Навигация
2.2 Мейоз
Мейоз ( от греч. «мейезис» – уменьшение ) – способ деления клеток, при котором в отличие от митоза происходит редукция (уменьшение) количества хромосом и переход клеток из диплоидного состояния в гаплоидное.
У растений мейоз – основа образования гамет и гаплоидных спор. Мейоз состоит из двух последовательных делений ядра, в процессе которых удвоение ДНК происходит один раз. Отличительной особенностью первого деления мейоза является сложная и растянутая во времени профаза. Хромосомы принимают вид длинных тонких нитей. Гомологичные хромосомы спариваются (конъюгируют) друг с другом. Каждый гомолог происходит от одного из родителей и состоит из двух идентичных хроматид. Таким образом, гомологичные пары состоят из четырех хроматид. Конъюгация гомологичных хромосом является обязательной частью мейоза. В гаплоидных клетках конъюгация не встречается, так как в них отсутствуют гомологи.
Объединенные пары гомологичных хромосом называются бивалентами. В это время четыре прижатые друг к другу хроматиды могут обмениваться отдельными участками (кроссинговер). В любой данной точке кроссинговер может быть у двух хроматид. В результате кроссинговера хроматиды имеют другой состав генов. В конце профазы I фрагментируются покровы ядра и образуется веретено деления. В метафазе I биваленты размещаются по экватору веретена, образуя экваториальную пластинку. Центромеры гомологичных хромосом расположены по противоположным сторонам от экваториальной плоскости. В метафазе митоза наоборот: центромеры отдельных хромосом находятся в экваториальной плоскости. В анафазе I начинается движение гомологичных хромосом к противоположным полюсам клетки. В анафазе митоза центромеры делятся и расходятся идентичные хроматиды. В анафазе I мейоза центромеры не делятся, хроматиды остаются вместе, а разъединяются гомологичные хромосомы. Однако из-за обмена фрагментами в результате кроссинговера хроматиды не идентичны, как в начале мейоза.
Завершается первое деление телофазой I, когда спирализация хромосом ослабляется, они удлиняются и вновь становятся невидимыми.
Телофаза переходит в интерфазу. Но у многих организмов интерфаза отсутствует. Затем в каждой из дочерних клеток происходит второе деление митоза. Во втором делении митоза также выделяют стадии: профаза II, метафаза II, анафаза II, телофаза II. По сущности, это митоз в клетке с гаплоидным набором хромосом. При этом к противоположным полюсам расходятся сестринские хроматиды. Несмотря на то, что в процессе мейоза генетический материал удваивается только один раз, происходит два деления, в результате чего образуется четыре клетки, каждая с уменьшенным вдвое (гаплоидным) количеством хромосом, а благодаря кроссинговеру, и с новыми, отличными от родительских, типами хромосом. При митозе наборы хромосом дочерних ядер и материнского были идентичными.
Эндомитоз – внутреннее деление ядра, которое отличается от обычного кариокинеза тем, что ядрышко и покровы ядра сохраняются, а хромосомы разделяются в продольном направлении, и все остаются в составе одного ядра. Встречается у видов родов Lycopersicum, Taraxacum. Распространенность и механизм эндомитоза пока не известны.
3. Онтогенез растительной клетки
С момента образования и до отмирания клетки проходят определенный путь развития, в котором выделяют три основные фазы: эмбриональную, растяжения и дифференциации.
Эмбриональная фаза развития – процесс новообразования клеток путем их деления. Образовавшиеся в результате деления клетки мелкие, расположены плотно, оболочка тонкая, богатая пектином, мало клетчатки. В цитоплазме развита ЭПР, много рибосом.
Фаза растяжения. Клетки растут, увеличиваются в объеме. Протопласт вакуолизируется. Мелкие многочисленные вакуоли сливаются, размер их увеличивается, количество сокращается. Цитоплазма становится более жидкой.
В фазе дифференциации клетки приобретают индивидуальные особенности. Специализация клеток происходит в соответствии с их расположением в растении. Обязательный момент дифференциации – образование вторичной оболочки. Характерные морфологические особенности приобретает протопласт.
