Реферат
На тему
Свойства основных нейронных сетей
Генетические факторы в развитии зрительных сетей
Определенные свойства основных нейронных сетей формируются еще до того, как животное получает шанс что-либо увидеть. Другие свойства развиваются только в течение первой недели жизни. Ретракция аксонов является неким аналогом событий, происходящих при развитии нервно-мышечного синапса у новорожденных крыс, при рождении каждая концевая пластинка получает сигналы от множества мотонейронов, но в течение нескольких недель большинство аксонов подвергается ретракции, таким образом каждое мышечное волокно получает иннервацию только от одного мотонейрона.
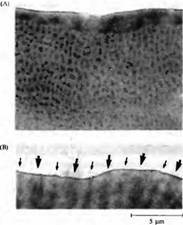
Рис. 1. Архитектура зрительной коры новорожденной обезьяны, не имеющей зрительного опыта, рожденной кесаревым сечением на 8 дней раньше срока и содержащейся в темноте в течение 7 дней. (А) Окраска на цитохромоксидазу, выявляющая пятна в зоне 17. (В) Толстые и тонкие полоски в зрительной зоне 2, отмеченные толстыми и тонкими стрелками.
В результате генетических дефектов в зрительной системе появляются интересные и важные системные аномалии. Например, цветовая слепота у мышей линии reeler, которые имеют аномальное строение зрительной коры. Также примером может служить дефект гена альбинизма у сиамских кошек и у альбиносов, который не только определяет цвет глаз животного через синтез меланина, но также способствует развитию косоглазия в результате систематических ошибок в соединениях между нейронами зрительной коры. Аномальный сенсорный опыт в ранние периоды жизни, и как это может коренным образом изменить анатомию и физиологию центральной нервной системы на всю оставшуюся жизнь животного.
Последствия аномального сенсорного опыта в ранние периоды жизниТри типа экспериментов, большой частью выполненных впервые еще Хьюбелем и Визелем, когда животное лишали нормальных зрительных стимулов. Они изучали физиологические ответы нервных клеток и структуру зрительной системы после (1) закрытия век обоих глаз; (2) предотвращения развития образного зрения (form vision), при сохранном доступе света в глаз; (3) при нормальном попадании света и формирующемся образном зрении, но в условиях искусственно вызванного страбизма (косоглазия) с одной стороны. Эти манипуляции вызывали значительное нарушение в функции и анатомии коры. Особенностью работы Хьюбеля и Визеля является то, что все результаты являются хорошо воспроизводимыми, явными и впечатляющими.
Развитие слепоты после закрытия векКогда веки одного глаза были сшиты вместе на протяжении первых двух недель жизни, обезьяны и котята продолжали развиваться нормально и использовали неоперированный глаз. Через 1—3 месяца, однако, когда открывался оперированный глаз, а нормальный закрывался, становилось очевидным, что животные были практически полностью слепы на оперируемый глаз. При этом котята ударялись о находящиеся вокруг предметы и падали со стола, но не было обнаружено какого-либо значительного физиологического дефекта в оперированных глазах, зрачковый рефлекс оставался нормальным, как и электроретинограмма, которая представляет собой карту суммарной электрической активности глаза. Сигналы от ганглионарных клеток сетчатки у депривированных животных ничем не отличались от нормальных, как и их рецептивные поля.
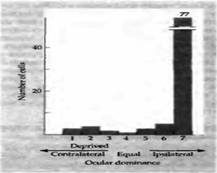
Рис. 2. Последствия закрытия одного глаза. Распределение глазного доминирования у обезьяны, правый глаз которой был закрыт с 21 до 30 дня жизни. Несмотря на то, что в течение последующих 4 лет обезьяна смотрела обоими глазами, большинство кортикальных нейронов остаются нечувствительными к сигналам из правого глаза.
