Навигация
3. Микро и макроэволюция
Эволюция биомассы на Земле рисуется следующим образом. Разделим существующие организмы на группы:
1) водоросли,
2) морские животные,
3) наземные растения,
4) наземные животные.
Водоросли появились в катархее, и в течение архея и протерозоя их биомасса нарастала до величины порядка сегодняшней, т.е. до 1 млрд. т. Морские животные появились в среднем протерозое, их биомасса очень медленно росла до венда, а в течение кембрия, вероятно, быстро выросла до величины порядка сегодняшней, т.е. до 20-30 млрд. т. Наземные растения появились, возможно, еще в ордовике или даже в кембрии, однако их биомасса стала ощутимой лишь в силуре, а за девон и карбон быстро выросла до колоссальных размеров, порядка нескольких триллионов тонн; в перми она, возможно, даже несколько уменьшилась и приблизилась к современному уровню. Наконец, наземные животные появились только в девоне, их биомасса достигла современной величины, вероятно, еще в палеозое.
Что же касается изменений со временем разнообразия форм организмов, то здесь имеются довольно надежные фактические данные. Они показывают, что в разнообразии форм монотонного нарастания со временем не было, так как при появлении новых форм некоторые из старых отмирали. Так, в кембрии появилось 28 классов животных, в ордовике еще 14 (но часть классов уже отмерла), в силуре еще 3 (но 2 отмерли). Начало верхнего палеозоя ознаменовалось новой вспышкой формообразования - в девоне появилось 9 новых классов (и их число достигло фанерозойского максимума), а карбоне еще 5 (но 8 отмерло), в перми новых классов не появилось (но 4 отмерли). К триасу 3 класса отмерли, но 1 появился (и число классов достигло фанерозойского минимума, если не считать кембрия), в юре и мелу был небольшой прирост. В кайнозое изменений не произошло. Аналогичны данные и по классам растений; здесь также разделяются нижний палеозой, верхний палеозой, мезозой и кайнозой, лишь отмирания классов почти не происходило [7. С. 197].
Жизнь на Земле зародилась, когда на дне мелких теплых морей катархея, богатых сложными органическими веществами до аминокислот включительно, начали образовываться двойные сахаро-фосфатные спиральные нити высокополимерных нуклеиновых кислот с закрепленными на них последовательностями оснований (служащими «кодами» для синтеза белков), способные при некоторых условиях разворачиваться в одинарные спирали и синтезировать на каждой из них недостающую вторую спираль, т.е. порождать пару себе подобных (передавать им информацию о процедурах синтеза белков, закодированных последовательностями оснований). Такие полимеры, которые синтезировали белки, обеспечивающие им достаточно длительное самосохранение, по-видимому, уже можно считать первичными организмами.
Эти первичные микроорганизмы, возможно, питались имевшейся органикой небиологического происхождения, осуществляя, например, бескислородное разложение белков и аминокислот - гниение или углеродов - брожение (анологично питаются современные сапрофиты, поглощая через свои стенки клеток органику биологического происхождения: например, дрожжи при отсутствии кислорода сбраживают глюкозу до спирта и углекислоты). Развитие этих организмов, вероятно, исключило условия для дальнейшего самозарождения жизни, и с тех пор все живое появляется уже только от живого. В результате эволюции микроорганизмов (вынуждаемой нехваткой подходившей для пищи органики) у них появилась способность синтезировать необходимые для самосохранения органические молекулы из неорганических. Наиболее эффективным способом оказался фотосинтез - продуцирование органического вещества из углекислоты и воды под действием солнечного света (энергия света, поглощаемого пигментами, прежде всего зеленым хлорофиллом, расходуется на расщепление молекул воды, кислород выделяется в атмосферу, а водород вместе с углекислотой идет на образование первичного органического продукта - фосфоглицериновой кислоты).
Первыми фотосинтезирующими растениями были, по-видимому, микроскопические синезеленые водоросли цианофиты, у которых хлорофилл рассеян в виде мелких зерен по плазме клеток (у более развитых растений он сосредоточен в специальных тельцах - хлоропластах), и имеется еще синий пигмент фикоцианин. Эти водоросли похожи на бактерий тем, что в их клетках трудно различить ядра, и размножаются они только делением.
