Огловление
Форма тела и способы движения
Кожные покровы рыб
Мышечная система и электрические органы
Пищеварительная система
Дыхательная система и газообмен (New)
Кровеносная система
Нервная система и органы чувств
Железы внутренней секреции
Ядоносность и ядовитость рыб
Форма тела рыб и способы движения рыб
Форма тела должна обеспечивать рыбе возможность передвигаться в воде (среде, значительно более плотной, чем воздух) с наименьшей затратой энергии и со скоростью, соответствующей ее жизненным потребностям.
Форма тела, отвечающая этим требованиям, выработалась у рыб в результате эволюции: гладкое, без выступов тело, покрытое слизью, облегчает движение; шеи нет; заостренная голова с прижатыми жаберными крышками и сжатыми челюстями рассекает воду; система плавников определяет движение в нужном направлении. В соответствии с образом жизни выделено до 12 различных типов формы тела

Рис. 1 - сарган; 2 - скумбрия; 3 - лещ; 4 - рыба-луна; 5 - камбала; 6 - угорь; 7 - рыба-игла; 8 - сельдяной король; 9 - скат; 10 - рыба-еж; 11 - кузовок; 12 - макрурус.
Стреловидная - кости рыла вытянуты и заострены, тело рыбы по всей длине имеет одинаковую высоту, спинной плавник отнесен к хвостовому и располагается над анальным, чем создается имитация оперения стрелы. Эта форма типична для рыб, не перемещающихся на большие расстояния, держащихся в засаде и развивающих высокие скорости движения на короткий промежуток времени за счет толчка плавников при броске на добычу или уходе от хищника. Это щуки (Esox), сарганы (Belone) и др. Торпедовидная (ее часто называют веретеновидной) - характеризуется заостренной головой, закругленным, имеющим в поперечном разрезе форму овала телом, утонченным хвостовым стеблем, нередко с дополнительными плавничками. Она свойственна хорошим пловцам, способным к продолжительным перемещениям - тунцам, лососям, скумбриям, акулам и др. Эти рыбы способны в течение длительного времени плыть, если так можно выразиться, с крейсерской скоростью 18 км в час. Лососи способны совершать двух-трех метровые прыжки при преодолении препятствий во время нерестовых миграций. Максимальная скорость, которую может развить рыба, равна 100-130 км в час. Этот рекорд принадлежит рыбе-паруснику. Симметрично сжатое с боков тело - сильно сжато с боков, высокое при относительно небольшой длине и высокое. Это рыбы коралловых рифов - щетинкозубы (Chaetodon), зарослей донной растительности - скалярии (Pterophyllum). Такая форма тела помогает им легко маневрировать среди препятствий. Симметрично сжатую с боков форму тела имеют и некоторые пелагические рыбы, которым необходимо быстро менять положение в пространстве для дезориентации хищников. Такую же форму тела имеют рыба-луна (Mola mola L.) и лещ (Abramis brama L.). Несимметрично сжатое с боков тело - глаза смещены на одну сторону, что создает асимметрию тела. Она свойственна придонным малоподвижным рыбам отряда Камбалообразные, помогая им хорошо маскироваться на дне. В движении этих рыб большую роль играют волнообразные изгибания длинных спинного и анального плавников. Уплощенное в дорзовентральном направлении тело - сильно сжато в спинно-брюшном направлении, как правило, хорошо развиты грудные плавники. Такую форму тела имеют малоподвижные донные рыбы - большинство скатов (Batomorpha), морской черт (Lophius piscatorius L.). Уплощенное тело маскирует рыб в условиях дна, а расположенные сверху глаза помогают видеть добычу. Угревидная форма - тело рыб удлиненное, закругленное, имеющее вид овала на поперечном разрезе. Спинной и анальный плавники длинные, брюшных плавников нет, а хвостовой плавник небольшой. Она характерна для таких донных и придонных рыб, как угреобразные (Anguilliformes), передвигающихся, латерально изгибая тело. Лентовидная - тело рыб удлиненное, но в отличие от угревидной формы сильно сжато с боков, что обеспечивает большую удельную поверхность и позволяет рыбам обитать в толще воды. Характер движения у них такой же, как и у рыб угревидной формы. Такая форма тела характерна для рыбы-сабли (Trichiuridae), сельдяного короля (Regalecus). Макруровидная - тело рыбы высокое в передней части, суженное с задней, особенно в хвостовом отделе. Голова крупная, массивная, глаза большие. Свойственна глубоководным малоподвижным рыбам- макрурусообразным (Macrurus), химерообразным (Chimaeriformes). Астеролепидная (или кузовковидная) - тело заключено в костный панцирь, что обеспечивает защиту от хищников. Эта форма тела характерна для придонных обитателей, многие из которых встречаются в коралловых рифах, например для кузовков (Ostracion). Шаровидная форма свойственна некоторым видам из отряда Иглобрюхообразные (Tetraodontiformes) - рыбе-шару (Sphaeroides), рыбе-ежу (Diodon) и др. Эти рыбы плохие пловцы и передвигаются с помощью ундулирующих (волнообразных) движений плавников на небольшие расстояния. При опасности рыбы раздувают воздушные мешки кишечника, наполняя их водой или воздухом; при этом расправляются имеющиеся на теле шипы и колючки, защищающие их от хищников. Игловидная форма тела характерна для морских игл (Syngnathus). Их удлиненное, скрытое в костном панцире тело имитирует листья зостеры, в зарослях которой они обитают. Рыбы лишены боковой подвижности и перемещаются с помощью ундулирующего (волнообразного) действия спинного плавника.
Нередко встречаются рыбы, форма тела которых напоминает одновременно различные типы форм. Для ликвидации демаскирующей тени на брюхе рыбы, возникающей при освещении сверху, мелкие пелагические рыбы, например сельдевые (Clupeidae), чехонь (Pelecus cultratus (L.)], имеют заостренное, сжатое с боков брюшко с острым килем . У крупных подвижных пелагических хищников-скумбрий (Scomber), рыбы-меча (Xiphias gladius L.), тунцов (Thunnus)- киль обычно не развивается. Их способ защиты состоит в быстроте движения, а не в маскировке. У придонных рыб форма поперечного сечения приближается к равнобедренной трапеции, обращенной большим основанием вниз, что исключает появление тени на боках при освещении сверху. Поэтому большинство придонных рыб имеют широкое уплощенное тело.
КОЖА, ЧЕШУЯ И ОРГАНЫ СВЕЧЕНИЯ
В кожном покрове рыб различают два слоя: наружный слой эпителиальных клеток, или эпидермис, и внутренний слой из соединительнотканных клеток - собственно кожа, дерма, кориум, кутис.
Эпидермис защищает организм от проникновения в него веществ извне. Он состоит из многослойного эпителия, форма клеток и количество слоев которого варьируют у разных рыб. Но как правило самые верхние слои клеток плоские, по мере углубления они по форме приближаются к цилиндрическим. Эти клетки имеют ядро и способны размножаться. Новые слои клеток, отходящие от цилиндрических кнаружи, постепенно заменяют верхний отмирающий слой. Самый наружный слой эпителиальных клеток ороговевает, но в отличие от наземных позвоночных у рыб он не отмирает, сохраняя связь с живыми клетками. В течение жизни рыбы итенсивность ороговения эпидермиса не остается неизменной, наибольшей степени оно достигает у некоторых рыб перед нерестом: так, у самцов карповых и сиговых в некоторых местах тела (особенно на голове, жаберных крышках, боках и др.) появляется так называемая жемчужная сыпь - масса мелких белых бугорков, придающих коже шероховатость. После нереста она исчезает.
Между клетками эпидермиса имеются протоплазматические связи, по которым циркулирует лимфа, обеспечивающая питание клеток. В эпидермисе кожи рыб имеются различные железистые клетки, особенно слизевые. В целом кожа рыб отличается от кожи других позвоночных большим количеством слизи. Слизь образуется в специализированных железистых клетках, лежащих в эпидермисе.
Как правило, рыбы с хорошо развитым чешуйчатым покровом , выделяют слизи меньше (лососевые, окуневые). Рыбы, лишенные чешуи или чешуя которых редуцирована (круглоротые, некоторые сомы, линь, вьюн), выделяют слизи очень много. С бактерицидными свойствами слизи линя связывают повышенную устойчивость его к заражению паразитами, тогда как другие - представители семейства карповых такой устойчивостью не обладают. Роль слизи в жизненных отправлениях рыб очень велика: она уменьшает трение тела о воду (механическая защита), предотвращает попадание в организм паразитов и бактерий (бактерицидная защита), ускоряет свертывание крови в случаях ранений, способствует выведению веществ из организма, регулирует проникновение воды и солей (осмотическая регуляция), осаждает муть, склеивая частицы взвеси, в случае наличия в воде солей тяжелых металлов (медь, цинк, свинец, хром) коагулирующая слизь образует на коже защитный слой и, наконец, выделяет специфический видовой запах. Слизь некоторых рыб ядовита. Слизь миног вызывает у хищников нарушение пищеварения.
У некоторых рыб в эпидермисе имеются ядоотделительные железы, расположенные в основном у основания шипов или колючих лучей плавников. Иногда ядоотделительные клетки образуются и функционируют только во время размножения, в других случаях - постоянно. Уколы таких ядоносных рыб могут быть опасны для человека.
У ската - хвостокола ядоотделительные клетки расположены в эпидермисе у основания хвостового шипа. При уколе яд по желобку шипа поступает в рану и вызывает острую боль, сильный отек, озноб, тошноту и рвоту. В некоторых случаях наблюдается расстройство дыхания и даже смерть.
Особенно ядоносной является тихоокеанская бородавчатка. Многие лучи в ее плавниках снабжены ядоотделительными железами. При прикосновении к рыбе яд по бороздкам колючих лучей попадает в раны. Он разрушает красные кровяные тельца, парализует нервную систему и может оказаться смертельным не только для водных животных, но и для человека.
Ядоносных рыб следует отличать от ядовитых, у которых отдельные части тела (мышцы, кровь, икра, печень, кожа и др.) при употреблении в пищу вызывают отравление. Наибольшее число ядовитых рыб относится к семействам фахаков и кузовков, распространенных в водах Юго-Восточной Азии и тропической части Тихого океана.
Особенно ядовиты собаки-рыбы из отряда сростночелюстных. Наиболее опасным представителем этого отряда является фугу. В печени, икре, молоках и коже фугу содержится сильный яд. Мясо фугу съедобно и вкусно и ценится в некоторых странах. В Японии фугу разводят искусственно.
Среди акуловых и костистых (в большинстве глубоководных) рыб многие обладают способностью к люминесценции. Это свечение особого рода, при котором светоиспускание (в отличие от обычного - возникающего при тепловом излучении - основанного на тепловом возбуждении электронов и потому сопровождающегося выделением тепла) связано с генерацией холодного света (необходимая энергия образуется в результате химической реакции). Некоторые виды генерируют свет сами, другие своим свечением обязаны симбиотическим светящимся бактериям, которые находятся на поверхности тела или в специальных органах. В обоих случаях светящиеся клетки или органы располагаются в эпидермисе, но иногда во внутренних органах, например в кишечнике.
Нижний слой кожи - кориум - представляет собой проходящие в косом направлении волокна соединительной ткани. Здесь же находятся кровеносные сосуды и нервные окончания. В нижних слоях эпидермиса и в пограничных с ним слоях кориума залегают пигментные клетки - хроматофоры. Это звездчатые клетки со множеством отростков, включающие зернышки пигмента. Они определяют все разнообразие окраски рыб, особенно яркой в тропиках. Различие в окраске достигается сочетанием разных хроматофоров: меланофоры имеют зерна черного пигмента, ксантофоры - желтого, эритрофоры - красного; гуанофоры или иридоциты не имеют пигментных зерен, но содержат кристаллики гуанина, благодаря которым рыбы приобретают серебристую окраску.
Интенсивность окраски определяется состоянием хроматофоров: при их расширении пигментные зерна растекаются на большее пространство и окраска тела становится яркой. Если хроматофоры сокращаются, пигментные зерна скапливаются в центре, оставляя большую часть клетки неокрашенной и окраска тела бледнеет.
Форма пигментных клеток постоянна и у близких видов сходна, а состояние меняется в зависимости от различных факторов: температуры и газового режима водоема, возраста, пола, состояния организма (голод, размножение и т. п.), эмоций (возбуждение, страх) рыбы и др.
Сочетания разных цветов хроматофоров и отражательных поверхностей иридоцитов и их различные комбинации делают окраску рыб удивительно разнообразной. Сгущение и расхождение пигментов в клетках, а отсюда и изменение окраски тела рыб происходит под влиянием нервной системы и связано со зрительными восприятиями, причем центр просветления находится в продолговатом, центр потемнения - в промежуточном мозгу.
Изменение окраски в период размножения (появление брачного наряда), часто наблюдаемое в этот период различие в окраске самок и самцов происходят под влиянием гормональных факторов.
Разумеется, главное в расцветке большинства рыб - ее защитные свойства. Покровительственная окраска рыб, обитающих в поверхностных слоях воды, - темная спина и белое или серебристое брюшко - делает их малозаметными. Особенно удивительна маскировка у донных рыб - их окраска соответствует цвету дна. Наиболее удивительным мастером камуфляжа является обыкновенная камбала: она с легкостью хамелеона подделывается под камни, песок, темный ил.
Кожа подстилается рыхлой соединительнотканной прослойкой (подкожная соединительная ткань, подкожная клетчатка). У многих рыб в подкожной клетчатке откладывается жир.
