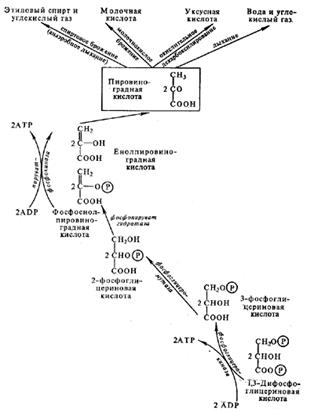
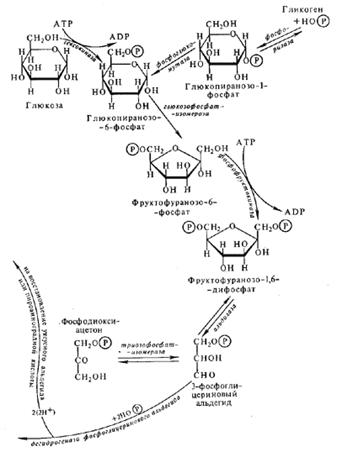
Дихотомический путь
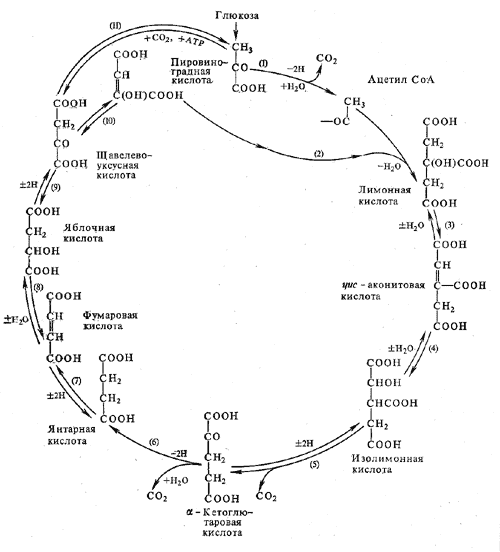
Цикл Кребса. Механизмы регуляции цикла. Энергетическая эффективность процесса, значение
Пентозофосфатный путь. Механизмы регуляции цикла. Энергетическая эффективность процесса, значение. Связь с другими процессами
Глиоксилатный цикл. Механизмы регуляции цикла. Энергетическая эффективность процесса, значение
Митохондрии как органоиды дыхания. Их структура и функции
Генетическая связь дыхания и брожения. Связь дыхания и фотосинтеза. Взаимосвязь дыхания с другими процессами обмена
Количественные показатели газообмена
Навигация
Количественные показатели газообмена
Дыхание растений
87296
знаков
0
таблиц
14
изображений
9. Количественные показатели газообмена
Интенсивность дыхания – количество поглощенного кислорода (выделенного углекислого газа) за 1 час 1 граммом растительного материала.
Дыхательный коэффициент - отношение объёма выделяемого из организма углекислого газа к объёму поглощаемого за то же время кислорода. Зависит от химической природы дыхательного субстрата, содержания CO2 и O2 в атмосфере и др. факторов, характеризуя, т. о., специфику и условия дыхания. При использовании клеткой для дыхания углеводов (проростки злаков) ДК равен примерно 1, жиров и белков (прорастающие семена масличных и бобовых) — 0,4—0,7. При недостатке О2 и затруднённом его доступе (семена с твёрдой оболочкой) ДК равен 2—3 и более; высокий ДК характерен также для клеток точек роста.
Р/О – отношение количества образующегося АТФ к количеству поглощенного кислорода за единицу времени единицей растительной массы. Показывает насколько активно идут процессы в ЭТЦ митохондрий, насколько энергетически эффективно дыхание.
10.Регуляция процесса дыхания. Зависимость дыхания от внутренних факторов
Дыхательный контроль. Возрастание функциональной активности клеток сопровождается усилением дыхания. В значительной степени это достигается благодаря механизму дыхательного контроля, или акцепторного контроля дыхания. Дыхательным .контролем называют зависимость скорости потребления 02 митохондриями от концентрации АДФ, который служит акцептором фосфата при окислительном фосфорилировании. В условиях полного сопряжения транспорта электронов по ЭТЦ с синтезом АТФ интенсивность дыхательного процесса в митохондриях зависит от концентрации АДФ или, точнее, от отношения действующих масс АТФ-системы: [ATP]/[АДФ] [PJ. Причем неорганический фосфат обычно присутствует в достаточном количестве и не является ограничивающим фактором. В клетке, находящейся в состоянии покоя, это отношение достаточно велико, так как почти весь АДФ фосфорилирован. При увеличении функциональной активности клеток АТФ расходуется на энергозависимые процессы, в результате чего возрастает концентрация АДФ, а это в свою очередь приводит к повышению скорости переноса электронов и интенсивности окислительного фосфорилирования. Важно подчеркнуть, что в данном случае уровень АДФ регулирует интенсивность транспорта электронов и окислительное фосфорилирование не как аллостерический фактор, а как субстрат фосфорилирования.