Наиболее древние остатки жизнедеятельности организмов, найденные в Трансваале в породах серии Свазиленд возрастом 3.1-3.4 млрд. лет, были тщательно изучены Э. Баргхорном и Дж. Шопфом. Они представляют собой микроскопические изолированные палочки длиной 0.45-0.7 мк. и диаметром 0.18-0.32 мк., имеющие двухслойные оболочки толщиной 0.045 мк.; там же обнаружены нитеподобные образования, а также микроскопические шаровидные, дисковидные и многоугольные оболочки одноклеточных водорослей акритархи. Почти столь же древними (более 2.9 млрд. лет) являются обызвествленные продукты жизнедеятельности цианофитов и бактерий - прикрепленные ко дну столбчатые строматолиты и неприкрепленные округлые онколиты, найденные в известковых прослоях зеленокаменных пород системы Булавайо в Южной Родезии и описанные А. Мак-Грегором еще в 1940 г. [7. С. 204]
Один из наиболее древних следов животных, найден А.М. Лейтесом в среднепротерозойских слоях Удоканской серии Забайкалья. Он представляет собой приуроченные к строматолитовым породам тонкостенные трубочки длиной до 2.5 см, диаметром до 4 мм, имеющие четырехгранное сечение с округленными углами и оставшиеся, по-видимому, от червей. По-видимому, к началу рифея кислорода было уже достаточно для развития примитивных животных. В породах среднего и верхнего рифея найдены следы роющих животных и трубочки червей. В породах венда обнаружены уже многочисленные отпечатки разнообразных бесскелетных животных, а начало кембрия маркируется массовым появлением скелетообразующих организмов.
Палеонтологические данные позволили воссоздать грандиозную и стройную картину эволюции форм жизни в фанерозое, не оставляющую никаких сомнений в естественности этого процесса. Проследим фанерозойскую эволюцию ряда важнейших типов организмов.
Начнем с растений, которые осуществляют фотосинтез первичного органического вещества и потому являются первоосновой всей жизни. Водоросли появились еще в начале архея (или, может быть, даже в катархее) и с тех пор благополучно существуют до настоящего времени преимущественно в виде одноклеточных форм. Доля водорослей в общей массе живых организмов на Земле в наше время составляет меньше одной тысячной (около 1.7 млрд. т из общей суммы в 2-3 триллиона т), но зато они чрезвычайно быстро размножаются - их клетки делятся каждые 1-2 суток, и в результате их продукция, около 550 млрд. т в год, составляет основную долю продукции биомассы на Земле в целом.
Первые наземные растения появились, вероятно, в ордовике или даже в кембрии, но достоверные их следы найдены лишь в силуре. Это были самые примитивные из высших растений типа сосудистых - псилофиты, мелкие растения без корней и листьев, а также первые плауны. В девоне появились представители двух других подтипов сосудистых растении - клинолистных (хвощей) и папоротникообразных (из последних - пока лишь наиболее примитивные растения класса папоротников а также отряда семенных папоротников птеридосперм из класса голосемянная), причем крупные древовидные формы плаунов хвощей и папоротников образовали первые леса нашей планеты. В карбоне обширные участки суши покрыли огромные заболоченные леса из гигантских плаунов лепидодендронов и сигиллярий, хвощей каламитов, древовидных папоротников и птеридосперм, а также новых отрядов вечнозеленых голосемянных кордаитов, а к концу периода - первых хвойных растении и гинкго. В перми появились бенеттиты, плауны пришли в упадок; на территориях Гондваны господствовала специфическая растительность во главе с голосемянными глоссоптерисами.
Триас и юра - это время расцвета голосемянных растений отрядов бенеттитов, хвойных, гинкго и цикадовых (саговников); птеридоспермы и кордаиты вымерли. В нижней половине мела, а возможно даже в юре, появились первые покрытосемянные (цветковые) растения, сначала двудольные. Во второй половине мела появились также однодольные цветковые растения, а двудольные образовали уже леса (кленовые и дубовые). Они вытеснили ряд отрядов голосемянных - бенеттиты вымерли, цикадовые и гинкго пришли в упадок. Наступивший затем кайнозой - эра расцвета цветковых растений.