Выполнившие свои физиологические функции клетки отмирают. Толстостенные клетки и после отмирания продолжают функционировать как проводящие или механические ткани. Тонкостенные после отмирания протопласта сплющиваются под давлением соседних клеток и постепенно рассасываются. Этот процесс называется облитерация (лат. obliteratus – прекращение).
Известно, что соседние клетки как бы сцементированы срединной пластинкой. Поэтому, когда она разрушается при помощи некоторых веществ, то оболочки соседних клеток отклеиваются друг от друга, и клетки разъединяются. Процесс разъединения клеток в результате разрушения срединной пластинки называют мацерацией. Впервые метод мацерации для изучения строения растительных клеток применил немецкий ботаник Мольденгауэр (1812). Естественная мацерация происходит, когда пектиновые вещества срединной пластинки при помощи пектиназы переходят в растворимое состояние и потом вымываются водой. Наблюдается естественная мацерация в переспевших плодах груши, дыни, персика. Естественная мацерация происходит в черешках листьев перед листопадом, в плодоножках и лепестках перед их опаданием. Мацерация наглядно показывает, что каждая клетка имеет свою собственную оболочку.
При переходе клеток из эмбрионального во взрослое состояние наблюдается частичная мацерация клеток, когда срединная пластинка разрушается по углам клеток. В результате тургорного давления соседние клетки в этих местах округляются, образуются межклетники. Межклетники такого типа называют схизогенными, образующиеся путем расщепления. Некоторые межклетники возникают путем растворения, распада целых клеток. Это лизигенные межклетники.
Межклетники обоих типов могут служить вместилищем разнообразных продуктов секреции.
Список литературы
1. Бавтуто Г.А., Еремин В.М. Ботаника: Морфология и анатомия растений. Мн., 1997. – 375 с.
2. Васильев А.Е. и др. Ботаника: Морфология и анатомия растений. М., 1988. – 480 с.
3. Грин Н., Стаут У., Тейлор Д. Биология. В 3-х т. М., 1990.
4. Лемеза Н.А., Лисов Н.Д. Клетка – основа жизни. Мн., 1997. – 168 с.
5. Хржановский В.Г. Курс общей ботаники. М., 1976. – 272 с.
6. Яковлев Г.П., Челомбитько В.А. Ботаника. М., 1990. – 367 с.
Похожие работы
... у взрослого организма остается объектом научных дискуссий, хотя исследования показали, что существует возможность образования плюрипотентных клеток из фибробластов взрослого человека. Ввиду пластичности и потенциально неограниченного потенциала самообновления, эмбриональные стволовые клетки имеют перспективы применения в регенеративной медицине и замещении поврежденных тканей. Однако в настоящий ...
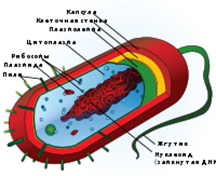
... организма, равнозначны по генетической информации, но отличаются друг от друга разной работой различных генов, что приводит к их морфологическому и функциональному разнообразию - к дифференцировке. История развития понятий о клетке XVII век 1665 год — английский физик Р. Гук в работе «Микрография» описывает строение пробки, на тонких срезах которой он нашёл правильно расположенные пустоты. ...
... (наличие в корме белков, углеводов, липидов, минеральных солей, витаминов, микроэлементов). Важны также кислород, температура, свет (синтез витамина Д). Рост и индивидуальное развитие животных организмов подвержены нейрогуморальной регуляции со стороны гуморальных и нервных механизмов регуляции. У растений обнаружены гормоноподобные активные вещества, получившие название фитогормонов. Последние ...
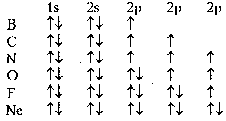
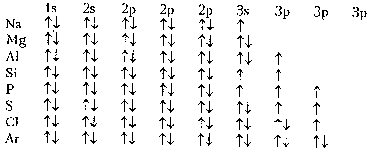
... о биологической причинности. Ряд феноменов, которые витализм считал специфическими для биологических объектов (способность к саморегуляции, усложнение строения, достижение одного результата разными способами) рассматриваются в современном естествознании как типичные проявления процессов самоорганизации любых достаточно сложных систем, а не только живых. Н.Бор: “ни один результат биологического ...
0 комментариев