Более полно состояние адениннуклеотидной системы выражается отношением, получившим название энергетического заряда:
[АТФ] + у2 [АДФ] [АТФ] + [АДФ] + [AMP] '
который характеризует меру заполнения всей адениннуклеотидной системы высокоэнергетическими фосфатными группами.
Эффект Пастера. Уровень 02 в тканях влияет не только на интенсивность дыхания, но определяет и величину расходования дыхательных субстратов, на что впервые обратил внимание Л.
Пастер. В его опытах с дрожжами в присутствии 02 снижались распад глюкозы и интенсивность брожения (уменьшалось количество спирта и выделяемого С02), но одновременно наблюдался интенсивный рост биомассы дрожжей вследствие усиления использования Сахаров на синтетические процессы. Торможение распада Сахаров и более эффективное их использование в присутствии кислорода получило название «эффекта Пастера».
Механизм эффекта Пастера состоит в том, что в присутствии 02 интенсивно идущий процесс окислительного фосфорилирования конкурентно уменьшает количество молекул АДФ, вступающих в гликолиз (на нужды субстратного фосфорилирования). По этой причине, а также из-за тормозящего действия АТФ (синтез которого резко возрастает в аэробных условиях) на фосфофруктокиназу, скорость процессов гликолиза в присутствии 02 снижается. Избыток АТФ может способствовать и ресинтезу глюкозы из части молекул пирувата, образующегося в ходе гликолиза. Без кислорода не функционируют цикл Кребса и ПФП и, следовательно, клетки не получают многих промежуточных соединений, необходимых для синтеза клеточных структур. В присутствии 02 все эти циклы работают. Увеличение концентрации молекул АТФ в условиях аэробиоза также способствует синтетическим процессам.
Изменение интенсивности дыхания в онтогенезе. У светолюбивых растений более высокая интенсивность дыхания по сравнению с теневыносливыми. Растения северных широт дышат более интенсивно, чем южные, особенно при пониженной температуре. Наиболее высока интенсивность дыхания у молодых активно растущих тканей и органов. После окончания роста дыхание листьев снижается до уровня, равного половине максимального и затем долго не меняется. При пожелтении листьев и в период, предшествующий полному созреванию плодов, у этих органов наблюдается активация синтеза этилена с последующим кратковременным усилением дыхания, которое называют климактерическим подъемом дыхания. Этилен увеличивает проницаемость мембран и гидролиз белков, что приводит к повышению содержания субстратов дыхания. Однако это дыхание не сопровождается образованием АТФ.
11.Зависимость процесса дыхания от факторов внешней среды
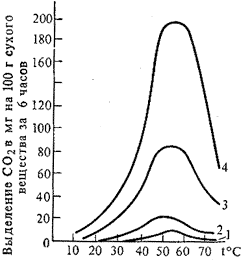
Температура. Дыхание у некоторых растений идет и при температуре ниже 0оС. Так, хвоя ели дышит при –25оС. Интенсивность дыхания, как всякой ферментативной реакции, возрастает при повышении температуры до определенного предела (35-40оС).
Кислород необходим для осуществления дыхания, так как он является конечным акцептором электронов в дыхательной электронтранспортной цепи. Увеличение содержания кислорода в воздухе до 8-10 % сопровождается повышением интенсивности дыхания. Дальнейшее увеличение концентрации кислорода существенно не влияет на дыхание. Однако в атмосфере чистого кислорода дыхание растений снижается, а при длительном его действии растение погибает. Гибель растения обусловлена усилением в клетках свободнорадикальных реакций и повреждением мембран вследствие окисления их липидов.
Углекислый газ является конечным продуктом дыхания. При высокой концентрации газа дыхание растений снижается по следующим причинам: 1) ингибируются дыхательные ферменты, 2) закрываются устьица, что препятствует доступу кислорода к клеткам.
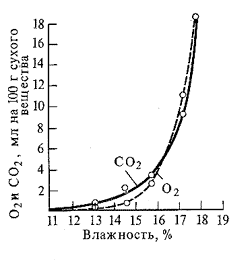
Содержание воды. Водный дефицит растущих тканей увеличивает интенсивность дыхания из-за активации распада сложных углеводов (например, крахмала) на более простые, которые являются субстратом дыхания. Однако при этом нарушается сопряжение окисления и фосфорилирования. Дыхание в этом случае представляет бесполезную трату вещества. Иная закономерность характерна для органов, находящихся в состоянии покоя. Повышение содержания воды в семенах приводит к резкому увеличению интенсивности дыхания.