Рассмотренная эволюция форм растений от водорослей до цветковых ттооисходила по линии уменьшения роли двуполых гаплоидных поколении, в телах которых (гаметофитах) клетки содержат одинарные наборы хромосом, и увеличений роли бесполых диплоидных поколений, в телах которых клетки содержат двойные наборы хромосом. Так, у многоклеточной водоросли улотрикс имеется только 1 диплоидная клетка, остальные - гаплоидны. У покрытосемянных, наоборот, все растение гаплоидно а гаметофит состоит лишь из нескольких клеток в тканях цветка; эволюционная выгода этого заключается как в том, что для размножения цветковых не требуется внешней влаги, так и в меньшей чувствительности диплоидных организмов к влиянию рецессивных генов (таких, которые действуют лишь при наличии их в обоих наборах хромосом в диплоидной клетке). В наше время наземные растения, преимущественно цветковые и хвойные, составляют 98-99% от 2-3 триллионов т биомассы на Земле, но растут они медленно - их годичная продукция составляет всего около 50 млрд. т (вдесятеро меньше продукции водорослей в океане), а средний возраст (очевидно, возраст деревьев в лесах) - 40-60 лет [7. С. 211].
Переходя к эволюции животных, отметим, что сведения палеонтологии о времени появления, расцвета и вымирания тех или иных групп животных вместе с данными о степени сложности их устройства, способах размножения и особенностях развития их зародышей позволяют судить о происхождении этих групп. Современные представления о происхождении основных типов животных (базирующиеся на принципе монофилии, т.е. на предположении о наличии единого корня происхождения каждой группы организмов) показаны на рис. 1. Согласно этим представлениям, одноклеточные простейшие животные породили как тупиковую ветвь губок, так и прогрессивную ветвь кишечнополостных, от которой пошли два главных ствола - первичноротых и вторичноротых. На первом из этих стволов промежуточным этапом были низшие черви, от которых пошли ветви немертин, плеченогих, мшанок, форонид, моллюсков и через кольчатых червей, наконец, членистоногие. От другого ствола поочередно отделялись ветви щетинкочелюстных червей, иглокожих, погонофор, кишечнодышащих, оболочников, головохордовых и, наконец, позвоночных.
Ряд типов животных - по крайней мере простейшие, кишечнополостные и некоторые низшие черви - появился, хотя бы в небольших популяциях, еще в среднем протерозое. Число семейств - одна из возможных количественных характеристик разнообразия форм данного типа организмов, свидетельствующая о темпах его эволюции: в периоды быстрой эволюции разнообразие форм должно возрастать.
К сожалению, мы не имеем возможности остановиться на истории развития каждого из типов животных - напомним, что всего имеется 23 типа!
Поэтому о первичноротых мы скажем лишь то, что все их типы появились, по-видимому, еще в докембрии. Нам придется оставить в стороне такие интересные и важные вопросы, как, например, связанная с эволюцией кишечно-полостных история коралловых рифов или же охватывающая весь фанерозой история «приматов моря» - головоногих моллюсков (не раз порождавших и массовые формы, и таких гигантов, как наутилусы ордовика с 6-метровыми прямыми раковинами, меловые аммониты со спиральными раковинами 3-метрового диаметра и современные гигантские «кракены» - глубоководные кальмары архитевтисы длиной до 15-20 м.).
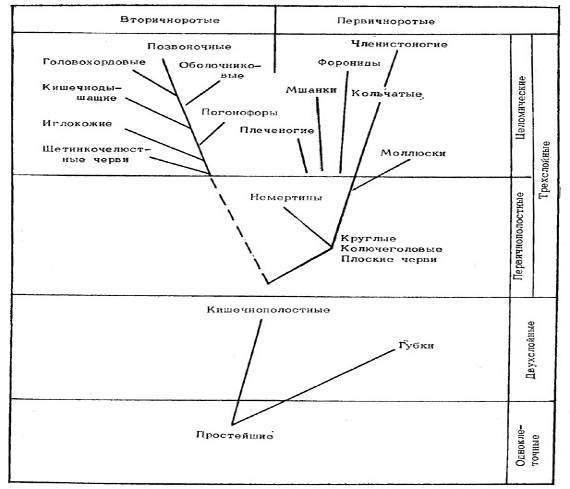
Рис. 1. Происхождение основных типов животных.