Ответы кортикальных клеток после монокулярной депривации
И хотя ответы клеток ЛКТ после монокулярной депривации практически не изменялись, тем не менее большие изменения происходили в ответах кортикальных клеток. При регистрации электрической активности в зрительной коре было обнаружено очень мало клеток, которые управлялись глазом, подвергнутым депривации. Большинство из тех клеток, которые все же отвечали на стимуляцию этого глаза, имели аномальные зрительные поля. Ответы же клеток недепривированного глаза оставались нормальными. На рис. 2 показаны гистограммы глазного доминирования, полученные для котят и новорожденных обезьян, у которых один из глаз был закрыт на протяжении нескольких первых недель жизни.
Относительная значимость диффузного света и формы объектов для поддержания в норме ответов кортикальных клеток
Результаты экспериментов, описанных нами, говорят о том, что если один глаз не используется в полной мере на протяжении первых недель жизни, его влияние ослабевает и он становится неспособен управлять клетками зрительной коры. Такие значительные изменения вызываются сравнительно небольшой процедурой по закрытию век, без перерезки нервов. Каково же главное условия для поддержания в норме и дальнейшего развития нормальных корковых ответов? Может ли в этом помочь рассеянный свет?
Закрытие век уменьшает количество света, достигающего сетчатки, но не полностью исключает его. Таким образом, можно предположить, что только при помоши диффузного освешения можно заставить глаз нормально функционировать. Или для этого необходимо не просто наличие освещения, но и предметное зрение, которое может быть важным стимулом в предотвращении аномального развития корковых связей? Эти гипотезы были проверены в серии экспериментов, выполненных на новорожденных котятах. Пластиковый полупрозрачный щиток (напоминающий разрезанный шарик для настольного тенниса) помещался поверх роговицы. Он предотвращал возможность различать какие-либо предметы, однако пропускал свет. У всех этих котят развилась слепота на депривированный глаз. Более того, также уменьшалось количество кортикальных клеток, управляемых этим глазом. При этом не регистрировалось каких-либо изменений ни в ответах клеток ЛКТ, ни в клетках самой сетчатки.
Морфологические изменения в ЛКТ после зрительной депривации
Клетки ЛКТ кошки и обезьяны организованы в виде слоев, каждый из которых управляется преимущественно только правым или левым глазом. У тех животных, у которых были обнаружены аномалии строения коры после закрытия век, клетки ЛКТ на первый взгляд выглядели нормальными. Тем не менее было показано, что закрытие век вызывает в них значительные морфологические изменения: клетки становились заметно меньше, чем в слоях, управляемых другим, нормальным глазом. Уменьшение в размерах зависело от продолжительности закрытия век. К удивлению экспериментаторов клетки ЛКТ демонстрировали значительные морфологические изменения одновременно с небольшим функциональным дефицитом. Некоторые из работ дают основания полагать, что размер клеток в ЛКТ может отражать степень их ветвления в коре.
Морфологические изменения в коре после зрительной депривации
Морфологические последствия закрытия одного глаза наиболее хорошо были заметны в слое 4 первичной зрительной коры, где волокна из ЛКТ прерываются, образуя строго определенный паттерн ветвления. Изменения в колонках глазного доминирования, возникающие вследствие закрытия век у обезьян, были выявлены при помощи авторадиографии коры после введения радиоактивных веществ в один глаз. После длительного закрытия век наблюдалось значительное уменьшение ширины колонок глазного доминирования, связанных с закрытым глазом. В то же самое время колонки, управляемые нормальным глазом, соответственно увеличивались в ширине по сравнению с тем, какими они должны быть у взрослых обезьян в норме. Уменьшение колонок глазного доминирования наглядно видно на рис. 3, где нормальные колонки можно сравнить с колонками животных, глаз которых был закрыт в возрасте 2 недель и оставался закрытым на протяжении 18 месяцев. Эти изменения говорят о том, что аксоны из ЛКТ, управляемые нормальным глазом, захватывают и удерживают территорию коры, управляемую более слабым, зрительно депривированным соседним глазом. Эти данные подтверждаются физиологическими наблюдениями по регистрации электрической активности клеток слоя 4. Почти все клетки получали управление только от недепривированного глаза. Некоторые свойства коры менее чувствительны к депривации. Так, например, энуклеация (удаление) одного или даже обоих глаз не вызывало изменений в распределении «пятен» в области V1 и полос при окраске на цитохромоксидазу в области V2.