В соединительнотканном слое кожи образуется чешуя. У рыб различают несколько видов чешуи: плакоидная, ганоидная и костная
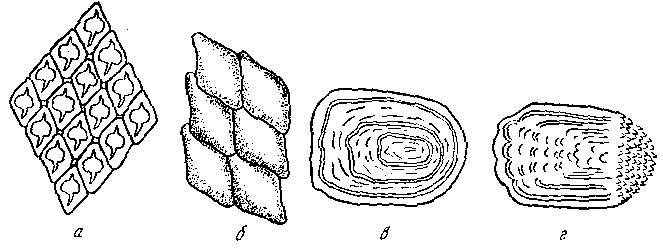
Рис. Форма чешуи рыб. а - плакоидная; б - ганоидная; в - циклоидная; г – ктеноидная
Плакоидная - наиболее древняя, сохранилась у хрящевых рыб (акулы, скаты). Состоит из пластинки, на которой возвышается шипик. Старые чешуйки сбрасываются, на их месте возникают новые. Ганоидная - преимущественно у ископаемых рыб. Чешуйки имеют ромбическую форму, тесно сочленяются одна с другой, так что тело оказывается заключенным в панцирь. Чешуйки со временем не меняются. Названием своим чешуя обязана ганоину (дентинообразному веществу), толстым слоем лежащему на костной пластинке. Среди современных рыб ее имеют панцирные щуки и многоперы. Кроме того, она имеется у осетровых в виде пластинок на верхней лопасти хвостового плавника (фулькры) и жучек, разбросанных по телу (модификация нескольких слившихся ганоидных чешуек).
Постепенно видоизменяясь, чешуя теряла ганоин. У современных костистых рыб его уже нет, а чешуйки состоят из костных пластинок (костная чешуя). Эти чешуйки могут быть циклоидными - округлыми, с гладкими краями (карповые) и ктеноидными с зазубренным задним краем (окуневые). Обе формы родственны, но циклоидная как более примитивная встречатся у низкоорганизованных рыб. Бывают случаи, когда в пределах одного вида самцы имеют ктеноидную, а самки - циклоидную чешую (камбалы рода Liopsetta), или даже у одной особи встречаются чешуйки обеих форм.
Размеры и толщина чешуи у рыб сильно различаются- от микроскопических чешуек обыкновенного угря до очень крупных, величиной с ладонь чешуи трехметрового усача-тора, живущего в индийских реках. Лишь немногие рыбы не имеют чешуи. У некоторых она слилась в сплошной неподвижный панцирь, как у кузовка, или образовала ряды тесно соединенных костных пластинок, как у морских коньков.
Костные чешуйки, как и ганоидные, постоянны, не сменяются и лишь ежегодно увеличиваются в соответствии с ростом рыбы, и на них остаются отчетливые годичные и сезонные метки. Зимний слой имеет более частые и тонкие напластования, чем летний, поэтому он темнее летнего. По числу летних и зимних слоев на чешуе можно определить возраст некоторых рыб.
Под чешуей у многих рыб имеются серебристые кристаллики гуанина. Отмытые от чешуи, они являются ценным веществом для получения искусственного жемчуга. Из чешуи рыб изготовляют клей.
По бокам тела многих рыб можно наблюдать ряд выделяющихся чешуек с отверстиями, которые образуют боковую линию - один из важнейших органов чувств. Количество чешуи в боковой линии -
В одноклеточных железах кожи образуются феромоны - летучие (пахучие) вещества, выделяемые в окружающую среду и воздействующие на рецепторы других рыб. Они специфичны для разных видов, даже близкородственных; в некоторых случаях определена их внутривидовая дифференцировка (возрастная, половая).
У многих рыб, в том числе у карповых, образуется так называемое вещество страха (ихтиоптерин), которое выделяется в воду из тела пораненной особи и воспринимается ее сородичами как сигнал, извещающий об опасности.
Кожа рыб быстро регенерирует. Через нее происходит, с одной стороны, частичное выделение конечных продуктов обмена веществ, а с другой - поглощение некоторых веществ из внешней среды (кислород, угольная кислота, вода, сера, фосфор, кальций и другие элементы, играющие большую роль в жизнедеятельности). Большую роль играет кожа и как рецепторная поверхность: в ней располагаются термо-, баро-, хемо- и другие рецепторы.
В толще кориума образуются покровные кости черепа и пояса грудных плавников.
Через мышечные волокна миомеров, соединенные с ее внутренней поверхностью, кожа участвует в работе туловищно-хвостовой мускулатуры.
Мышечная система и электрические органы
Мышечную систему рыб, как и других позвоночных, разделяют на мышечную систему тела (соматическую) и внутренних органов (висцеральную).
В первой выделяют мускулы туловища, головы и плавников. Внутренние органы имеют свою мускулатуру.
Мышечная система взаимосвязана со скелетом (опора при сокращении) и нервной системой (к каждому мышечному волокну подходит нервное волокно, и каждая мышца иннервируется определенным нервом). Нервы, кровеносные и лимфатические сосуды располагаются в соединительнотканной прослойке мышц, которая в отличие от мышц млекопитающих невелика,
У рыб, как и других позвоночных, сильнее всего развита туловищная мускулатура. Она обеспечивает плавание рыбы. У настоящих рыб она представлена двумя большими тяжами, расположенными вдоль тела от головы до хвоста (большая боковая мышца - m. lateralis magnus) (рис. 1). Продольной соединительнотканной прослойкой эта мышца делится на спинную (верхнюю) и брюшную (нижнюю) части.
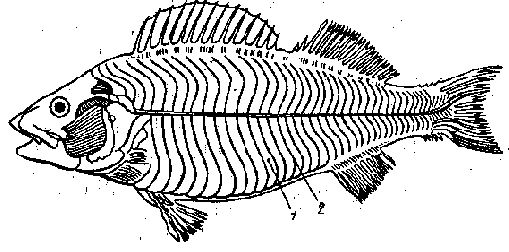
Рис. 1 Мускулатура костистой рыбы (по Кузнецову, Чернову, 1972):
1 - миомеры, 2 - миосепты
Боковые мышцы разделены миосептами на миомеры, число которых соответствует количеству позвонков. Наиболее отчетливо миомеры видны у личинок рыб, пока их тела прозрачны.
Мышцы правой и левой сторон, поочередно сокращаясь, сгибают хвостовой отдел тела и изменяют положение хвостового плавника, благодаря чему тело двигается вперед.
Над большой боковой мышцей вдоль тела между плечевым поясом и хвостом у осетровых и костистых лежит прямая боковая поверхностная мышца (m. rectus lateralis, m. lateralis superficialis). У лососевых в ней откладывается много жира. По нижней стороне тела тянется прямая брюшная мышца (m. rectus abdominalis); у некоторых рыб, например угрей, ее нет. Между ней и прямой боковой поверхностной мышцей располагаются косые мышцы (m. obliguus).
Группы мышц головы управляют движениями челюстного и жаберного аппаратов (висцеральная мускулатура), Плавники имеют свою мускулатуру.
Наибольшее скопление мускулов определяет и расположение центра тяжести тела: у большинства рыб он находится в спинной части.
Деятельность туловищных мышц регулируется спинным мозгом и мозжечком, а висцеральная мускулатура иннервируется периферической нервной системой, возбуждаемой непроизвольно.
Различают поперечнополосатые (действующие в значительной степени произвольно) и гладкие мышцы (которые действуют независимо от воли животного). К поперечно-полосатым относятся скелетные мышцы тела (туловищные) и мышцы сердца. Туловищные мышцы могут быстро и сильно сокращаться, однако скоро утомляются. Особенностью строения сердечных мышц является не параллельное расположение обособленных волокон, а разветвление их кончиков и переход из одного пучка в другой, что обусловливает непрерывную работу этого органа.
Гладкие мышцы также состоят из волокон, но гораздо более коротких и не обнаруживающих поперечной исчерченности. Это мышцы внутренних органов и стенок кровеносных сосудов, имеющие периферическую (симпатическую) иннервацию.
Поперечнополосатые волокна, а следовательно, и мышцы делят на красные и белые, различающиеся, как следует из названия, цветом. Цвет обусловлен наличием миоглобина - белка, легко связывающего кислород. Миоглобин обеспечивает дыхательное фосфорилирование, сопровождающееся выделением большого количества энергии.
Красные и белые волокна различны по целому ряду морфофизиологических характеристик: цвету, форме, механическим и биохимическим свойствам (интенсивность дыхания, содержание гликогена и т. д.).
Волокна красной мышцы (m. lateralis superficialis) - узкие, тонкие, интенсивно кровоснабжаемые, расположенные более поверхностно (у большинства видов под кожей, вдоль тела от головы до хвоста), содержат в саркоплазме больше миоглобина;
в них обнаружены скопления жира и гликогена. Возбудимость их меньше, отдельные сокращения длятся дольше, но протекают медленней; окислительный, фосфорный и углеводный обмен интенсивнее, чем в белых.
В мышце сердца (красной) мало гликогена и много ферментов аэробного обмена (окислительный обмен). Она характеризуется умеренной скоростью сокращений и утомляется медленнее, чем белые мышцы.
В широких, более толстых, светлых белых волокнах m. lateralis magnus миоглобина мало, меньше в них гликогена и дыхательных ферментов. Углеводный обмен происходит преимущественно анаэробно, и количество выделяемой энергии меньше. Отдельные сокращения быстры. Мышцы быстрее сокращаются и утомляются, чем красные. Лежат они более глубоко.
Красные мышцы постоянно деятельны. Они обеспечивают длительную и непрерывную работу органов, поддерживают постоянное движение грудных плавников, обеспечивают изгибы тела при плавании и поворотах, непрерывную работу сердца.
При быстром движении, бросках активны белые мышцы, при медленном - красные. Поэтому наличие красных или белых волокон (мышц) зависит от подвижности рыб: "спринтеры" обладают почти исключительно белыми мышцами, у рыб, которым свойственны продолжительные миграции, кроме красных Боковых мышц имеются добавочные красные волокна в белых мышцах.
Основную массу мышечной ткани у рыб составляют белые мышцы. Например, у жереха, плотвы, чехони на их долю приходится 96,3; 95,2 и 94,9% соответственно.
Белые и красные мышцы различаются по химическому составу. В красных мышцах содержится больше жира, тогда как в белых мышцах больше влаги и белка.
Толщина (диаметр) мышечного волокна изменяется в зависимости от вида рыб, их возраста, величины, образа жизни, а у прудовых рыб - от условий содержания. Например, у карпа, выращенного на естественной пище, диаметр мышечного волокна составляет (мкм): у мальков - 5 ... 19, сеголетков - 14 ... 41, двухлетков - 25 ... 50.
Туловищная мускулатура образует основную долю мяса рыбы. Выход мяса в процентах общей массы тела (мясистость) неодинаков у разных видов, а у особей одного вида различается в зависимости от пола, условий содержания и др.
Мясо рыб усваивается быстрее, чем мясо теплокровных животных. Оно чаще бесцветно (судак) или имеет оттенки (оранжевый - у лососевых, желтоватый у осетровых и др.) в зависимости от наличия различных жиров и каротиноидов.
Основную массу белков мышц рыб составляют альбумины и глобулины (85%), всего же у разных рыб выделяют 4 ... 7 фракций белков.
Химический состав мяса (вода, жиры, белки, минеральные вещества) различен не только у разных видов, но и в разных частях тела. У рыб одного вида количество и химический состав мяса зависят от условий питания и физиологического состояния рыбы.
В нерестовый период, особенно у проходных рыб, расходуются резервные вещества, наблюдается истощение и, как следствие, уменьшается количество жира и ухудшается качество мяса. У кеты, например, во время подхода к нерестилищам относительная масса костей увеличивается в 1,5 раза, кожи - в 2,5 раза. Мышцы оводняются - содержание сухого вещества снижается более чем в два раза; из мышц практически исчезают жир и азотистые вещества - рыба теряет до 98,4% жира и 57% белка.
Особенности окружающей среды (в первую очередь пищи и воды) могут сильно изменять пищевую ценность рыбы: в заболоченных, тинистых или загрязненных нефтепродуктами водоемах рыбы имеют мясо с неприятным запахом. Качество мяса зависит и от диаметра мышечного волокна, а также количества жира в мышцах. В значительной мере оно определяется соотношением массы мышечной и соединительной тканей, по которому можно судить о содержании в мышцах полноценных мышечных белков (по сравнению с неполноценными белками соединительнотканной прослойки). Это соотношение изменяется в зависимости от физиологического состояния рыбы и факторов внешней среды. В мышечных белках костистых рыб на белки приходится: саркоплазмы 20 ... 30%, миофибрилл - 60 ... 70, стромы - около 2%.
Все многообразие движений тела обеспечивает работа мышечной системы. Она главным образом обеспечивает и выделение тепла и электричества в организме рыбы. Электрический ток образуется при проведении нервного импульса по нерву, при сокращении миофибрилл, раздражении светочувствительных клеток, механохеморецепторов и др.
Электрические органы
Своеобразно измененными мышцами являются электрические органы. Эти органы развиваются из зачатков поперечно-полосатой мускулатуры и расположены по бокам тела рыб. Они состоят из множества мышечных пластинок (у электрического угря их около 6000), преобразованных в электрические пластинки (электроциты), переслаиваемые студенистой соединительной тканью. Нижняя часть пластинки заряжена отрицательно, верхняя - положительно. Разряды происходят под действием импульсов продолговатого мозга. Вследствие разрядов вода разлагается на водород и кислород, поэтому, например, в заморных водоемах тропиков около электрических рыб скапливаются мелкие обитатели - моллюски, рачки, привлеченные более благоприятными условиями дыхания.
Электрические органы могут располагаться в разных частях тела: например, у ската морской лисицы - на хвосте, у электрического сома - на боках.
Генерируя электрический ток и воспринимая силовые линии,
искаженные встречающимися на пути предметами, рыбы ориентируются в потоке, обнаруживают препятствия или добычу с расстояния нескольких метров даже в мутной воде.
В соответствии со способностью к генерации электрических полей рыб разделяют на три группы:
1. Сильно электрические виды - имеют большие электрические органы, генерирующие разряды от 20 до 600 и даже 1000 В. Основное назначение разрядов - нападение и оборона (электрический угорь, электрический скат, электрический сом).