Свет. Трудно выявить влияние света на дыхание зеленых растений, так как одновременно с дыханием осуществляется противоположный процесс – фотосинтез. Освещенность, при которой интенсивность фотосинтеза равна интенсивности дыхания по уровню поглощенного и выделенного углекислого газа, называют компенсационным пунктом. Дыхание незеленых тканей активируется светом коротковолновой части спектра, так как максимумы поглощения флавинов и цитохромов расположены в области 380-600 нм.
Минеральные вещества. Такие элементы как фосфор, сера, железо, медь, марганец необходимы для дыхания, являясь составной частью ферментов или как фосфор промежуточным продуктом. При повышении концентрации солей в питательном растворе, на котором выращивают проростки, их дыхание активируется (эффект «солевого дыхания»).
Механическое повреждение усиливает дыхание из-за быстрого окисления фенольных и других соединений, которые выходят из поврежденных вакуолей и становятся доступными для оксидаз.
Список использованной литературы
1. Алехина Н.Д., Балнокин Ю.В., Гавриленко В.Ф. и др. под ред. Ермакова И.П. Физиология растений. – М.: Академия, 2004
2. Грин Н., Стаут Т., Тейлор Д. Биология т.2. – М.: Мир, 1990
3. Малиновский В.И. Физиология растений. – Владивосток: Изд-во ДВГУ, 2004
4. Полевой В.В. Физиология растений. – М.: Высшая школа, 1983
5. Рубин Б.А., Ладыгина М.Е. Физиология и биохимия дыхания растений. – М.: Наука, 1974
6. Якушкина Н.И. Физиология растений. – М.: Просвещение, 1993
Похожие работы
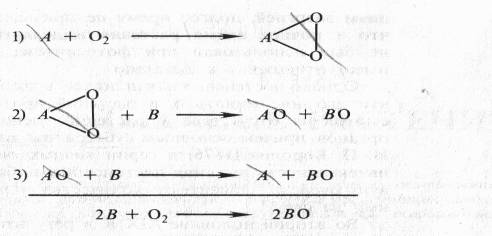
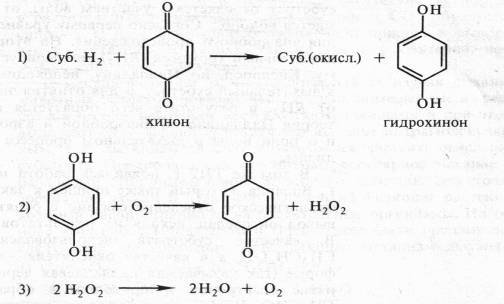
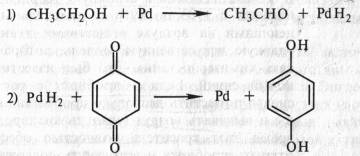
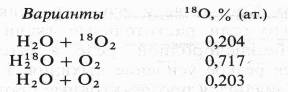
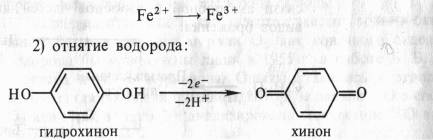
... дыхание — начальный этап нормального аэробного дыхания. Эту точку зрения поддержал Б. Пфеффер — немецкий физиолог растений, который распространил ее на растительные организмы. На основе этих работ Пфеффером и Пфлюгером были предложены следующие два уравнения, описывающие механизм дыхания: На первом, анаэробном, этапе происходит спиртовое брожение, образуются две молекулы этанола и две ...
... , часть накопленных в процессе фотосинтеза углеводов теряется. Однако все попытки искусственно ингибировать фотодыхание приводили к общему снижению интенсивности фотосинтеза. На современном этапе развития физиологии растений принято считать, что основное значение фотодыхания заключается в его защитной роли. Сбрасывая таким образом избыточную энергию, растение избегает разрушения фотосистем, ...
... цитохромоксидаза, подтверждением чего служит также максимальная величина подавления дыхания азидом . Соотношения указанного типа не являются, однако, общей закономерностью, поскольку в ряде случаев максимальная интенсивность дыхания обнаруживается в клетках зоны растяжения. Такая закономерность выявляется при пересчете на белок, а также на единицу кислотонерастворимого фосфора. Вполне вероятно, ...
... дальнейшим изменениям. Правильность своей теории Бах подтвердил на сотнях примеров и фактов, как известных до него, так и полученных им самим экспериментально. История современного учения о дыхании растений неразрывно связана с именем академика В.И. Палладина. В годы первого петербургского периода работы Палладин исследовал ферментативную природу дыхательного процесса. Палладин показал, что и ...
0 комментариев