Критический период чувствительности к закрытию век
При закрытии век одного глаза у взрослых кошек или обезьян не наблюдается каких-либо последствий. Например, у взрослого животного, даже если глаз остается закрытым на протяжении более года, клетки в коре по прежнему получают нормальное управление от обоих глаз и имеют нормальную гистограмму распределения колонок глазного доминирования. Более того, даже если полностью удалить один глаз у взрослой обезьяны, структура слоя 4 остается нормальной, что можно видеть при помощи ауторадиографии и других методов окраски, даже в случае развития атрофии ЛКТ. Эти результаты говорят о значительной устойчивости к изменениям клеток слоя 4 у взрослых, по сравнению с изменениями, наблюдаемыми у незрелых животных.
Клетки коры обезьян имеют наибольшую чувствительность в течение первых 6 недель жизни. В течение любого периода на протяжении этих 6 недель, с максимумом в первую неделю жизни, незначительные изменения в глазном предпочтении и колоночной архитектуре коры возникают даже тогда, когда глаз закрыт на протяжении всего нескольких дней. Затем на протяжении нескольких месяцев (вплоть до 12-16-го месяца жизни) требуется несколько недель, чтобы вызывать заметные изменения в гистограммах глазного доминирования или в ширине колонок слоя 4. После этого времени изменения не развиваются, даже если провести хирургическое удаление одного глаза.
Период наибольшей чувствительности к закрытию век у котят сужен до 4-5 недель после рождения. На протяжении первых трех недель жизни закрытие одного глаза практически никак не влияет на формирование коры. Это и не удивительно, ведь глаза котят в норме закрыты в течение первых 10 дней. Однако в течение 4—5-й недели чувствительность к закрытию резко возрастает. Закрытие глаза в это время даже всего на 3-4 дня приводит к резкому уменьшению количества клеток, управляемых депривированным глазом. Эксперимент по сравнению животных одного помета показан на рис. 4. В данном случае закрытие глаза на протяжении 6 и 8 дней, начиная с 23-го и 30-го дня жизни (рис. 4А и В), приводило к такому же эффекту, как и монокулярная депривация на протяжении трех месяцев с момента рождения. Чувствительность к закрытию век уменьшалась по мере прохождения критического периода и в конце концов исчезала к трем месяцам жизни (рис. 4С). Критический период, однако, может продлиться, если держать котят в темноте. При отсутствии зрительного опыта чувствительность к монокулярной депривации может быть обнаружена даже в возрасте 6 месяцев. Однако нет данных о том, что если котенка в течение непродолжительного времени подержать на свету (для получения зрительного опыта), то это предотвратит продление критического периода.
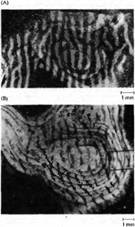
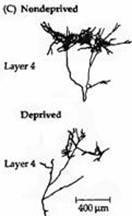
Рис. 3. Колонки глазного доминирования в слое 4 коры после закрытия одного глаза. (А) Нормальная взрослая обезьяна. В правый глаз 10 дней назад была введена радиоактивная смесь пролина и фукозы. В слое 4 можно видеть перемежающиеся светлые и темные полоски одинаковой ширины. Помеченные с помощью радиоактивных меток аксоны из ЛКТ в слое 4 правого полушария выглядят как мелкие белые гранулы, образующие колонки. Находящиеся между ними темные полоски связаны с другим глазом. Данное изображение представляет собой фотореконструкцию параллельных срезов слоя 4 с помощью авторадиографии. (В) Реконструкция слоя 4 у 18-месячной обезьяны, правый глаз которой был закрыт в возрасте 2 недель. Радиоактивная метка вводилась в нормальный глаз. Белые гранулы показывают колонки в слое 4, образованные недепривированным глазом. Они шире, чем в норме. Колонки, управляемые закрытым глазом (показано черным), уже, чем в норме. (С) Корковые разветвления аксонов ЛКТ, заканчивающихся в слое 5 коры котенка, один глаз которого был закрыт на протяжении 33 дней. Терминальное ветвление аксона ЛКТ депривированного глаза показывает значительное уменьшение количества веточек по сравнению с недепривированным глазом.