2. Слабоэлектрические виды - имеют небольшие электрические органы, генерирующие разряды напряжением менее 17 В. Основное назначение разрядов - локация, сигнализация, ориентация (обитающие в мутных реках Африки многие мормириды, гимнотиды, некоторые скаты).
3. Неэлектрические виды - не имеют специализированных органов, но обладают электрической активностью. Генерируемые ими разряды распространяются на 10 ... 15 м в морской воде и до 2 м в пресной воде. Основное назначение генерируемого электричества - локация, ориентация, сигнализация (многие морские и пресноводные рыбы: например, ставрида, атерина, окунь и др.).
Пищеварительная система
В пищеварительном тракте настоящих рыб различают ротовую полость, глотку, пищевод, желудок, кишечник (тонкая, толстая, прямая кишка, заканчивающаяся анусом). У акул, скатов и некоторых других рыб перед анусом имеется клоака — расширение, куда изливается прямая кишка и протоки мочевой и половой систем.
В ротовой полости рыб нет слюнных желез. Железистые клетки ротовой полости и глотки выделяют слизь, которая не имеет пищеварительных ферментов и способствует лишь проглатыванию пищи, а также защищает эпителий ротовой полости с вкрапленными вкусовыми почками (рецепторами).
Мощный и выдвижной язык имеют только круглоротые, у костистых рыб он не обладает собственной мускулатурой.
Рот обычно снабжен зубами. Наличием эмалевого колпачка и слоев дентина они напоминают зубы высших позвоночных. У хищников они располагаются как на челюстях, так и на других костях полости рта, иногда даже на языке; они острые. часто крючкообразные, наклонены внутрь к глотке и служат для схватывания и удержания жертвы. У многих мирных рыб (многие сельдевые, карповые и др.) на челюстях зубов нет.
Механизм питания координирован с дыхательным механизмом. Вода, всасываемая в рот при вдохе, несет и мелкие планктонные организмы, которые при выталкивании воды из жаберной полости (выдох) задерживаются в ней жаберными тычинками.
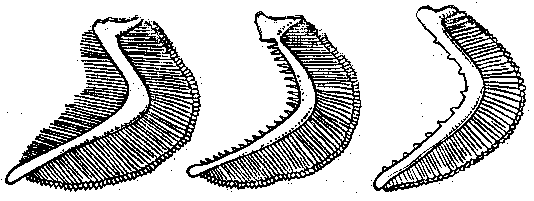
Рис. 1 Жаберные тычинки планктоноядной (а), бентосоядной (б), хищной (в) рыб.
Они так тонки, длинны и многочисленны у рыб, питающихся планктоном (планктонофагов), что образуют фильтрующий аппарат. Отцеженный комочек пищи направляется в пищевод. Хищные рыбы не нуждаются в отфильтровывании пищи, тычинки у них редкие, низкие, грубые, острые или крючковатые: они участвуют в удержании жертвы.
У некоторых бентосоядных рыб на задней жаберной дуге имеются широкие и массивные глоточные зубы, Они служат для перетирания пищи.
Следующий за глоткой пищевод, обычно короткий, широкий и прямой с сильными мускулистыми стенками проводит пищу в желудок. В стенках пищевода имеются многочисленные клетки, выделяющие слизь. У открытопузырных рыб в пищевод открывается проток плавательного пузыря.
Не все рыбы имеют желудок. К безжелудочным относятся карповые, многие бычки и некоторые другие.
В слизистой оболочке желудка имеются железистые клетки. вырабатывающие соляную кислоту и пепсин, расщепляющий белок в кислой среде, и слизь. Здесь у хищных рыб переваривается основная часть пищи.
В начальную часть кишечника (тонкую кишку) впадают желчный проток и проток поджелудочной железы. По ним в кишечник попадают желчь и ферменты поджелудочной железы, под действием которых происходит расщепление белков до аминокислот, жиров до глицерина и жирных кислот и расщепление полисахаридов до сахаров, главным образом глюкозы.
В кишечнике помимо расщепления питательных веществ происходит их всасывание, наиболее интенсивно протекающее в заднем участке. Этому способствует складчатое строение его стенок, наличие в них ворсинкообразных выростов, пронизанных капиллярами и лимфатическими сосудами, наличие клеток, секретирующих слизь.
У многих видов в начальной части кишечника помещаются слепые отростки — пилорические придатки, число которых сильно варьирует: от 3 у окуня до 400 у лососей
Карповые, сомовые, щуки и некоторые другие рыбы пилорических придатков не имеют. При помощи пилорических придатков всасывающая поверхность кишечника увеличивается в несколько раз.
У рыб, не имеющих желудка, кишечный тракт представляет собой большей частью недифференцированную трубку, суживающуюся к концу. У некоторых рыб, в частности у карпа, передняя часть кишечника расширена и напоминает по форме желудок. Однако это лишь внешняя аналогия: здесь нет характерных для желудка желез, вырабатывающих пепсин.
Строение, форма и длина пищеварительного тракта разнообразны в связи с характером пищи (объектами питания, их усвояемостью), особенностями переваривания. Наблюдается определенная зависимость длины пищеварительного тракта от рода пищи. Так, относительная длина кишечника (отношение длины кишечника к длине тела.) составляет у растительноядных (пинагора и толстолобика)—б... 15, у всеядных (карася и карпа) — 2 ... 3, у хищных (щуки, судака, окуня) — 0,6 ... 1,2.
Печень — крупная пищеварительная железа, по размерам уступающая у взрослых рыб только гонадам. Ее масса составляет у акул 14... 25%, у костистых — 1 ... 8% массы тела Это сложная трубчато-сетчатая железа, по происхождению связанная с кишечником. У зародышей является его слепым выростом.
Желчные протоки проводят желчь в желчный пузырь (только у единичных видов его нет). Желчь благодаря щелочной реакции нейтрализует кислую реакцию желудочного сока. Она эмульгирует жиры, активирует липазу-—фермент поджелудочной железы.
Из пищеварительного тракта вся кровь медленно протекает через печень. В печеночных клетках кроме образования желчи происходит обезвреживание попавших с пищей чужеродных белков и ядов, откладывается гликоген, а у акул и тресковых (треска, налим и др.). — жир и витамины. Пройдя через печень, кровь по печеночной вене направляется к сердцу.
Барьерная функция печени (очищение крови от вредных веществ) обусловливает ее важнейшую роль не только в пищеварении, но и кровообращении.
Поджелудочная железа — сложная альвеолярная железа, также производная кишечника, является компактным органом только у акул и немногих других рыб. У большинства рыб она визуально не обнаруживается, так как диффузно внедрена в ткань печени (большей частью), и поэтому ее можно различить только на гистологических препаратах. Каждая долька связана с артерией, веной, нервным окончанием и протоком, выводящим секрет к желчному пузырю. Обе железы носят общее название hepatopancreas.
В поджелудочной железе вырабатываются пищеварительные ферменты, действующие на белки, жиры и углеводы (трипсин, эрепсин, энтерококиназа, липаза, амилаза, мальтаза), которые выводятся в кишечник.
У костистых рыб (впервые среди позвоночных) встречаются в паренхиме поджелудочной железы островки Лангерганса, в которых многочисленны клетки, синтезирующие инсулин, выделяемый прямо в кровь и регулирующий углеводный обмен.
Таким образом, поджелудочная железа является железой внешней и внутренней секреции.
Из мешкообразного впячивания спинной части начала кишечника образуется у рыб плавательный пузырь — орган, свойственный только рыбам.
ДЫХАТЕЛЬНАЯ СИСТЕМА И ГАЗООБМЕН
Эволюция рыб привела к появлению жаберного аппарата, увеличению дыхательной поверхности жабр, а отклонение от основной линии развития — к выработке приспособлений для использования кислорода воздуха. Большинство рыб дышит растворенным в воде кислородом, но есть виды, приспособившиеся частично и к воздушному дыханию (двоякодышащие, прыгун, змееголов и др.).
Основные органы дыхания. Основным органом извлечения кислорода из воды являются жабры.
Форма жабр разнообразна и зависит от видовой принадлежности и подвижности: мешочки со складочками (у рыбообразных), пластинки, лепестки, пучки слизистой, имеющие богатую сеть капилляров. Все эти приспособления направлены на создание наибольшей поверхности при наименьшем объеме.
У костистых рыб жаберный аппарат состоит из пяти жаберных дуг, располагающихся в жаберной полости и прикрытых жаберной крышкой. Четыре дуги на внешней выпуклой стороне имеют по два ряда жаберных лепестков, поддерживаемых опорными хрящами. Жаберные лепестки покрыты тонкими складками — лепесточками. В них и происходит газообмен. Число лепестков варьирует; на 1 мм жаберного лепестка их приходится:
у щуки — 15, камбалы — 28, окуня — 36. В результате полезная дыхательная поверхность жабр очень велика. К основанию жаберных лепестков подходит приносящая жаберная артерия, ее капилляры пронизывают лепесточки; из них окисленная (артериальная) кровь по выносящей жаберной артерии попадает в корень аорты. В капиллярах кровь течет в направлении, противоположном току воды
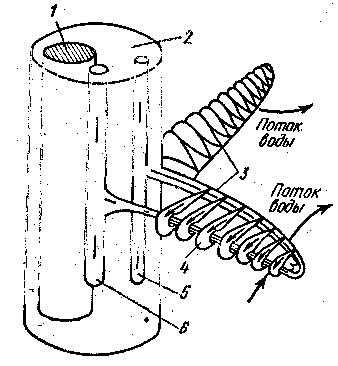
Рис.1 Схема противотока крови и воды в жабрах рыб:
1 - хрящевой стержень; 2 - жаберная дуга; 3 - жаберные лепестки; 4 - жаберные пластинки; 5 - приносящая артерия от брюшной аорты; 6 - выносящая артерия к спинной аорте.
Более активные рыбы имеют большую поверхность жабр: у окуня она почти в 2,5 раза больше, чем у камбалы. Противоток крови в капиллярах и омывающей жабры воды обеспечивает полное насыщение крови кислородом. При вдохе рот открывается, жаберные дуги отходят в стороны, жаберные крышки наружным давлением плотно прижимаются к голове и закрывают жаберные щели. Вследствие уменьшения давления вода всасывается в жаберную полость, омывая жаберные лепестки. При выдохе рот закрывается, жаберные дуги и жаберные крышки сближаются, давление в жаберной полости увеличивается, жаберные щели открываются и вода выталкивается через них наружу.
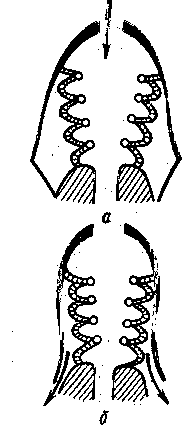
Рис. 2 Механизм дыхания взрослой рыбы
При плавании рыбы ток воды может создаваться за счет движения с открытым ртом. Таким образом, жабры расположены как бы между двумя насосами — ротовым (связанным с ротовыми мышцами) и жаберным (связанным с движением жаберной крышки), работа которых создает прокачивание воды и вентиляцию жабр. За сутки через жабры прокачивается не меньше 1 м3 воды на 1 кг массы тела.
В капиллярах жаберных лепесточков из воды поглощается кислород (он связывается гемоглобином крови) и выделяются двуокись углерода, аммиак, мочевина.
Большую роль играют жабры и в водносолевом обмене, регулируя поглощение или выделение воды и солей. Жаберный аппарат чутко реагирует на состав воды: такие токсиканты, как аммиак, нитриты, СО2 при повышенном содержании поражают респираторные складки в первые же 4 ч контакта.
Замечательны приспособления для дыхания у рыб в эмбриональный период развития — у зародышей и личинок, когда жаберный аппарат еще не сформирован, а кровеносная система уже функционирует. В это время органами дыхания служат:
а) поверхность тела и система кровеносных сосудов — Кювьеровы протоки, вены спинного и хвостового плавников, подкишечная вена, сеть капилляров на желточном мешке, голова, плавниковой кайме и жаберной крышке; б) наружные жабры
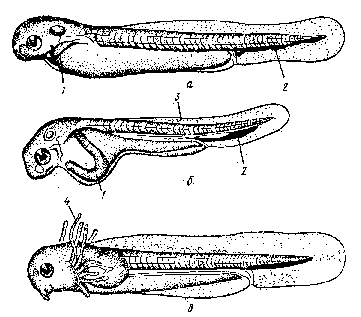
Рис. 3 Органы дыхания у эмбрионов рыб
а - пелагическая рыба; б - карп; в - вьюн; 1 - Кювьеровы протоки; 2 - нижняя хвостовая вена; 3 - сеть капилляров; 4 - наружные жабры.
Это временные, специфические личиночные образования, исчезающие после образования дефинитивных органов дыхания. Чем хуже условия дыхания эмбрионов и личинок, тем сильнее развивается кровеносная система или наружные жабры. Поэтому у рыб, близких в систематическом отношении, но различающихся экологией нереста, степень развития личиночных органов дыхания различна.