Из вторичноротых рассмотрим лишь историю позвоночных. Первые их представители появились в кембрии, это так называемые панцирные рыбы - небольшие рыбообразные донные животные без челюстей и плавников, покрытые панцирем из костных пластинок на голове и толстых чешуи на туловище; они еще не были настоящими рыбами - из современных позвоночных животных к ним ближе всего бесчерепные (ланцетники) и бесчелюстные - круглоротые (миноги и миксины). В силуре появились, а в девоне получили широкое развитие уже и настоящие рыбы, как хрящевые (сначала панцирные, а затем и беспанцирные акулы, благоденствующие с тех пор по настоящее время), так и костные, уже в середине девона поделившиеся на лопастеперых (двоякодышащих и кистеперых) и лучеперых, сначала ганоидных в триасе, а с юры и костистых, имеющих абсолютное господство среди рыб в настоящее время. Девон был временем первых попыток рыб выбраться на сушу; двоякодышащие рыбы в этом не преуспели, а от кистеперых пошли земноводные животные (амфибии), которые осваивали сушу одновременно с насекомыми.
Первые настоящие наземные позвоночные - это пресмыкающиеся (рептилии), у которых размножение и развитие происходят на суше. История рептилий перми и мезозоя, этих двухсот миллионов лет господства чудовищ на нашей планете, является, вероятно, самой драматической главой истории жизни на Земле до появления человека. Современное представление о пресмыкающихся, - как о холоднокровных животных с чешуйчатой кожей, плохо приспособленных к среде с низкой или резко колеблющейся температурой, возможно, не следует распространять на динозавров. Для выяснения этого вопроса Роберт Беккер (1975 г.) использовал три признака, отличающих теплокровных животных от холоднокровных:
1) наличие в костях большого количества кровеносных сосудов, а также каналов, в которых происходит быстрый обмен фосфатом кальция, необходимый для нормального функционирования мышц и нервов;
2) малое отношение биомасс хищников и их жертв (порядка 1-5% против десятков процентов у холоднокровных животных);
3) способность существовать в условиях холодного климата [8. С. 92].
По всем трем признакам установлено, что если раннепермские хищные пеликозавры (например, диметродоны) еще были холоднокровными, то уже позднепермские терапсиды и позднетриасовые текодонты были теплокровными (отношения хищник-жертва порядка 10-15%); у динозавров признаки теплокровности выражены даже ярче, чем у современных млекопитающих (отношение хищник-жертва всего 1-3%, так что находки костей хищных динозавров чрезвычайно редки). Динозавры, особенно жившие в высоких широтах, вероятно, имели и волосяные покровы.
Таким образом, эволюция форм организмов происходит, во-первых, вследствие мутаций (вызываемых физико-химическими воздействиями случайных изменений в нуклеинокислотных носителях наследственности) и, во-вторых, вследствие отбора тех мутантов, которые оказываются способными к адаптации (т.е. к приспособлению) к окружающей их живой и неживой природе. В большинстве случаев способными к адаптации оказываются мутанты с полезными усложнениями организмов; полезные упрощения возникают гораздо реже, и жизнь в целом эволюционирует от простого к сложному. Иногда же возникают особенно удачные формы организмов, столь хорошо приспособленные не только к среде, окружавшей их в период образования, но и к широкому спектру ее изменений, что им удается просуществовать чрезвычайно долго, до сотен миллионов лет включительно. Иначе говоря, высокая мера адаптированности приводит к низкой мере эволюционирования (и наоборот) [8. С. 89].
Человеческое общество оказывает все усиливающееся влияние на развитие жизни на Земле, которое началось в форме охоты, рыболовства и истребления опасных для человека животных и вытеснения дикой жизни из населенных районов, а в наши дни идет по пути создания и разведения культурных сортов растений и пород животных и может, в конце концов, привести почти к стопроцентному управлению развитием всех форм жизни.
Это может касаться и вида Homo sapiens, развитие которого определяется не столько биологическими, сколько социальными факторами - прежде всего уровнем производительных сил общества и производственными отношениями его членов (и такими надстройками, как складывающаяся стихийно или формируемая сознательно общественная мораль, которая, например, в прошлые столетия вследствие высокой детской смертности поощряла многодетность; в последние же десятилетия при снижении детской смертности, но создаваемых урбанизацией жилищных ограничениях это отношение изменяется).