Восстановление во время критического периода
В какой степени возможно восстановление после депривации во время критического периода? Даже если депривированный глаз после критического периода остается открытым на протяжении месяцев и лет, повреждение является перманентным, происходит лишь небольшое восстановление в редких случаях. Животное на всю жизнь остается слепым на один глаз, что сопровождается уменьшением колонок и асимметрией в гистограмме глазного доминирования. На животных с монокулярной депривацией проводились эксперименты, когда открывался ранее депривированный глаз и закрывался глаз, который раньше был нормальным. Эта процедура, называемая «обратное зашивание», приводила к значительному восстановлению зрения, но только в том случае, если она проводилась на протяжении критического периода. Детеныши обезьян и котята не только начинали снова видеть на изначально депривированный глаз, но они также становились слепыми на другой глаз. Соответственно этим изменениям происходил сдвиг гистограммы глазного доминирования таким образом, что глаз, открытый после депривации, управлял большим количеством клеток, чем глаз, который был открыт на протяжении первых недель и затем закрыт. Более того, анатомическое строение слоя 4, выявленное при помоши авторадиографии, также претерпевало соответствующие изменения: уменьшенные регионы, управляемые первоначально закрытым глазом, расширялись за счет регионов противоположного глаза. На рис. 5 показана гистограмма глазного доминирования и авторадиограмма коры обезьяны, у которой правый глаз был закрыт в возрасте 2 дней на протяжении трех недель. По прошествии этого времени депривированный глаз уже не мог управлять корковыми клетками и колонки, которые им управлялись, уменьшились. После этого правый глаз был открыт, а левый закрыт на протяжении следующих 9 месяцев. В конце этого периода все нейроны управлялись только первоначально депривированным правым глазом и зоны коры, контролируемые им, расширились.
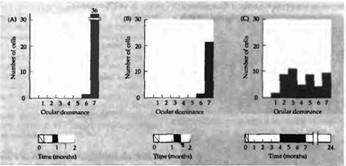
Рис. 4. Критический период у котят. На гистограммах показано глазное предпочтение в зрительной коре котят одного помета, у которых правый глаз был закрыт в разное время жизни. Период, в течение которого глаз оставался закрытым, показан внизу гистограммы. (А) Веки были зашиты на 6 дней в возрасте 23 дней. (В) Веки были зашиты на 9 дней в 30-дневном возрасте. (С) Правый глаз был открыт на протяжении первых 4 месяцев, затем закрыт на протяжении трех, а затем оставался открытым до достижения возраста 2 лет, когда и производилось исследование.
Из этого эксперимента можно сделать несколько выводов: (1) на протяжении критического периода у нормального животного волокна из ЛКТ, направляющиеся в слой 4 коры, подвергаются ретракции, таким образом в коре формируются зоны, контролируемые только одним глазом; (2) закрытие одного глаза во время критического периода приводит к несимметричной ретракции; (3) смена депривированного глаза во время критического периода приводит к повышенному ветвлению (спраутингу (sprouting)) аксонов ЛКТ, в результате они вновь образуют связи с теми клетками, с которыми эта связь была потеряна (см. рис. 5.). Если обратное зашивание выполнить во взрослом состоянии, то оно не дает результатов. Например, у обезьяны, у которой обратное зашивание было выполнено в возрасте 1 года, колонки депривированного глаза, выявленные при окрашивании, так и остались суженными.