Дополнительные органы дыхания. К дополнительным приспособлениям, помогающим переносить неблагоприятные кислородные условия, относятся водное кожное дыхание, т. е. использование растворенного в воде кислорода при помощи кожи, и воздушное дыхание—использование воздуха при помощи плавательного пузыря, кишечника или через специальные добавочные органы
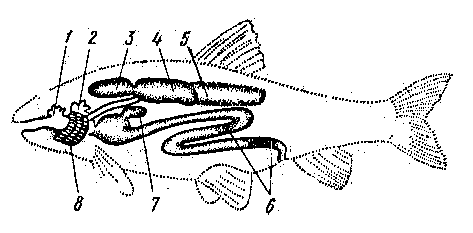
| Рис. 4. Органы водного и воздушного дыхания у взрослых рыб: 1 - выпячивание ротовой полости; 2 - наджаберный орган; 3, 4, 5 - отделы плавательного пузыря; 6 - участки поглощения кислорода в кишечнике; 7 - выпячивание в желудке; 8 - жабры. |
Дыхание через кожу тела — одна из характерных особенностей водных животных. И хотя у рыб чешуя затрудняет дыхание поверхностью тела, у многих видов роль так называемого кожного дыхания велика, особенно в неблагоприятных условиях. По интенсивности такого дыхания пресноводных рыб делят на три группы:
1. Рыбы, приспособившиеся жить в условиях сильного дефицита кислорода. Это рыбы, населяющие хорошо прогреваемые, с повышенным содержанием органических веществ водоемы, в которых часто наблюдается недостаток кислорода. У этих рыб доля кожного дыхания в общем дыхании составляет 17... 22%, у отдельных особей—42... 80%. Это карп, карась, сом, угорь, вьюн. При этом рыбы, у которых кожа имеет наибольшее значение в дыхании, лишены чешуи или она мелкая и не образует сплошного покрова. Например, у вьюна 63% кислорода поглощается кожей, 37%—жабрами; при выключении жабр через кожу потребляется до 85% кислорода, а остальная часть поступает через кишечник.
2. Рыбы, испытывающие меньший недостаток кислорода и попадающие в неблагоприятные условия реже. К ним относятся обитающие у дна, но в проточной воде, осетровые — стерлядь, осетр, севрюга. Интенсивность кожного дыхания у них составляет 9... 12%.
3. Рыбы, не попадающие в условия дефицита кислорода, живущие в проточных или непроточных, но чистых, богатых кислородом водах. Интенсивность кожного дыхания не превышает 3,3 ... 9%. Это сиги, корюшка, окунь, ерш.
Через кожу происходит также выделение углекислоты. Так, у вьюна этим путем выделяется до 92% общего количества.
В извлечении кислорода из воздуха во влажной атмосфере участвует не только поверхность тела, но и жабры. Важное значение при этом имеет температура.
Наибольшей выживаемостью во влажной среде отличаются карась (11 сут), линь (7 сут), сазан (2 сут), в то же время лещ, красноперка, уклея могут жить без воды всего несколько часов и то при низкой температуре.
При перевозке живой рыбы без воды кожное дыхание почти целиком обеспечивает потребность организма в кислороде.
У некоторых рыб, живущих в неблагоприятных условиях, выработались приспособления для дыхания кислородом воздуха. Например, дыхание при помощи кишечника. В стенках кишечника образуются скопления капилляров. Воздух, заглатываемый ртом, проходит через кишечник, и в этих местах кровь поглощает кислород и выделяет двуокись углерода, при этом из воздуха поглощается до 50% кислорода. Такой вид дыхания свойствен вьюновым, некоторым сомовым и карповым рыбам; значение его у разных рыб неодинаково. Например, у вьюнов условиях большого недостатка кислорода именно этот способ дыхания становится почти равным жаберному.
При заморах рыбы заглатывают ртом воздух; воздух аэрирует находящуюся в ротовой полости воду, которая проходит затем через жабры.
Другим способом использования атмосферного воздуха служит образование специальных добавочных органов: например, лабиринтового у лабиринтовых рыб, наджаберного у змееголова и др.
Лабиринтовые рыбы имеют лабиринт—расширенный карманообразный участок жаберной полости, складчатые стенки которого пронизаны густой сетью капилляров, в которых происходит газообмен. Таким способом рыбы дышат кислородом атмосферы и могут находиться вне воды в течение нескольких дней (тропический окунь-ползун Anabas sp. выходит из воды и лазит по камням и деревьям).
У тропических илистых прыгунов (Periophthalmus sp.) жабры окружены губкообразной тканью, пропитанной водой. При выходе этих рыб на сушу жаберные крышки плотно закрываются и предохраняют жабры от высыхания. У змееголова выпячивание глотки образует наджаберную полость, слизистая оболочка ее стенок снабжена густой сетью капилляров. Благодаря наличию наджаберного органа он дышит воздухом и может находиться на мелководье при 30 °С. Для нормальной жизнедеятельности змееголову, как и ползуну, нужен и растворенный в воде кислород, и атмосферный. Однако во время зимовки в прудах, покрытых льдом, он атмосферным воздухом не пользуется.
Для использования кислорода воздуха предназначен и плавательный пузырь. Наибольшего развития как орган дыхания он достигает у двоякодышащих рыб. У них он ячеистый и функционирует как легкое. При этом возникает “легочный круг” кровообращения,
Состав газов в плавательном пузыре определяется как содержанием их в водоеме, так и состоянием рыбы.
Подвижные и хищные рыбы имеют большой запас кислорода в плавательном пузыре, который расходуется организмом при бросках за добычей, когда поступление кислорода через органы дыхания оказывается недостаточным. В неблагоприятных кислородных условиях воздух плавательного пузыря у многих рыб используется для дыхания. Вьюн и угорь могут в течение нескольких дней жить вне воды при условии сохранения влажности кожи и жабр: если в воде жабры обеспечивают угрю 85... 90% общего поглощения кислорода, то в воздухе—только треть. Вне воды угорь использует для дыхания кислород плавательного пузыря и воздух, проходящий через кожу и жабры. Это позволяет ему даже переползать из одного водоема в другой. Карп и сазан, которые не имеют каких-либо специальных приспособлений для использования атмосферного воздуха, при нахождении вне воды частично поглощают кислород из плавательного пузыря.
Осваивая различные водоемы, рыбы приспособились к жизни при разных газовых режимах. Наиболее требовательны к содержанию кислорода в воде лососевые, которым для нормальной жизнедеятельности нужна концентрация кислорода 4,4... 7 мг/л; хариус, голавль, налим хорошо себя чувствуют при содержании не менее 3,1 мг/л; карповым обычно достаточно 1,9... 2,5 мг/л.
Каждому виду свойствен свой кислородный порог, т. е. минимальная концентрация кислорода, при которой рыба гибнет. Форель начинает задыхаться при концентрации кислорода 1,9 мг/л, судак и лещ погибают при 1,2, плотва и красноперка — при 0,25 ... 0,3 мг/л; у сеголетков-карпов, выращенных на естественной пище, кислородный порог отмечен при 0,07 ... 0,25 мг/л, а для двухлетков—0,01 ...0,03 мг/л кислорода. Караси и ротаны — частичные анаэробы — несколько суток могут жить совсем без кислорода, но при низкой температуре. Предполагают, что сначала организм использует кислород из плавательного пузыря, затем — гликоген печени и мышц. По-видимому, рыбы имеют специальные рецепторы в передней части спинной аорты или в продолговатом мозгу, воспринимающие падение концентрации кислорода в кровяной плазме. Выносливости рыб способствует большое количество каротиноидов в нервных клетках мозга, которые способны накапливать кислород и отдавать его при недостатке.
Интенсивность дыхания зависит от биотических и абиотических факторов. Внутри одного вида она изменяется в зависимости от размера, возраста, подвижности, активности питания, пола, степени зрелости гонад, физико-химических факторов среды. По мере роста рыб активность окислительных процессов в тканях уменьшается; созревание гонад, наоборот, вызывает увеличение потребления кислорода. Расход кислорода в организме самцов выше, чем у самок.
На ритм дыхания кроме концентрации в воде кислорода влияют содержание СО2, рН, температура и др. Например, при температуре 10 °С и содержании кислорода 4,7 мг/л форель совершает 60... 70 дыхательных движений в минуту, а при 1,2 кг/л частота дыхания возрастает до 140... 160; карп при 10 °С дышит почти вдвое медленнее, чем форель (30... 40 раз в минуту), зимой он совершает в минуту 3... 4 и даже 1 ... 2 дыхательных движения.
Как и резкий недостаток кислорода, на рыб губительно действует чрезмерное перенасыщение им воды. Так, летальной границей для эмбрионов щуки является 400% насыщения воды кислородом, при 350 .. .430% насыщения нарушается двигательная активность эмбрионов плотвы. Прирост осетровых снижается при 430% насыщения.
Инкубация икры в перенасыщенной кислородом воде приводит к замедлению развития эмбрионов, сильному увеличению отхода и количества уродов и даже гибели. У рыб появляются пузырьки газа на жабрах, под кожей, в кровеносных сосудах, органах, а затем наступают судороги и смерть. Это называется газовая эмболия или газопузырьковая болезнь. Однако гибель наступает не из-за избытка кислорода, а из-за большого количества азота. Например, у лососевых личинки и мальки гибнут при 103 ... 104%, сеголетки — 105 ... 113, взрослые рыбы — при 118% насыщения воды азотом.
Для поддержания оптимальной концентрации кислорода в воде, обеспечивающей наиболее эффективное течение физиологических процессов в организме рыб, нужно использовать аэрационные установки.
К небольшому пересыщению кислорода рыбы адаптируются быстро. У них повышается обмен и как результат увеличивается потребление корма и снижается кормовой коэффициент, развитие эмбрионов ускоряется, отходы снижаются.
Для нормального дыхания рыб очень важно содержание в воде СО2. При большом количестве двуокиси углерода дыхание рыб затруднено, так как уменьшается способность гемоглобина крови связывать кислород, насыщение кислородом крови резко снижается и рыба задыхается. При содержании CO2 в атмосфере 1...5% СО2; крови не может поступать наружу, а кровь не может принимать кислород даже из насыщенной кислородом воды.
Кровеносная система
Главным отличием кровеносной системы рыб от других позвоночных является наличие одного круга кровообращения и двухкамерного сердца, наполненного венозной кровью (за исключением двоякодышащих и кистеперых).
Сердце состоит из одного желудочка и одного предсердия и помещается в околосердечной сумке, сразу за головой, позади последних жаберных дуг, т. е. по сравнению с другими позвоночными сдвинуто вперед. Перед предсердием имеется венозная пазуха, или венозный синус, со спадающими стенками; через эту пазуху кровь поступает в предсердие, а из него — в желудочек.
Расширенный начальный участок брюшной аорты у низших рыб (акулы, скаты, осетровые” двоякодышащи╦) образует сокращающийся артериальный конус, а у высших рыб — луковицу аорты, стенки которой сокращаться не могут. Обратному току крови препятствуют клапаны.
Схема кровообращения в самом общем виде представлена следующим образом. Венозная кровь, заполняющая сердце, при сокращениях сильного мускульного желудочка через артериальную луковицу по брюшной аорте направляется вперед и поднимается в жабры по приносящим жаберным артериям. У костистых рыб их четыре с каждой стороны головы—по числу жаберных дуг. В жаберных лепестках кровь проходит через капилляры и окисленная, обогащенная кислородом направляется по выносящим сосудам (их также четыре пары) в корни спинной аорты, которые затем сливаются в спинную аорту, идущую вдоль тела назад, под позвоночником. Соединение корней аорты спереди образует характерный для костистых рыб - головной круг. Вперед от корней аорты ответвляются сонные артерии.
От спинной аорты идут артерии к внутренним органам и мускулатуре. В хвостовом отделе аорта переходит в хвостовую артерию. Во всех органах и тканях артерии распадаются на капилляры. Собирающие венозную кровь венозные капилляры впадают в вену, несущую кровь к сердцу. Хвостовая вена, начинающаяся в хвостовом отделе, войдя в полость тела, разделяется на воротные вены почек. В почках разветвления воротных вен образуют воротную систему, а выйдя из них, сливаются в парные задние кардинальные вены. В результате слияния вен задних кардинальных с передними кардинальными (яремными), собирающими кровь из головы, и подключичными, приносящими кровь из грудных плавников, образуются два Кювьерова протока, по которым кровь попадает в венозный синус. Кровь из пищеварительного тракта (желудка, кишечника) и селезенки, идущая по нескольким венам, собирается в воротную вену печени, разветвления которой в печени образуют воротную систему. Собирающая кровь из печени печеночная вена впадает прямо в венозный синус
Рис. 1 Схема кровеносной системы костистой рыбы:
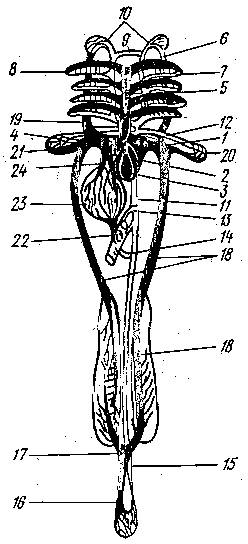 1 - венозная пазуха; 2 - предсердие; 3 - желудочек; 4 - луковица аорты; 5 - брюшная аорта; 6 - приносящие жаберные артерии; выносящие жаберные артерии; 8 - корни спинной аорты; 9 - передняя перемычка, соединяющая корни аорты; 10 - сонная артерия; 11 - спинная аорта; 12 - подключичная артерия; 13 - кишечная артерия; 14 - брыжеечная артерия; 15 - хвостовая артерия; 16 - хвостовая вена; 17 - воротные вены почек; 18 - задняя кардинальная вена; 19 - передняя кардинальная вена; 20 - подключичная вена; 21 - Кювьеров проток; 22 - воротная вена печени; 23 - печень; 24 - печеночная вена; черным показаны сосуды с венозной кровью, белым с артериальной.
1 - венозная пазуха; 2 - предсердие; 3 - желудочек; 4 - луковица аорты; 5 - брюшная аорта; 6 - приносящие жаберные артерии; выносящие жаберные артерии; 8 - корни спинной аорты; 9 - передняя перемычка, соединяющая корни аорты; 10 - сонная артерия; 11 - спинная аорта; 12 - подключичная артерия; 13 - кишечная артерия; 14 - брыжеечная артерия; 15 - хвостовая артерия; 16 - хвостовая вена; 17 - воротные вены почек; 18 - задняя кардинальная вена; 19 - передняя кардинальная вена; 20 - подключичная вена; 21 - Кювьеров проток; 22 - воротная вена печени; 23 - печень; 24 - печеночная вена; черным показаны сосуды с венозной кровью, белым с артериальной.
