Навигация
У межвидовых гибридов морфология петель такая же, как и у вида - донора Y-хромосомы
19361
знак
1
таблица
3
изображения
2. У межвидовых гибридов морфология петель такая же, как и у вида - донора Y-хромосомы.
Более детальный анализ показал, что гены ферментильности самцов локализованы в петлях.
1.Сначала были установлены корреляции между числом генов и петель. Затем, используя хромосомные – перестройки, установили прямое соответствие в их локализации. Так, фактор kl-5 соответствует петле А, поскольку и петля, и фактор располагаются между точками разрывов одних и тех же перестроек (см. рис. 3). Фактор kl- 3 расположен в петле B, ks-1 - в петле С.
2. При удалении делециями хотя бы одной петли самец становится стерильным.
После получения клонов ДНК из Y-хромосом дрозофил появилась возможность анализа молекулярной организации этой хромосомы. Общая длина петель составляет около 1000 мкм, или 1/12 всей длины ДНК в Y-хромосоме. Функции остальных 11/12 пока неизвестны. В состав ДНК Y-хромосомы входят два типа повторенных последовательностей.
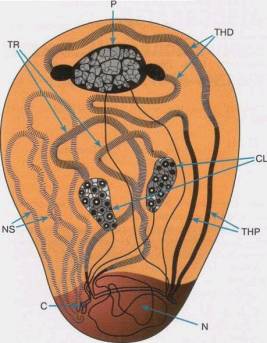
Рис. 3. Общий вид ядра спермотоцита у самца Drosofilia hydei (из [1], с.62). TR, P, THD, CL, THP, NS – названия петель, С – центромера, N - ядрышко
Вывод:
Таким образом, муха Дрозофила играет большую роль, как объект генетических исследований. Исследование ее генов принесло известность многим генетикам. Изучая гены Дрозофилы, ученые открыли много законов, таких как: закон Моргана - сцепленного наследования генов и закон Менделя. К тому же у Дрозофилы нашли много различных генов, отвечающих за наследование некоторых признаков. В последствии многие из этих открытий применялись к людям. И если бы не было этих мух, ученые еще долго бы не открыли законы наследования у людей.
Таблица 1.
| Свойства | Эухроматин | Гетерохроматин |
| Доля генома | 67% | 33% |
| Расположение в хромосомах | Плечи хромосом | В прицентромерных областях, вся Y-хромосом |
| Состояние компактности в клеточном | В ходе митотического и мейоти- | На протяжении всего клеточного цикла |
| Цикле | ческого делений | |
| Компактизующее влияние на приближен | Не оказывает | Участки эухроматина, приближенные к гетеро |
| ные участки хромосом (эффект положе | хроматину, также становятся компактными, | |
| ния мозаичного типа) | гены в них инактивируются | |
| Способность объединяться с другими | Не отмечена | Гетерохроматиновые участки объединяйте; |
| районами хромосом | образуя хромоцентры | |
| Образование хромосомных перестроек | Обычная частота обнаружения | Повышенная частота обнаружения |
| Расположение в клеточном ядре | По всему объему ядра | Главным образом на ядерной оболочке |
| Время синтеза ДНК в клеточном цикле | Первые 3/4 периода синтеза ДНК | Последняя половина S-периода. Завершени |
| в интерфазе (S-периода) | процесса репликации ДНК сильно задержано | |
| Дифференциальная окраска специфичес | Отсутствие окраски | Интенсивная окраска |
| кими красителями (С-окраска) | ||
| Фракции ДНК по степени повторенности | -90% уникальных последователь | Основная масса ДНК представлена высокопое |
| ностей и -10% умеренно повто | торенными фракциями, в меньшей степен | |
| ренных | умеренными повторами и совсем мало уникал! | |
| ных последовательностей | ||
| Наличие особых компактизующих бел | Почти отсутствует | Обильно присутствует по всему гетерохромг |
| ков, например белка НР1 | тину | |
| Варьирование количества материала в | Заметное варьирование не обна | Варьирование количества гетерохроматина un |
| хромосомах | ружено | роко представлено в каждой хромосоме |
| Генетическое содержание | Основная часть всех генов генома | Гены почти отсутствуют |
| локализована в эухроматине |
Литература:
«Соросовский Образовательный Журнал» том 6 №2 2000
И. Ф. Жимулев «Молекулярная и генетическая организация гетерохромотина в хромосомах дрозофилы».
Биология №14 1996.
А.А. Замятнин «Хромосомные матрицы, или Ода в честь плодовой мушки-дрозофилы»
Газета «Поиск» №11 2001Похожие работы


... состав мух птицехозяйств г. Семипалатинска, установить наиболее приемлемого в экологическом и биологическом отношении вида для использования как продуцента биомассы личинок используемого в качестве кормового белка. Сборы фаунистического материала и экологические наблюдения зоофильных мух проведены на территориях птицефабрик и малых птицехозяйств (ТОО КАСКЕ) Семипалатинского Прииртышья в период с ...
... самом деле, а каким нам его удобно видеть. И моделируем поведение Вселенной исходя из неких математических абстракций, в принципе доступных нашему воображению, но всё равно не укладывающихся по существу в «закон мухи». Даже ВЕЧНОСТЬ и БЕСКОНЕЧНОСТЬ мы понимаем по-своему – не так, как их задумал Господь, а так, как нам их позволяет видеть купированное восприятие на графике, лишённом областей свыше ...
... На смену бабочке-Музе, крылья которой и после смерти поражают красотой и гармонией запечатленного в них узора, приходит обессиленная, полусонная муха. В стихотворениях этих лет просматриваются прямые параллели между мухой и Музой поэта <…>» (Глазунова О.И. Иосиф Бродский: метафизика и реальность. СПб., 2008. С. 108). Эти параллели заданы созвучностью «пенья» насекомого и голоса Музы: Еле ...
... При цьому моделюються різні варіанти вирішення актуальних проблем, і кожна людина ладна самотужки приймати правильне на її думку рішення. РОЗДІЛ ІІІ. Філософсько-алегорична основа поетики роману Вільяма Голдінга «Володар мух» Роман В.Голдінга «Володар мух» вважають антиробінзонадою, антиутопією, філософсько-алегоричним романом. Твір доцільно розглядати саме як колективну антиробінзонаду, робі ...
0 комментариев