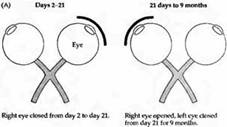


Рис. 5. Влияние обратного зашивания на распределение глазного доминирования и колончатую организацию коры у обезьян. (А) Были выполнены следующие процедуры: правый глаз был закрыт со 2-го до 21-го дня после рождения, после чего был открыт, а левый закрыт с 21 дня до возраста 9 месяцев. (В) Гистограмма глазного доминирования показывает, что почти все клетки получают управление исключительно от правого глаза, который был первично депривирован. Фактически ни одна клетка не получает управления от левого глаза. Таким образом, волокна, получающие управление от правого глаза, вернули себе те корковые клетки, которые потеряли в результате депривации (С) Тангенциальный срез коры, проходящий через слои 4С и 4С. Окрашенные полоски, принадлежащие путям от правого глаза, распространились в слой 4C несмотря на то, что этот глаз подвергся световой депривации на протяжении 19 дней. В течение первых дней колонки, получающие сигналы от правого глаза, уменьшились в размерах, однако после расширились. Восстановление после депривации в других слоях, таких как 4С, не происходит так же хорошо.
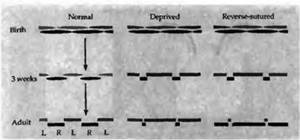
Рис. 6. Результат закрытия одного глаза. У нормальной обезьяны колонки глазного доминирования хорошо определяются в слое 4 коры к возрасту 6 недель. Закрытие век приводит к значительной ретракции волокон, идущих из ЛКТ, связанного с депривированным глазом, в то время как волокна, связанные с нормальным глазом, подвергаются меньшей ретракции, чем обычно, поэтому колонки этого глаза в слое 4 коры более широкие, чем у нормальных взрослых животных. После проведения обратного зашивания, если это сделано в критический период, изначально депривированный глаз может вернуть себе территорию в слое 4, которую он потерял.
Однако концепция строго определенного, жесткого (hard-and-fast) критического периода является сильным упрощением. Эксперименты с обратным зашиванием, выполненные на обезьянах, дают основания полагать, что различные слои в стриарной коре могут развиваться с разной скоростью; критический период может уже закончиться для одного слоя, в то время как соседний еще способен к изменениям в структуре и функции (см. рис.5.С и 6).
Литература
1 Son, Y-J., and Thompson, W.J. 1995. Nerve sprouting in muscle is induced and guided by processes extended by Schwann cells. Neuron 14:33-14.
2 Wilson Hoh, H. L and Sargent, P. B. 1996. Effects of deneation on acetylcholine receptor clusters on frog cardiac ganglion neurons as revealed by quantative laser scanning confoal microscopy. . Neurosci. 16: 1720-1729.
Похожие работы

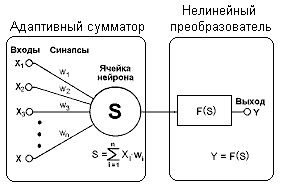
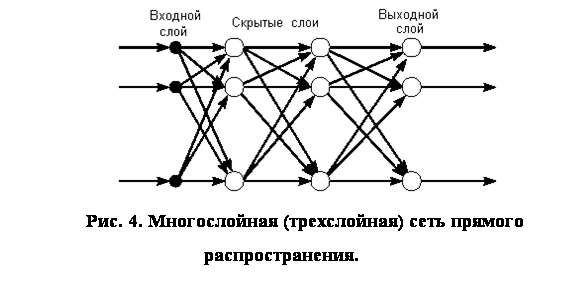
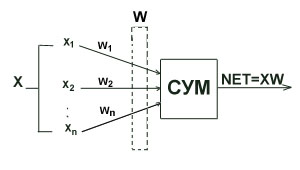
... (15) получить значения для вех нейронов всех слоев. Окончательно формулу (7) для модификации весовых коэффициентов можно записать в виде . (16) Таким образом, полный алгоритм обучения нейронной сети с помощью алгоритма обратного распространения строиться следующим образом. Присваиваем всем весовым коэффициентам сети случайные начальные значения. При этом сеть будет осуществлять какое-то ...