Как и у других позвоночных, у круглоротых и рыб имеются так называемые дополнительные сердца, поддерживающие давление в сосудах. Так, в спинной аорте радужной форели есть эластичная связка, выполняющая роль нагнетающего насоса, который автоматически увеличивает циркуляцию крови во время плавания, особенно в мускулатуре тела. Интенсивность работы дополнительного сердца зависит от частоты движений хвостового плавника.
У двоякодышащих рыб появляется неполная перегородка предсердия. Это сопровождается и возникновением легочного круга кровообращения, проходящего через плавательный пузырь, превращенный в легкое.
Сердце рыб гораздо меньше и слабее, чем сердце наземных позвоночных. Масса его обычно не превышает 2,5%, в среднем 1% массы тела, тогда как у млекопитающих оно достигает 4,6%, а у птиц даже 16%.
Кровяное давление (Па) у рыб низкое—2133,1 (скат), 11198,8 (щука), 15998,4 (лосось), тогда как в сонной артерии лошади — 20664,6.
Невелика и частота сокращений сердца—18...30 ударов в минуту, причем она сильно зависит от температуры: при низкой температуре у рыб, зимующих на ямах, она уменьшается до 1...2; у рыб, переносящих вмерзание в лед, пульсация сердца на этот период прекращается.
Количество крови у рыб меньше, чем у всех остальных позвоночных животных (1,1,..7,3% массы тела, в том числе у карпа 2,0...4,7%, сома—до 5, щуки—2, кеты--1,6, тогда как у млекопитающих—6,8% в среднем). Это связано с горизонтальным положением тела (нет необходимости проталкивать кровь вверх) и меньшими энергетическими тратами в связи с жизнью в водной среде. Вода является гипогравитационной средой, т. е. сила земного притяжения здесь почти не сказывается.
Морфологическая и биохимическая характеристика крови различна у разных видов в связи с систематическим положением, особенностями среды обитания и образа жизни. Внутри одного вида эти показатели колеблются в зависимости от сезона года, условий содержания, возраста, пола, состояния особей. Эритроциты рыб крупнее, а их количество в крови меньше, чем у высших позвоночных, лейкоцитов же, как правило, больше. Это связано, с одной стороны, с пониженным обменом рыб, а с другой—с необходимостью усилить защитные функции крови, так как окружающая среда изобилует болезнетворными организмами. В 1 мм3 крови количество эритроцитов составляет (млн): у приматов—9,27; копытных—11,36; китообразных— 5,43; птиц—1,61...3,02; костистых рыб—1,71 (пресноводные), 2,26 (морские), 1,49 (проходные).
Количество эритроцитов у рыб колеблется в широких пределах, прежде всего в зависимости от их подвижности: у карпа— 0,84...1,89 млн/мм3 крови, щуки—2,08, пеламиды—4,12млн/мм3. Количество лейкоцитов составляет у карпа 20...80, у ерша — 178 тыс/мм3. Лейкоциты рыб отличаются большим разнообразием. У большинства видов в крови имеются и зернистые (нейтрофилы, эозинофилы), и незернистые (лимфоциты, моноциты) формы лейкоцитов. Преобладают лимфоциты, на долю которых приходится 80...95%, моноциты составляют 0,5...11%, нейтрофилы—13...31%. Эозинофилы встречаются редко. Например, они есть у карповых, амурских растительноядных и некоторых окуневых рыб.
Соотношение разных форм лейкоцитов в крови карпа зависит от возраста и условий выращивания.
Количество лейкоцитов сильно изменяется в течение года:
у карпа оно повышается летом и понижается зимой при голодании в связи со снижением интенсивности обмена.
Разнообразие форм, размеров и количества характерно и для тромбоцитов, участвующих в свертывании крови.
Кровь рыб окрашена гемоглобином в красный цвет, но есть рыбы и с бесцветной кровью. У таких рыб кислород в растворенном состоянии переносится плазмой. Так, у представителей семейства Chaenichthyidae (из подотряда нототениевых), обитающих в антарктических морях в условиях низкой температуры (<2°С), в воде, богатой кислородом, эритроцитов и гемоглобина в крови нет. Дышат они через кожу, в которой очень много капилляров: протяженность капилляров на 1 мм2 поверхности тела достигает 45 мм. Кроме того, у них ускорена циркуляция крови в жабрах. У нототениевых, тресковых и других обитателей полярных широт в крови образуются вещества (антифризы), благодаря которым они не замерзают при отрицательной температуре.
Количество гемоглобина в организме рыб значительно меньше, чем у наземных позвоночных: на 1 кг тела у них приходится 0,5...4 г, тогда как у млекопитающих он составляет 5...25 г. У рыб, передвигающихся быстро, гемоглобина больше, чем у малоподвижных: у проходного осетра 4 г/кг, у налима 0,5 г/кг. Количество гемоглобина зависит от сезона (у карпа повышается зимой и понижается летом), гидрохимического режима водоема (в воде с рН 5,2 количество гемоглобина в крови возрастает), условий питания (карпы, выращенные на естественной пище и дополнительных кормах, имеют разное количество гемоглобина). Темп роста рыб зависит от количества гемоглобина.
Жизнь в среде с небольшим содержанием кислорода определила низкую интенсивность обмена и более высокую способность насыщения при более низком парциальном давлении кислорода в отличие от позвоночных, дышащих воздухом. Способность гемоглобина извлекать кислород из воды у разных рыб неодинакова. У быстро плавающих (макрели, трески, форели) гемоглобина в крови много, и они очень требовательны к содержанию кислорода в воде. У многих морских придонных рыб, а также угря, карпа, карасей и некоторых других, наоборот, гемоглобина в крови мало, но он может забирать кислород из среды даже с незначительным количеством.
Например, судаку для насыщения крови кислородом (при 16 °С) необходимо содержание в воде 2,1...2,3 О2 мг/л; при наличии в воде 0,56...0,6 O2 мг/л кровь начинает его отдавать, дыхание оказывается невозможным, и рыба гибнет. Лещу при этой же температуре для полного насыщения гемоглобина кислородом достаточно присутствие в литре, воды 1,0...1,06 мг кислорода.
Чувствительность рыб к изменениям температуры воды также связана со свойствами гемоглобина: при повышении температуры потребность организма в кислороде увеличивается, но способность гемоглобина его забирать—уменьшается.
Уменьшает способность гемоглобина забирать кислород и углекислота: для того чтобы насыщенность крови угря кислородом достигла 50% при содержании в воде 1% СО2, необходимо давление кислорода в 666,6 Па, а в отсутствие CO2 для этого достаточно давления кислорода почти вдвое меньшего — 266,6.„399,9 Па,
Группы крови у рыб впервые были определены на байкальском омуле и хариусе в 30-х годах нынешнего столетия. К настоящему времени установлено, что групповая антигенная дифференцировка эритроцитов широко распространена: выявлено 14 систем групп крови, включающих более 40 эритроцитарных антигенов. При помощи иммуносерологических методов изучают изменчивость на разных уровнях: выявлены различия между видами и подвидами и даже между внутривидовыми группировками у лососевых (при изучении родства форелей), осетровых (при сравнении локальных стад) и других рыб.
Кровь, будучи внутренней средой организма, выполняет важнейшие функции: переносит белки, углеводы (гликоген, глюкоза и др.) и другие питательные вещества, играющие большую роль в энергетическом и пластическом обмене; дыхательную—транспортировка кислорода к тканям и углекислоты к органам дыхания; выделительную—вынос конечных продуктов обмена к органам выделения; регуляторную—перенос гормонов и других активных веществ от желез внутренней секреции к органам и тканям; защитную—в крови содержатся противомикробные вещества (лизоцим, комплемент, интерферон, пропердин), образуются антитела, циркулирующие в ней лейкоциты обладают фагоцитарной способностью. Уровень этих веществ в крови зависит от биологических особенностей рыб и абиотических факторов, а подвижность состава крови позволяет использовать ее показатели для оценки физиологического состояния.
Костного мозга, являющегося основным органом образования форменных элементов крови у высших позвоночных, и лимфатических желез (узлов) у рыб нет.
Кроветворение у рыб по сравнению с высшими позвоночными отличается рядом особенностей.
1. Образование клеток крови происходит во многих органах. Очагами кроветворения являются: жаберный аппарат (эндотелий сосудов и ретикулярный синцитий, сосредоточенный у основания жаберных лепестков), кишечник (слизистая), сердце (эпителиальный слой и эндотелий сосудов), почки (ретикулярный синцитий между канальцами), селезенка, сосудистая кровь, лимфоидный орган (скопления кроветворной ткани — ретикулярного синцития—под крышей черепа). На отпечатках этих органов видны кровяные клетки разных стадий развития.
2. У костистых рыб наиболее активно гемопоэз происходит в лимфоидных органах, почке и селезенке, причем главным органом кроветворения являются почки, а именно их передняя часть. В почках и селезенке происходит как образование эритроцитов, лейкоцитов, тромбоцитов, так и распад эритроцитов.
3. Наличие в периферической крови рыб и зрелых и молодых эритроцитов является нормальным и не служит патологическим показателем в отличие от крови взрослых млекопитающих.
4. В эритроцитах имеется ядро, как и у других водных животных, вследствие чего жизнеспособность их дольше, чем млекопитающих.
Селезенка рыб располагается в передней части полости тела, между петлями кишечника, но независимо от него. Это плотное компактное темно-красное образование различной формы (шарообразной, лентовидной), но чаще вытянутой.
Селезенка быстро меняет объем под влиянием внешних условий и состояния рыбы. У карпа она увеличивается зимой, когда в связи с пониженным обменом веществ ток крови замедляется и она скапливается в селезенке, печени и почках, которые служат депо крови, то же наблюдается при острых заболеваниях. При недостатке кислорода, загрязнении воды, перевозке и сортировке рыбы, облове прудов запасы из селезенки поступают в кровеносное русло.
Одним из важнейших факторов внутренней среды является осмотическое давление крови, так как от него зависит взаимодействие крови и клеток тела, водный обмен в организме.
Кровеносная система подчиняется нервной (блуждающий нерв) и гуморальной (гормоны, ионы Са, К) регуляции. Центральная нервная система рыб получает информацию о работе сердца от барорецепторов жаберных сосудов.
Лимфатическая система рыб не имеет желез. Она представлена рядом парных и непарных лимфатических стволов, в которые лимфа собирается из органов и по ним же выводится в конечные участки вен, в частности в Кювьеровы протоки, У некоторых рыб есть лимфатические сердца.
НЕРВНАЯ СИСТЕМА И ОРГАНЫ ЧУВСТВ
Нервная система. У рыб она представлена центральной нервной системой и связанной с ней периферической и вегетативной (симпатической) нервной системой.
Центральная нервная система состоит из головного и спинного мозга. К периферической нервной системе относятся нервы, отходящие от головного и спинного мозга к органам. Вегетативная нервная система в основе имеет многочисленные ганглии и нервы, иннервирующие мышцы внутренних органов и кровеносных сосудов сердца.
Нервная система рыб по сравнению с нервной системой высших позвоночных характеризуется рядом примитивных черт.
Центральная нервная система имеет вид нервной трубки, тянущейся вдоль туловища: часть ее, лежащая над позвоночником и защищенная верхними дугами позвонков, образует спинной мозг, а расширенная передняя часть, окруженная хрящевым или костным черепом, составляет головной мозг.
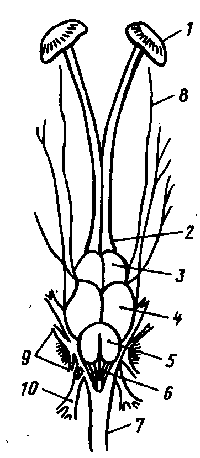
Рис. 1 Головной мозг рыбы (окунь):
1- обонятельные капсулы; 2- обонятельные доли; 3- передний мозг; 4- средний мозг; 5- мозжечок; 6- продолговатый мозг; 7- спинной мозг; 8,9,10- головные нервы.
Полости переднего, промежуточного и продолговатого мозга называются желудочками: полость среднего мозга — сильвиевым водопроводом (она соединяет полости промежуточного и продолговатого мозга, т. е. третий и четвертый желудочки).
Передний мозг благодаря продольной борозде имеет вид двух полушарий. К ним прилегают обонятельные луковицы (первичный обонятельный центр) или непосредственно "^у большинства видов), или через обонятельный тракт (карповые, сомовые, тресковые).
В крыше переднего мозга нет нервных клеток. Серое вещество в виде полосатых тел сосредоточено главным образом в основании и обонятельных долях, выстилает полость желудочков и составляет главную массу переднего мозга. Волокна обонятельного нерва связывают луковицу с. клетками обонятельной капсулы.
Передний мозг является центром обработки информации, поступающей от органов обоняния. Благодаря своей связи с промежуточным и средним мозгом он участвует в регуляции движения и поведения. В частности, передний мозг принимает участие в формировании способности к таким актам, как икрометание, охрана икры, образование стаи, агрессия и др.
В промежуточном мозге развиты зрительные бугры. От них отходят зрительные нервы, образующие хиазму (перекрест, т. е. часть волокон правого нерва переходит в левый нерв и наоборот). На нижней стороне промежуточного мозга, или гипоталамусе, имеется воронка, к которой прилегает гипофиз, или питуитарная железа; в верхней части промежуточного мозга развивается эпифиз, или пинеальная железа. Гипофиз и эпифиз являются железами внутренней секреции.
Промежуточный мозг выполняет многочисленные функции. Он воспринимает раздражения от сетчатки глаза, участвует /в координации движений, переработке информации от других органов чувств. Гипофиз и эпифиз осуществляют гормональную регуляцию обменных процессов.