Заключение
В современной теории эволюции значительное место занимают вопросы происхождения жизни на Земле, ибо в рамках теории эволюции происхождение жизни рассматривают в качестве изначального процесса образования живых форм (необиогенеза) из неорганических веществ, т.е. в виде химической эволюции. В наше время абиогенезу противопоставлена концепция постепенного возникновения и развития живого материала в течение длительного периода времени. Эта концепция получила название архебиоза или археогенеза. В соответствии с теорией Опарина жизнь является результатом исторического односторонне направленного развития в виде постепенного усложнения органических субъединиц и развития их в сложные системы, обладающие свойствами живого. Несмотря на остающуюся неопределенность в вопросе о происхождении жизни на Земле, не вызывает сомнения факт в естественности ее происхождения как определенного закономерного этапа развития материи.
1. Возникновение жизни на Земле подготовлено ходом эволюции неживой материи во Вселенной.
2. Существование жизни на Земле определяется постоянством действия космических факторов: мощностью и спектральным составом солнечного излучения, неизменностью основных характеристик орбиты Земли и ее осевого вращения, наличием магнитного поля и атмосферы планеты.
3. Развитие жизни на Земле во многом обусловлено плавными незначительными изменениями космических факторов; сильные изменения ведут к катастрофическим последствиям: космические лучи и их рассмотрение как мутагенных факторов.
4. На определенном этапе своего развития жизнь становится фактором космического масштаба, оказывающим влияние на физико-химические характеристики основных оболочек планеты (например, состав и температуру атмосферы, гидросферы и верхних слоев литосферы).
Список используемой литературы
1. Барг О.А. Живое в едином мировом процессе. Изд-во Пермского университета 1993. – 227 с.
2. Вселенная, астрономия, философия. М.: изд. МГУ 1988. – 192 с.
3. Данилова В.С., Кожевников Н.И. Основные концепции современного естествознания: учебник, М.: Аспект-пресс, 2000 – 256 с.
4. Карпенков С.Х. Современное естествознание: учеб., М.: Академический проект 2003. – 560 с.
5. Найденыш В.М. Концепции современного естествознания: Учеб. пособие д/вузов, М.: Гардарики 2002, - 476 с.
6. Соколов Е.Ф. Концепции современного естествознания: Учеб. пособие д/вузов, М.: ВЛАДОС, 1999. – 232 с.
7. .Шкловский И.С. Вселенная, жизнь, разум – М.: Наука, 1997 г. – 320 с.
8. Щербаков В.П. Эволюция как сопротивление энтропии – М.: ЭКСМО, 2006 г. – 125 с.
Похожие работы
... и К.А.Тимирязев: «Мы вынуждены допустить, что живая материя осуществлялась так же, как и все остальные процессы, путём эволюции… Процесс этот, вероятно, имел место и при переходе из неорганического мира в органический» (1912).- 5 - 3. Гипотеза происхождения жизни А.И. Опарина Еще Ч.Дарвин понял, что жизнь может возникнуть только при отсутствии жизни. В 1871 г. он писал: «Но если бы сейчас ...

... 6. Свое видение вопроса Изучив вышеуказанные варианты происхождения жизни на Земле, я убедился, что в каждой концепции есть свои минусы. Мне пришлось взять на себя смелость и озвучить свою концепцию происхождения жизни на Земле. Для этого мне придется объединить креационизм, уникальное однократное зарождение жизни и панспермию в одно целое. В свое время мне пришлось прочитать немало книг по ...
гом», описанный в Библии, гипотезы Аристотеля, Эпикура и Демокрита. Исследования Луи Пастера в 19-м веке окончательно подтвердили ошибочность представлений происхождения жизни как о спонтанном самозарождении. Правда, они не дали окончательных выводов о происхождении жизни. И только 3 мая 1924 г. на собрании Русского ботанического общества ученый А. И. Опарин с новой точки зрения рассмотрел ...
... содержание отдельных металлов одинаково у бактерий, губок, растений, животных и в океанской воде. Однако вернемся к началу XX в. Все большее число ученых склонно признать, что проблема возникновения жизни не может быть решена наукой. Основания для такого мнения налицо: тысячелетнее господство религиозных мифов о сотворении мира и наивные представления о самозарождении заменяются умозрительными ...
0 комментариев