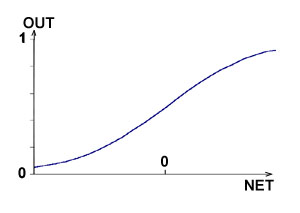
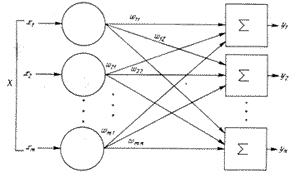
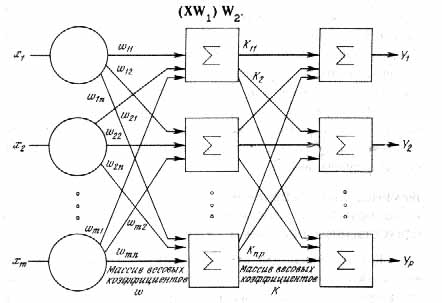
... одном из элективных курсов. Выбор естественно-математического профиля, во-первых, определяется целью введения данного курса в школе (расширение научного мировоззрения) и, во-вторых, сложностью темы в математическом аспекте. Глава 2. Содержание обучения технологии нейронных сетей Авторы данной работы предлагают следующее содержание обучения технологии нейронных сетей. Содержание образования ...

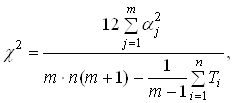
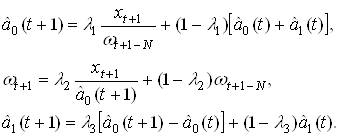
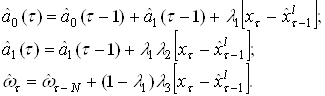
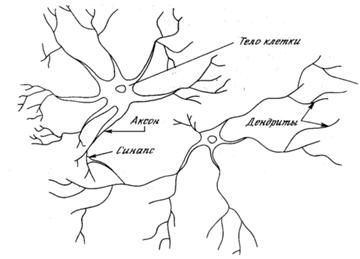
... МП к некритическому экстраполированию результата считается его слабостью. Сети РБФ более чувствительны к «проклятию размерности» и испытывают значительные трудности, когда число входов велико. 5. МОДЕЛИРОВАНИЕ НЕЙРОННЫХ СЕТЕЙ ДЛЯ ПРОГНОЗИРОВАНИЯ СТОИМОСТИ НЕДВИЖИМОСТИ 5.1 Особенности нейросетевого прогнозирования в задаче оценки стоимости недвижимости Использование нейронных сетей можно ...
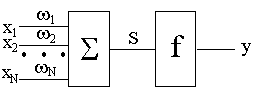
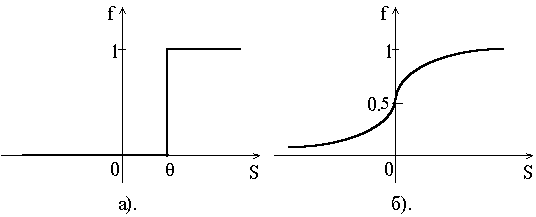
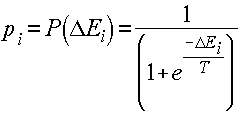
... резонанса. Пожалуй, одна из самых развитых и продуманных с биологической точки зрения концепций нейросетевой обработки информации предложена в работах Гроссберга. Ее стержнем является модель нейронной сети и алгоритмы теории адаптивного резонанса, которая была разработана в начале 70-х годов и детализирована в 80-х. Нейронная система теории адаптивного резонанса способна обучаться распознаванию ...
0 комментариев