Средний мозг наибольший по объему. Он имеет вид двух полушарий, которые называют зрительными долями. Эти доли являются первичными зрительными центрами, воспринимающими возбуждение. Из них берут начало волокна зрительного нерва.
В среднем мозгу обрабатываются сигналы, идущие от органов зрения и равновесия; здесь помещаются центры связи с мозжечком, продолговатым и спинным мозгом, регуляции окраски, вкуса.
Мозжечок расположен в задней части мозга и может иметь форму или маленького бугорка, прилегающего сзади к среднему мозгу, или большого мешковидно-вытянутого образования, примыкающего сверху к продолговатому мозгу. Особенно большого развития достигает мозжечок у сомов, а у мормируса он наибольший среди всех позвоночных. В мозжечке рыб имеются клетки Пуркинье.
Мозжечок является центром всех моторных иннерваций при плавании и схватывании пищи. Он 'обеспечивает координацию движений, поддержание равновесия, мышечную деятельность, связан с рецепторами органов боковой линии, направляет и координирует деятельность других отделов мозга. При повреждении мозжечка, например, у карпа и серебряного карася наступает атония мышц, нарушается- равновесие, не вырабатываются или пропадают условные рефлексы на свет и звук.
Пятый отдел головного мозга—продолговатый мозг без резкой границы переходит в спинной мозг. Полость продолговатого мозга — четвертый желудочек продолжается в полость
спинного мозга--невроцель. Значительная масса продолгоча-того мозга состоит из белого вещества.
От продолговатого мозга отходит большая часть (шесть из десяти) черепно-мозговых нервов. Он является центром регуляции деятельности спинного мозга и вегетативной нервной системы. В нем располагаются наиболее важные жизненные центры, регулирующие деятельность дыхательной, скелетно-мышечной, кровеносной, пищеварительной, выделительной систем, органов слуха и равновесия, вкуса, боковой линии и электрических органов. Поэтому при разрушении продолговатого мозга, например при перерезке туловища позади головы, наступает быстрая смерть рыбы.
Через приходящие в продолговатый мозг спинномозговые волокна осуществляется связь продолговатого и спинного мозга.
От головного мозга отходят 10 пар черепно-мозговых нервов: 1—обонятельный нерв (nervus olfactorius) от чувствующего эпителия обонятельной капсулы доводит раздражения до обонятельных луковиц переднего мозга; 2—зрительный нерв (n. opticus) тянется до сетчатки глаза от зрительных бугров промежуточного мозга; 3—глазодвигательный нерв (n. oculo-motorius) иннервирует мышцы глаза, отходя от среднего мозга;
4 — блоковый нерв (n. trochlearis) — глазодвигательный, тянущийся от среднего мозга к одной из мышц глаза; 5—тройничный нерв (n. trigeminus), отходящий от боковой поверхности продолговатого мозга и дающий три основные ветви—глазничную, верхнечелюстную и нижнечелюстную; 6 — отводящий нерв (n. abducens) тянется от дна мозга к прямой мышце глаза; 7—лицевой нерв (n. facialis) отходит от продолговатого мозга и дает многочисленные разветвления к мускулатуре подъязычной дуги, слизистой ротовой полости, коже головы (в том числе боковой линии головы); 8—слуховой нерв (n. acusticus) связывает продолговатый мозг и слуховой аппарат; 9—языко-глоточный нерв (n. glossopharingeus) идет от продолговатого мозга к глотке, иннервирует слизистую глотки и мускулатуру первой жаберной дуги; 10—блуждающий нерв (n. vagus) — наиболее длинный, связывает продолговатый мозг с жаберным аппаратом, кишечным трактом, сердцем, плавательным пузырем, боковой линией.
Степень развития разных отделов головного мозга различна у разных групп рыб и связана с образом жизни.
Передний мозг и обонятельные доли лучше развиты у хрящевых рыб (акулы и скаты) и хуже у костистых. У малоподвижных, например донных рыб (камбалы), мозжечок мал, но сильнее развиты передний и продолговатый отделы мозга в соответствии с большой ролью обоняния и осязания в их жизни. У хорошо плавающих рыб (пелагических, питающихся планктоном, и хищничающих) большее развитие получают средний мозг (зрительные доли) и мозжечок (в связи с необходимостью быстрой координации движения). Рыбы, обитающие в мутной воде, имеют маленькие зрительные доли и небольшой мозжечок. Слабо развиты зрительные доли у глубоководных рыб. Различна и электрическая активность разных отделов мозга: у серебряного карася электрические волны в мозжечке идут с частотой 25...35 раз в секунду, в переднем мозгу— 4...8.
Спинной мозг является продолжением продолговатого мозга. Он имеет форму округлого тяжа и лежит в канале, образованном верхними дугами позвонков. В отличие от высших позвоночных он способен к регенерации и восстановлению деятельности. В спинном мозге серое вещество расположено внутри, а белое—снаружи.
Функция спинного мозга — рефлекторная и проводящая. В нем находятся центры сосудодвигательные, туловищной мускулатуры, хроматофоров, электрических органов. От спинного мозга метамерно, т. е. соответственно каждому позвонку, отходят спинномозговые нервы, иннервирующие поверхность тела, туловищные мышцы, а благодаря соединению спинномозговых нервов с ганглиями симпатической нервной системы—и внутренние органы. В спинном мозгу костистых рыб имеется секреторный орган — урогипофиз, клетки которого вырабатывают гормон, участвующий в водном обмене.
Вегетативная нервная система у хрящевых рыб представлена разобщенными ганглиями, лежащими вдоль позвоночника. Клетки ганглиев своими отростками контактируют со спинномозговыми нервами и внутренними органами.
У костистых рыб ганглии вегетативной нервной системы соединяются двумя продольными нервными стволами. Соединительные ветви ганглиев связывают вегетативную нервную систему с центральной. Взаимосвязи центральной и вегетативной нервной систем создают возможность некоторой взаимозаменяемости нервных центров.
Вегетативная нервная система действует независимо от центральной нервной системы и определяет непроизвольную автоматическую деятельность внутренних органов даже в том случае, если ее связь с центральной нервной системой нарушена.
Реакцию организма рыбы на внешние и внутренние раздражения определяет рефлекс. У рыб можно выработать, условный рефлекс на свет, форму, запах, вкус, звук, температуру воды и соленость. Так, аквариумные и прудовые рыбы вскоре после начала регулярного кормления скапливаются в определенное время у кормушек. Привыкают они и к звукам во время кормления (постукивание по стенкам аквариума, звон колокольчика, свист, удары) и какое-то время подплывают на эти раздражители и при отсутствии пищи. При этом рефлексы на получение корма образуются у рыб быстрее, а пропадают медленнее, чем у кур, кроликов, собак, обезьян. У карасей рефлекс появляется через 8 сочетаний условного раздражителя с безусловным, а затухает через 28...78 неподкрепленных сигналов.
Поведенческие реакции вырабатываются у рыб быстрее в Группе (подражание, движение за лидером в стае, реакция на хищника и др.). Временная память и дрессировка имеет большое значение и в рыбоводной практике. Если рыб не учат оборонительным реакциям, навыкам общения с хищниками, то молодь, выпускаемая из рыбоводных заводов, быстро гибнет в естественных условиях.
Органы восприятия окружающей среды (органы чувств) рыб обладают рядом особенностей, отражающих их приспособленность к условиям жизни. Способность рыб воспринимать информацию из окружающей среды многообразна. Их рецепторы могут улавливать различные раздражения как физической, так и химической природы: давление, звук, цвет, температуру, электрические и магнитные поля, запах, вкус. Одни раздражения .воспринимаются в результате непосредственного прикосновения (осязание, вкус), другие—на расстоянии.
Органы, воспринимающие химические, тактильные (прикосновение), электромагнитные, температурные и другие раздражения, имеют простое строение. Раздражения улавливаются свободными нервными окончаниями чувствующих нервов на поверхности кожи. У некоторых групп рыб они представлены специальными органами или входят в состав боковой линии.
В связи с особенностями жизненной среды у рыб большое значение имеют системы химического чувства. Химические раздражения воспринимаются при помощи обоняния (ощущения запаха) или органов необонятельной рецепции, обеспечивающих восприятие вкуса, изменение активности среды и т. д.
Химическое чувство называется хеморецепцией, а чувствующие органы — хеморецепторами. Хеморецепция помогает рыбам отыскивать и оценивать пищу, особей своего вида и другого пола, избегать врагов, ориентироваться в потоке, защищать территорию.
Органы обоняния. У рыб, как и других позвоночных, они находятся в передней части головы и представлены парными обонятельными (носовыми) мешками (капсулами), открывающимися наружу отверстиями—ноздрями. Дно носовой капсулы выстлано складками эпителия, состоящего из опорных и чувствующих клеток (рецепторов). Наружная поверхность чувствующей клетки снабжена ресничками, а основание связано с окончаниями обонятельного нерва. Рецепторная поверхность
органа велика: на I кв. мм. обонятельного эпителия приходится у Phoxinus 95000 рецепторных клеток. В обонятельном эпителии многочисленны клетки, секретирующие слизь.
Ноздри расположены у хрящевых рыб на нижней стороне рыла впереди рта, у костистых — на дорсальной стороне между ртом и глазами. Круглоротые имеют по одной ноздре, настоящие рыбы—по две. Каждая ноздря разделяется кожистой перегородкой на две части, называемые отверстиями. Вода проникает в переднее, омывает полость и выходит через заднее отверстие, омывая и раздражая при этом волоски рецепторов.
Под влиянием пахучих веществ в обонятельном эпителии происходят сложные процессы: перемещения липидов, белково-мукополисахаридных комплексов и кислой фосфатазы. Электрическая активность обонятельного эпителия в ответ на разные пахучие вещества различна.
Величина ноздрей связана с образом жизни рыб: у подвижных рыб они небольшие, так как при быстром плавании вода в обонятельной полости обновляется быстро; у рыб малоподвижных ноздри большие, они пропускают через носовую полость больший объем воды, что особенно важно для плохих пловцов, в частности, обитающих у дна.
Рыбы обладают тонким обонянием, т. е. пороги обонятельной чувствительности у них очень низки. Это Особенно относится к ночным и сумеречным рыбам, а также к живущим в мутных водах, которым зрение мало помогает в отыскивании пищи и общении с сородичами.
Наиболее чувствительно обоняние у проходных рыб. Дальневосточные лососи совершенно точно находят путь от мест нагула в море к нерестилищам в верховьях рек, где они вывелись несколько лет назад. При этом они преодолевают огромные расстояния и препятствия—течения, пороги, перекаты. Однако рыбы верно находят путь лишь в том случае, если ноздри открыты, а если они заполнены ватой или вазелином, то рыбы идут беспорядочно. Предполагают, что лососи в начале миграции ориентируются по солнцу и звездам и примерно за 800 км от родной реки безошибочно определяют путь благодаря хеморецепции.
В опытах при смывании носовой полости этих рыб водой с родного нерестилища в обонятельной луковице мозга возникала сильная электрическая реакция. На воду из нижерасположенных притоков реакция была слабой, а на воду с чужих нерестилищ рецепторы вообще не реагировали.
Молодь нерки может различать при помощи клеток обонятельной луковицы воду разных озер, растворы различных аминокислот в разведении Ю"4, а также концентрацию кальция в воде. Не менее поразительна аналогичная способность европейского
угря, мигрирующего из Европы к нерестилищам, расположенным в Саргассовом море. Подсчитано, что угорь в состоянии распознавать концентрацию, создаваемую разведением 1 г фенилэтилового спирта в соотношении 1:3-10-18. Рыбы улавливают феромон страха при концентрации 10-10 г/л: Высокая избирательная чувствительность к гистамину, а также к углекислоте (0,00132...0,0264 г/л) обнаружена у карпа.
Обонятельный рецептор рыб кроме химических способен воспринимать и механические воздействия (струи потока) и изменения температуры.
Органы вкуса. Они представлены вкусовыми почками, образованными скоплениями чувствующих и опорных клеток. Основания чувствующих клеток оплетены концевыми разветвлениями лицевого, блуждающего и языкоглоточного нервов. Восприятие химических раздражителей осуществляется также свободными нервными окончаниями тройничного, блуждающего и спинномозговых нервов.
Восприятие вкуса рыбами не обязательно связано с ротовой пол остью, так как вкусовые почки расположены в слизистой оболочке ротовой полости, на губах, в глотке, на усиках, жаберных лепестках, плавниковых лучах и по всей поверхности тела, в том числе на хвосте.
Сом воспринимает вкус главным образом при помощи усов, так как в их эпидермисе сосредоточены вкусовые почки. Количество этих почек увеличивается по мере увеличения размеров тела рыбы.
Рыбы различают и вкус пищи: горькое, соленое, кислое, сладкое. В частности, восприятие солености связано с ямковид-ным органом, помещающимся в ротовой полости.
Чувствительность органов вкуса у некоторых рыб очень высока: например, пещерные рыбы Anoptichtys, будучи слепыми, ощущают раствор глюкозы в концентрации 0,005%. Рыбы распознают изменения солености до О.З^/оо, рН—0,05...0,007, углекислоты—0,5 г/л, NaCI—0,001...0,005 моля (карповые), а гольян — даже 0,00004 моля.
Органы чувств боковой линии. Специфическим органом, свойственным только рыбам и живущим в воде амфибиям, является орган бокового чувства, или боковой линии. Это сейсмосенсор-ный специализированный кожный орган. Наиболее просто эти органы устроены у круглоротых и личинок карповых. Чувствующие клетки (механорецепторы) лежат среди скоплений эктодермальных клеток на поверхности кожи или в мелких ямках, У основания они оплетены конечными разветвлениями блуждающего нерва, а на участке, возвышающемся над поверхностью, имеют реснички, воспринимающие колебания воды. У большинства взрослых костистых эти органы представляют собой
погруженные в кожу каналы, тянущиеся по бокам тела вдоль средней линии. Канал открывается наружу через отверстия (поры) в чешуйках, расположенных над ним. Разветвления боковой линии имеются и на голове.
