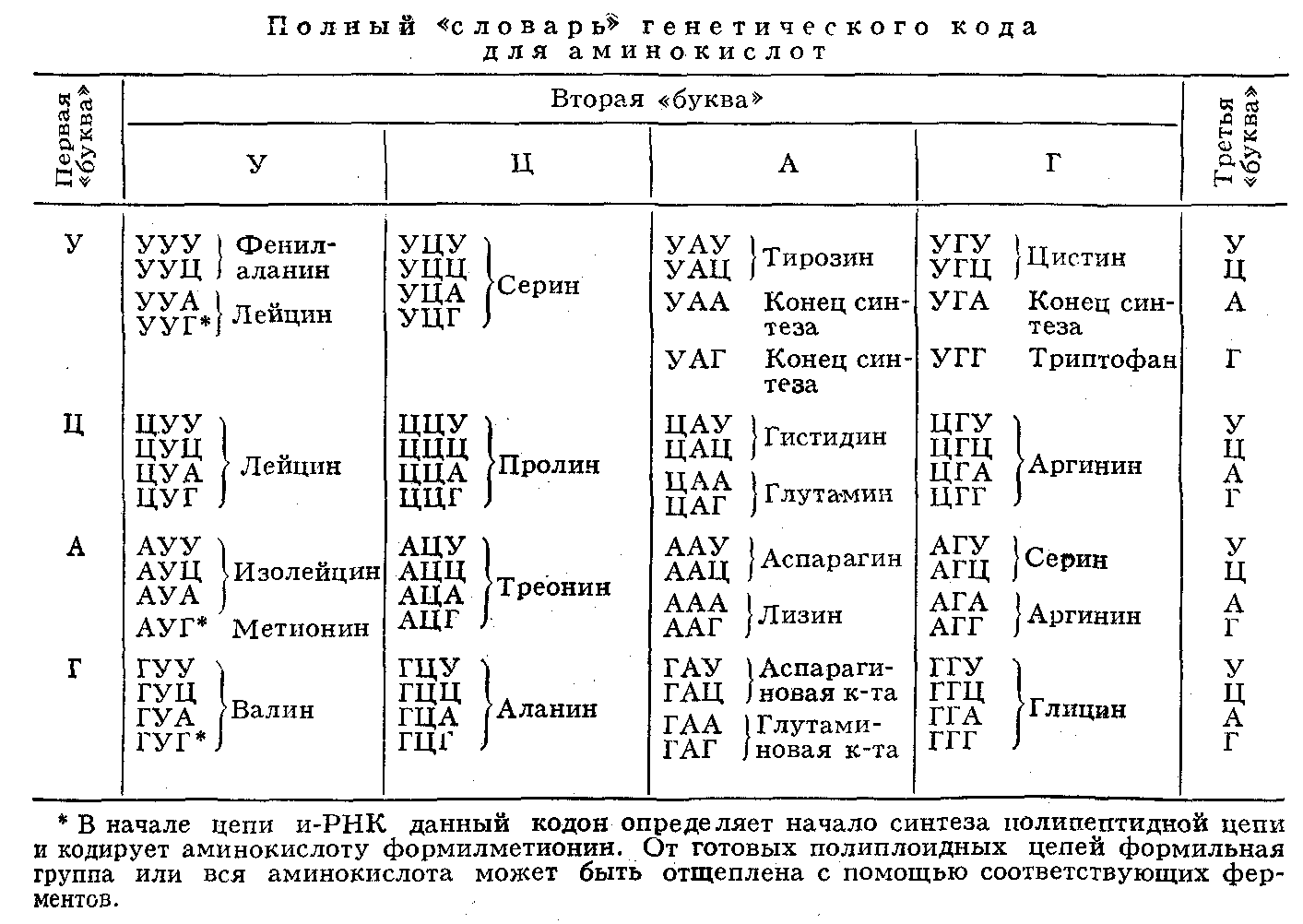
... живые организмы-и удивительное многообразие генов, кодирующих эти белки. В геноме каждого человека есть какие-то области, определяющие его индивидуальность. Некоторые гены человека отличаются от генов крысы всего несколько нуклеотидами-знаками генетического кода. Другие гены у них разные, но одинаковые у двух людей. Изменчивость, связанная с существованием генов , подобных генам группы крови у ...
... один или несколько генов в чистом виде. Такое относительно простое применение принципов и методов, разработанных Важные открытия Как это часто бывает при развитии науки, методологии рекомбинантных ДНК прокладывали путь многие открытия, казавшиеся ранее незначительными и не имеющими отношения к делу. Одним из таких открытий была трансдукция. Другие связаны с выделением и изучением свойств ...
0 комментариев