На дне канала группами лежат чувствующие клетки с ресничками. Каждая такая группа рецепторных клеток вместе с контактирующими с ними нервными волокнами образует собственно орган—невромаст. Вода свободно протекает через канал и реснички ощущают ее давление. При этом возникают нервные импульсы разной частоты.
Органы боковой линии связаны с центральной нервной системой блуждающим нервом.
Боковая линия может быть полной, т. е, тянуться по всей длине тела, или неполной и даже отсутствовать, но в последнем случае сильно развиваются головные каналы, как, например, у сельдей.
Боковой линией рыба ощущает изменение давления текущей воды, вибрации (колебания) низкой частоты, инфразвуковые колебания и электромагнитные поля. Например, карп улавливает ток при плотности 60 мкА/см2, карась—16 мкА/см2.
Боковая линия улавливает давление движущегося потока, а изменение давления при погружении на глубину она не воспринимает. Улавливая колебания водной толщи, рыба обнаруживает поверхностные волны, течения, подводные неподвижные (скалы, рифы) и движущиеся (враги, добыча) предметы.
Боковая линия — весьма чувствительный орган: акула улавливает движение рыб на расстоянии 300 м, проходные рыбы ощущают в море даже незначительные токи пресной воды.
Способность улавливать отраженные от живых и неживых объектов волны очень важна для глубоководных рыб, так как в темноте больших глубин невозможно обычное зрительное восприятие.
Предполагают, что во время брачных игр рыбы воспринимают боковой линией волны как сигнал самки или самца к нересту. Функцию кожного чувства выполняют и так называемые кожные почки — клетки, имеющиеся в покровах головы и усиков, к которым подходят нервные окончания, однако они имеют гораздо меньшее значение.
Органы осязания. Ими служат скопления чувствующих клеток (осязательные тельца), разбросанные по поверхности тела. Они воспринимают прикосновение твердых предметов (тактильные ощущения), давление воды, изменение температуры и боль.
Особенно много чувствующих кожных почек находится во рту и на губах. У некоторых рыб функцию этих органов выполняют удлиненные лучи плавников: у гурами это первый луч брюшного плавника, у триглы (морской петух) осязание связано с лучами грудных плавников, ощупывающими дно. У обитателей мутных вод или донных рыб, наиболее активных ночью, наибольшее количество чувствующих почек сосредоточено на усиках и плавниках. У самцов усы служат рецепторами вкуса.
Механические травмы и боль рыбы, по-видимому, ощущают слабее, чем другие позвоночные. Так, акулы, набросившиеся на добычу, не реагируют на удары острым предметом в голову.
Терморецепторы. Ими являются находящиеся в поверхностных слоях кожи свободные окончания чувствующих нервов, при помощи которых рыбы воспринимают температуру воды. Различают рецепторы, воспринимающие тепло (тепловые) и холод (холодовые). Точки восприятия тепла найдены, например, у щуки на голове, восприятия холода — на поверхности тела. Костистые рыбы улавливают перепады температуры в 0,1...0,4 град. У форели можно выработать условный рефлекс на очень малые (менее 0,1 град.) и быстрые изменения температуры.
Очень чувствительны к температуре боковая линия и головной мозг. В мозгу рыб обнаружены чувствительные к температуре нейроны, сходные с нейронами в центрах терморегуляции млекопитающих. У форели в промежуточном мозгу имеются нейроны, реагирующие на повышение и понижение температуры.
Органы электрического чувства. Органы восприятия электрического и магнитного полей располагаются в коже на всей поверхности тела рыб, но главным образом в разных участках головы и вокруг нее. Они сходны с органами боковой линии:
это ямки, заполненные слизистой массой, хорошо проводящей ток; на дне ямок помещаются чувствующие клетки (электрорецепторы), передающие 'нервные импульсы в мозг. Иногда они входят в состав системы боковой линии. Электрическими рецепторами у хрящевых рыб служат и ампулы Лоренцини. Анализ информации, получаемой электрорецепторами, осуществляет анализатор боковой линии, который расположен в продолговатом мозгу и мозжечке. Чувствительность рыб к току велика — до 1 мкВ/см2: карп ощущает ток напряжением 0,06...0,1, форель—0,02...0,08, карась 0,008...0,0015 В. Предполагают, что восприятие изменения электромагнитного поля Земли позволя-
ет рыбам обнаруживать приближение землетрясения за 6...24 ч до начала в радиусе до 2 тыс. км.
Органы зрения. Они устроены в основном так же, как у других позвоночных. Сходен с остальными позвоночными у них и механизм восприятия зрительных ощущений: свет проходит в глаз через прозрачную роговицу, далее зрачок (отверстие в радужной оболочке) пропускает его на хрусталик, а хрусталик передает (фокусирует) свет на внутреннюю стенку глаза (сетчатку), где и происходит его непосредственное восприятие (рис. 3). Сетчатка состоит из светочувствительных (фоторецепторные), нервных и опорных клеток.
Светочувствительные клетки располагаются со стороны пигментной оболочки. В их отростках, имеющих форму палочек и колбочек, имеется светочувствительный пигмент. Количество этих фоторецепторных клеток очень велико: на 1 мм2 сетчатки у карпа их насчитывается 50 тыс. шт., у кальмара—162 тыс. шт., паука—16, человека—400 тыс. шт. Посредством сложной системы контактов конечных разветвлений чувствующих клеток и дендритов нервных клеток световые раздражения поступают в зрительный нерв.
Колбочки при ярком свете воспринимают детали предметов и цвет: они улавливают длинные волны спектра. Палочки воспринимают слабый свет, но детального изображения создать не могут: воспринимая короткие волны, они примерно в 1000 раз чувствительнее колбочек.
Положение и взаимодействие клеток пигментной оболочки, палочек и колбочек меняется в зависимости от освещенности. На свету пигментные клетки расширяются и прикрывают находящиеся около них палочки; колбочки подтягиваются к ядрам клеток и таким образом передвигаются к свету. В темноте к ядрам подтягиваются палочки и оказываются ближе к поверхности; колбочки приближаются к пигментному слою, а сократившиеся в темноте пигментные клетки прикрывают их.
Количество рецепторов разного рода зависит от образа жизни рыб. У дневных рыб в сетчатке превалируют колбочки, у сумеречных и ночных—палочки: у налима палочек в 14 раз больше, чем у щуки. У глубоководных рыб, живущих в темноте глубин, колбочек нет, а палочки становятся больше и количество их резко увеличивается—до 25 млн на 1 мм2 сетчатки; вероятность улавливания даже слабого света возрастает. Большая часть рыб различает цвета. Некоторые особенности в строении глаз рыб связаны с особенностями жизни в воде. Они эллипсовидной формы и имеют серебристую оболочку между сосудистой и белковой, богатую кристалликами гуанина, что придает глазу зеленовато-золотистый блеск. Роговица у
рыб почти плоская (а не выпуклая), хрусталик шаровидный (а не двояковыпуклый)—это расширяет поле зрения. Отверстие .в радужной оболочке (зрачок) может изменять диаметр только в небольших пределах. Век у рыб, как правило, нет. Лишь акулы имеют мигательную перепонку, закрывающую глаз, как занавеска, и некоторые сельди и кефали имеют жировое веко—прозрачную пленку, закрывающую часть глаза.
Расположение глаз у большинства видов по бокам головы является причиной того, что рыбы обладают в основном монокулярным зрением, а способность к бинокулярному зрению ограничена. Шаровидность хрусталика и перемещение его вперед к роговице обеспечивает широту поля зрения: свет в глаз попадает со всех сторон. Угол зрения по вертикали составляет 150°, по горизонтали—168...170°. Но вместе с тем шаровидность хрусталика обусловливает близорукость рыб. Дальность их зрения ограничена и колеблется в связи с мутностью воды от нескольких сантиметров до нескольких десятков метров. Видение на дальние расстояния становится возможным благодаря тому, что хрусталик может быть оттянут специальной мышцей—серповидным отростком, идущим от сосудистой оболочки дна глазного бокала, а не за счет изменения кривизны хрусталика, как у млекопитающих.
При помощи зрения рыбы ориентируются и относительно предметов, находящихся на земле.
Улучшение зрения в темноте достигается наличием отражательного слоя (тапетум) —кристалликов гуанина, подстилаемых пигментом. Этот слой т пропускает свет к лежащим позади сетчатки тканям, а отражает его и возвращает вторично
на сетчатку. Так увеличивается возможность рецепторов использовать свет, попавший в глаз.
В связи с условиями обитания глаза рыб могут сильно видоизменяться. У пещерных или абиссальных (глубоководных) форм глаза могут редуцироваться и даже исчезать. Некоторые же глубоководные рыбы, наоборот, имеют огромные глаза, позволяющие улавливать совсем слабый свет, или телескопические глаза, собирающие линзы которых рыба может поставить параллельно и обрести бинокулярное зрение. Глаза некоторых угрей и личинок тропических рыб вынесены вперед на длинных выростах (стебельчатые глаза). Необычна модификация глаз у четырехглазки, обитающей в водах Центральной и Южной Америки. Ее глаза помещаются на верху головы, каждый из них разделен перегородкой на две самостоятельные части:
верхней рыба видит в воздухе, нижней—в воде. В воздушной среде могут функционировать глаза рыб, выползающих на сушу.
Кроме глаз воспринимают свет эпифиз (железа внутренней секреции) и светочувствительные клетки, расположенные в хвостовой части, например,у миног.
Роль зрения как источника информации для большинства рыб велика: при ориентации во время движения, отыскивании » захвате пищи, сохранении стаи, в нерестовый период (восприятие оборонительных и агрессивных поз и движений самцами—соперниками, а между особями разных полов'—брачного наряда и нерестового «церемониала»), в отношениях жертва—хищник и т. д. Карп видит при освещенности 0,0001 лк, карась—0,01 лк.
Способность рыб воспринимать свет издавна использовалась в рыболовстве: лов рыбы на свет.
Известно, что рыбы разных видов неодинаково реагируют на свет разной интенсивности и разной длины волны, т. е. разного цвета. Так, яркий искусственный свет привлекает одних рыб (каспийская килька, сайра, ставрида, скумбрия) и отпугивает других (кефаль, минога, угорь). Так же избирательно относятся разные виды к разным цветам и разным источникам света—надводным и подводным. Все это положено в основу организации промышленного лова рыбы на электросвет. Так ловят кильку, сайру и других рыб.
Орган слуха и равновесия рыб. Он расположен в задней части черепной коробки и представлен лабиринтом. Ушных отверстий, ушной раковины и улитки нет, т. е. орган слуха представлен внутренним ухом.
Наибольшей сложности достигает он у настоящих рыб:
большой перепончатый лабиринт помещается в хрящевой или костной камере под прикрытием ушных костей. В нем различают верхнюю часть — овальный мешочек (ушко, utriculus) и нижнюю—круглый мешочек (sacculus). От верхней. части во взаимно перпендикулярных направлениях отходят три полукружных канала, каждый из которых на одном конце расширен в ампулу
Овальный мешочек с полукружными каналами составляет орган равновесия (вестибулярный аппарат). Боковое расширение нижней части круглого мешочка (lagena), являющегося зачатком улитки, не получает у рыб дальнейшего развития. От круглого мешочка отходит внутренний лимфатический (эндолимфатический) канал, который у акул и скатов через специальное отверстие в черепе выходит наружу, а у остальных рыб слепо заканчивается у кожи головы.
Эпителий, выстилающий отделы лабиринта, имеет чувствующие клетки с волосками, отходящими во внутреннюю полость. Основания их оплетены разветвлениями слухового нерва.
Полость лабиринта заполнена эндолимфой, в ней находятся «слуховые» камешки, состоящие из углекислой извести (отолиты), по три с каждой стороны головы: в овальном и круглом мешочках и лагене. На отолитах, как и на чешуе, образуются концентрические слои, поэтому отолиты, особенно наибольший,-часто используют для определения возраста рыб, а иногда и для систематических определений, так как их размеры и контуры неодинаковы у различных видов.
У большинства рыб наибольший отолит располагается в круглом мешочке, но у карповых и некоторых других—в лагене.
С лабиринтом связано чувство равновесия: при передвижении рыбы давление эндолимфы в полукружных каналах, а также со стороны отолита изменяется, и возникшее раздражение улавливается нервными окончаниями. При экспериментальном разрушении верхней части лабиринта с полукружными каналами рыба теряет способность удерживать равновесие и лежит на боку, спине или брюхе. Разрушение нижней части лабиринта не ведет к утрате равновесия.
С нижней частью лабиринта связано восприятие звуков: при удалении нижней части лабиринта с круглым мешочком и лаге-ной рыбы не могут различать звуковые тона, например, при выработке условных рефлексов. Рыбы без овального мешочка и полукружных каналов, т. е. без верхней части лабиринта, дрессировке поддаются. Таким образом, установлено, что рецепторами звука являются именно круглый мешочек и лагена.
Рыбы воспринимают как механические, так и звуковые колебания частотой от 5 до 25 Гц органами боковой линии, от 16 до 13000 Гц:—лабиринтом. Некоторые виды рыб улавливают колебания, находящиеся на границе инфразвуковых волн, боковой линией, лабиринтом и кожными рецепторами.
Острота слуха у рыб меньше, чем у высших позвоночных, и у разных .видов неодинакова: язь воспринимает колебания, длина волны которых составляет 25. ..5524 Гц, серебряный карась—25...3840, угорь—36...650 Гц, причем низкие звуки улавливаются ими лучше. Акулы слышат звуки, издаваемые рыбами на расстоянии 500 м.
Рыбы улавливают и те звуки, источник которых находится не в воде, а в атмосфере, несмотря на то, что такой звук на 99,9% отражается поверхностью воды и, следовательно, в воду проникает только 0,1% образующихся звуковых волн.
В восприятии звука у карповых и сомовых рыб большую роль играет плавательный пузырь, соединенный с лабиринтом и служащий резонатором.
Рыбы могут и сами издавать звуки. Звукоиздающие органы у рыб различны. Это плавательный пузырь (горбыли, губаны и др.), лучи грудных плавников в комбинации с костями плечевого пояса (сомы), челюстные и глоточные зубы (окуневые и карповые) и др. В связи с этим неодинаков и характер звуков. Они могут напоминать удары, цоканье, свист, ворчанье, хрюканье, писк, кваканье,, рычание, треск, рокот, звон, хрип, гудок, крики птиц и стрекотание насекомых.
Сила и частота звуков, издаваемых рыбами одного вида, зависит от пола, возраста, пищевой активности, здоровья, причиняемой боли и др.
Звучание и восприятие звуков имеет большое значение в жизнедеятельности рыб. Оно помогает особям разного пола найти друг друга, сохранить стаю, сообщить сородичам о присутствии пищи, охранять территорию, гнездо и потомство от врагов, является стимулятором созревания во время брачных игр, т. е. служит важным средством общения. Предполагают, что у глубоководных рыб, рассредоточенных в темноте на океанических глубинах, именно слух в сочетании с органами боковой линии и обонянием обеспечивает общение, тем более, что звукопроводимость, более высокая в воде, чем в воздухе, на глубине возрастает. Особенно важен слух для ночных рыб и обитателей мутных вод.
Реакция разных рыб на посторонние звуки различна: при шуме одни уходят в сторону, другие (толстолобик, семга, кефаль) выпрыгивают из воды. Это используют при организации лова. В рыбоводных хозяйствах, в период нереста, движение транспорта около нерестовых прудов запрещено.
Железы внутренней секреции
Железами внутренней секреции являются гипофиз, эпифиз, надпочечники, поджелудочная, щитовидная и ультимобронхиальная (подпищеводная) железы, а также урогипофиз и гонады, Они выделяют гормоны в кровь.
Гипофиз—непарное, неправильной овальной формы образование, отходящее от нижней стороны промежуточного мозга (гипоталамуса). Очертание, размеры и положение его чрезвычайно разнообразны. У сазана, карпа и многих других рыб гипофиз сердцевидной формы, лежит почти перпендикулярно мозгу. У серебряного карася он вытянут, немного сплющен с боков и лежит параллельно мозгу.
В гипофизе различают два основных отдела различного происхождения: мозговой (нейрогипофиз), составляющий внутреннюю часть железы, который развивается из нижней стенки промежуточного мозга как впячивание дна третьего мозгового желудочка, и железистый (аденогипофиз), образующийся из впячивания верхней стенки глотки. В аденогипофизе выделяют три части (лопасти, доли): главную (переднюю, расположенную на периферии), переходную (наибольшую) и промежуточную (рис. 34). Аденогипофиз является центральной железой эндокринной системы. В железистой паренхиме его долей вырабатывается секрет, содержащий ряд гормонов, стимулирующих рост (соматический гормон необходим для роста костей), регулирующих функции половых желез и таким образом воздействующих на половое созревание, влияющих на деятельность пигментных клеток (определяют окраску тела и прежде всего появление брачного наряда) и повышающих устойчивость рыб к высокой температуре, стимулирует синтез белка, работу щитовидной железы, участвует в осморегуляции. Удаление гипофиза влечет за собой остановку роста и созревания.
Гормоны, выделяемые нейрогипофизом, синтезируются в ядрах гипоталамуса и переносятся по нервным волокнам в нейрогипофиз, а затем попадают в пронизывающие его капилляры, Таким образом, это нейтросекреторная железа. Гормоны принимают участие в осморегуляции, вызывают нерестовые реакции.
Единую систему с гипофизом образует гипоталамус, клетки которого выделяют секрет, регулирующий гормонообразующую деятельность гипофиза, а также водно-солевой обмен и др.
Наиболее интенсивное развитие гипофиза приходится на период превращения личинки в малька, У половозрелых рыб активность его неравномерна в связи с биологией размножения рыб и, в частности, с характером икрометания. У единовременно икромечущих рыб секрет в железистых клетках накапливается почти одновременно 'после выведения секрета, к моменту овуляции гипофиз опустошается, и в секреторной деятельности его наступает перерыв, В яичниках к моменту нереста заканчивается развитие овоцитов, подготовляемых к вымету в данный сезон. Овоциты выметываются в один прием и составляют таким образом единственную генерацию,
У порционно икромечущих рыб секрет в клетках образуется неодновременно. Вследствие этого после вывода секрета во время первого нереста остается часть клеток, в которых процесс образования коллоида не закончился. В результате он может выделяться порциями на протяжении всего нерестового периода. В свою очередь, овоциты, подготавливаемые к вымету в данный сезон, развиваются также асинхронно. К моменту первого нереста в яичниках содержатся не только созревшие овоциты, но и те, развитие которых еще не завершено. Такие овоциты созревают через некоторое время после выведения первой генерации овоцитов, т. е. первой порции икры. Так образуется несколько порций икры.
Исследование путей стимуляции созревания рыб привели почти одновременно в первой половине нашего века, но независимо друг от друга бразильских (Иеринг и Кардозо, 1934— 1935) и советских ученых (Гербильский и его школа, 1932— 1934) к разработке метода гипофизарных инъекций производителям для ускорения их созревания. Этот метод позволил в значительной мере управлять процессом созревания рыб и тем самым увеличивать размах рыбоводных работ по воспроизводству ценных видов. Гипофизарные инъекции широко применяют при искусственном разведении осетровых и карповых рыб.
Третий нейросекреторный отдел промежуточного мозга•— эпифиз. Его гормоны (серотин, мелатонин, адреногломеруло-тропин) участвуют в сезонных перестройках обмена веществ. На его активность влияют освещенность и продолжительность светового дня: при их увеличении повышается активность рыб, ускоряется рост, изменяются гонады и др.
Щитовидная железа расположена в области глотки, около брюшной аорты. У одних рыб (некоторые акулы, лососевые) она является плотным парным образованием, состоящим из фолликулов, выделяющих гормоны, у других (окуневые, карповые) железистые клетки не образуют оформленного органа, а лежат диффузно в соединительной ткани.
Секреторная деятельность щитовидной железы начинается очень рано. Например, у личинок осетра на 2-й день после выклева железа, хотя и не вполне сформированная, проявляет активную секреторную деятельность, а на 15-й день формирование фолликулов почти заканчивается. Содержащие коллоид фолликулы обнаруживаются у 4-дневных личинок севрюги.
В дальнейшем железа периодически выделяет скапливаюшийся секрет, причем усиление ее деятельности отмечается у молоди во время метаморфоза, а у половозрелых рыб—в преднерестовый период, до появления брачного наряда. Максимум активности совпадает с моментом овуляции.
Активность щитовидной железы меняется в течение жизни, постепенно падая в процессе старения, а также в зависимости от обеспеченности рыб пищей: недокорм вызывает усиление функции.
У самок щитовидная железа развита сильнее, чем у самцов, однако у самцов она более активна.
Щитовидной железе принадлежит важная роль в регуляции обмена веществ, процессов роста и дифференцировки, углеводного обмена, осморегуляции, поддержании нормальной деятельности нервных центров, коры надпочечников, половых желез. Добавление препарата щитовидной железы в корм ускоряет развитие молоди. При нарушении функции щитовидной железы появляется зоб.
Половые железы—яичники и семенники выделяют половые гормоны. Секреция их периодична: наибольшее количество гормонов образуется в период зрелости гонад. С этими гормонами связывают появление брачного наряда.
В яичниках акул и речного угря, а также в плазме крови акул обнаружены гормоны 17^-эстрадиол и эстерон, локализующиеся преимущественно в яйцеклетках, меньше — в ткани яичника. У самцов акул и лосося обнаружены дезоксикортикостерон и прогестерон.
У рыб существует зависимость между гипофизом, щитовидной железой и гонадами. В преднерестовый и нерестовый периоды созревание гонад направляется активностью гипофиза и щитовидной железы, а деятельность этих желез тоже взаимосвязана.
Поджелудочная железа у костистых рыб выполняет двойную функцию—железы внешней (выделение ферментов) и внутренней (выделение инсулина) секреции.
Образование инсулина локализовано в островках Лангерганса, вкрапленных в ткань печени. Он играет важную роль в регуляции углеводного обмена и синтеза белков.
Ультимобранхиальные (супраперибранхи-альные, или подпищеводные) железы обнаружены как у морских, так и пресноводных рыб. Это парные или непарные образования, лежащие, например у щук и лососевых, по бокам пищевода. Клетки желез секретируют гормон кальцитонин, который препятствует резорбции из костей кальция и таким образом не дает повышаться его концентрации в крови.
Надпочечники. В отличие от высших животных у рыб мозговое и корковое вещество разобщено и не образует единого органа. У костистых рыб они располагаются в разных участках почки. Корковое вещество (соответствующее кортикальной ткани высших позвоночных) внедрено в переднюю часть почки и носит название интерреналовой ткани. В нем обнаружены те же вещества, что и у других позвоночных, но содержание, например, липидов, фосфолипидов, холестерина, аскорбиновой кислоты у рыб больше.
Гормоны коркового слоя оказывают многостороннее влияние на жизнедеятельность организма. Так, глюкокортикоиды (у рыб обнаружены кортизол, кортизон, 11-дезоксикортизол) и половые гормоны принимают участие в развит!!и скелета, мышц, половом поведении, углеводном обмене. Изъятие интерреналовой ткани ведет к остановке дыхания еще до остановки сердца. Кортизол участвует в осморегуляцин.
Мозговому веществу надпочечников у высших животных у рыб соответствует хромаффинная ткань, отдельные клетки которой разбросаны и ткани почек. Выделяемый ими гормон адреналин воздействует на сосудистую и мышечную системы, увеличивает возбудимость и силу пульсации сердца, вызывает расширение и сужение сосудов. Увеличение концентрации адреналина в крови вызывает чувство тревоги.
Нейросекреторным и эндокринным органом у костистых рыб является и урогипофиз, находящийся в каудальной области спинного мозга и участвующий в осморегуляции, оказывающий большое влияние на работу почек.
Ядоносность и ядовитость рыб
Ядоносные рыбы имеют ядоносный аппарат, состоящий из шипов и ядовитых желез, расположенных у основания этих шипов (Mvoxocephalus scorpius в период икрометания) или в их желобках шипов и желобках плавниковых лучей (Scorpaena, Frachinus, Amiurus, Sebastes и др.).
Сила действия ядов различна: от образования в месте укола нарыва до расстройства дыхания и сердечной деятельности и смерти (в тяжелых случаях поражения Trachurus). В наших морях ядоносными являются морской дракончик (скорпион), звездочет (морская корова), морской ерш (скорпена), скат-хвостокол, морской кот, колючая акула катран), керчак, морской окунь, ерш-носарь, ауха (китайский ерш), морская мышь (лира), высоколучевой окунь.
При употреблении в пищу эти рыбы безвредны.
Рыбы, ткани и органы которых ядовиты по химическому составу, относятся к ядовитым и употребляться в пищу не должны. Они особенно многочисленны в тропиках. У акулы Carcharinus glaucus ядовита печень, у скалозуба Tetradon—яичники и икра. В нашей фауне у маринки Schizothorax и османа Diptychus ядовиты икра и брюшина, у усача Barbus и храмули Varicorhynus икра оказывает слабительное действие. Яд ядовитых рыб действует на дыхательные и вазомоторные центры, не разрушается при кипячении. У некоторых рыб ядовита кровь (угри Muraena, Anguilla, Conger, минога, линь, тунец, карп и др.). Ядовитые свойства проявляются при инъекции кровяной сыворотки этих рыб; они пропадают при нагревании, под действием кислот и щелочей.
Отравления несвежей рыбой связаны с появлением в ней ядовитых продуктов жизнедеятельности гнилостных бактерий. Специфический же “рыбий яд” образуется в доброкачественной рыбе (преимущественно в осетровых и белорыбице) как продукт жизнедеятельности анаэробных бактерий Bacillus ichthyismi, близкой к В. botulinus. Действие яда проявляется при употреблении сырой, в том числе соленой рыбы.
Похожие работы
... формированию крепких, здоровых и высокопродуктивных пород животных. Продуктивность животных Молочная, мясная, яичная, шерстная и рабочая продуктивность. Оценка животных по продуктивности. Учет рабочей производительности и оценка рабочих качеств животных. Сельскохозяйственных животных разводят главным образом для получения от них пищевых продуктов и сырья для перерабатываемой промышленности. ...

способлениями от избытка взвеси в воде у обитателей мутных вод являются уменьшение глаз (т. е. сокращение наиболее уязвимых участков поверхности тела) и усиленное выделение слизи, которая, осаждая муть, обеспечивает чистоту воды вокруг тела рыбы и таким образом улучшает условия дыхания. Глава I. Строение и некоторые физиологические особенности рыб · Строение и функции организма рыбы ...

... она связана с возможностью заражения при контактировании с забортной водой. Норма посадки производителей рыбца в прорези с полезным объёмом 30 м3 составляет 7000-7700 штук. Глава 6. Биологические основы акклиматизации рыбца Акклиматизация рыб и кормовых беспозвоночных является составной частью комплексных мероприятий по воспроизводству рыбных запасов и кормовых ресурсов в водоемах. Задачей ...


... . На Графике №2 отображена интересная особенность течения МКБ кошек и собак в 2002 году в зависимости от изменения погодных условий. Интересная особенность отмечена нами в процессе обработки статистических данных по 2002 году, которая нашла свое отображение в графике Сезонности уролитиаза кошек и собак за 2002г. На графике видно смещение пиков активности в группе кошек с марта на февраль и с ...
0 комментариев