Навигация
Географическое распространение
44774
знака
3
таблицы
0
изображений
1. Географическое распространение.
Американские: бассейн Амазонки, Боливия, Эквадор, Перу, Колумбия до Южной Мексика.
Плейстоцен: Вест - Индия, Куба, Гаити, Ямайка.
Узконосые
Sectio Cattarhini
suprafamilia Cercopithecoidea
familia Cercopithecidae
subfamilia Cercopithecinae
Cercopithecus, Erythrocebus (красная мартышка-гусар), Cercocebus (мангобей), Papio (павианы), Mandrillus (мандрилы), Theropithecus (гелада), Macaca (=Macacus, макаки).
subfamilia Presbitinae (=Colobinae, тонкотелые)
Presbytis (лангур), Nasalis (носач, Калимантан, Суматра), Rhinopithecus (носатая обезьяна), Colobus (колобус, Африка), Simias, Pygathrix (тоноктел, пигатрикс, Индокитай).
Старый свет: тропический, субтропический район Африки и Азии.
В третичном периоде - много в Европе, сейчас сохранились только маготы (Гибралтар).
Большинство видов обитают в Африке:
мартышки - Египет - ЮАР, за исключением Сахары. До 20 видов.
Отдельный род - мартышки-гусары, обитают в Сенегале, обладают защитной окраской (красноватая шерсть на голове и спине).
Мангобеи - промежуточное положение между мартышками и павианообразными.
Павианообразные (собакоголовые):
собственно павианы
мандрилы (самые крупные из узконосых, неуклюжие, яркие мозоли)
гелады (рисунок в виде песочных часов на груди, морда напоминает таковую у мартышек).
Азиатские представители:
макаки 15-19 видов (типичные представители - макак-резус, яванский, свинохвостый, черный (раньше причисляли к павианам)).
хорошо переносят зиму, обитают до 40о северной широты. Наличие латентных луковиц (японский макак).
тонкотелые (много времени проводят в воде, теория водного происхождения человека).
В основном, азиатские представители (1 род в Африке - толстотел (=гвереца абиссинская =колобус ("изувеченный" - нет больших пальцев на руках)
Центральный род тонкотелы.
Носачи Nasalis, Калимантан; наличие у старых самцов хоботообразного хрящевого придатка.
Носатые обезьяны Rhinopithecus обитают в Китае, Вьетнаме; выступают носовые кости; обычно вогнутый профиль.
Обезьяны Нового Света – в основном древесные формы; у обезьян Старого Света очень разнообразные экологические ниши: медвежий павиан (чакма) живет в пустыне, ассамский макак и гималайский тонкотел – высокогорные районы.
павианы – четвероногие проноградные формы, опираются на всю поверхность подошвы и кисти. Кисть выполняет функции стопы, что приводит к увеличению гороховидной стопы – аналог пяточного бугра.
Все высшие приматы – дневные формы; исключение составляет Ateles. Характерен дневной тип сетчатки и стереоскопическое зрение.
Глазничная область
Формирование стенки глазницы: обезьяны Старого Света – глазничная поверхность лобной кости и большого крыла основной, слезно-носовой канал открывается внутри глазницы; обезьяны Нового Света – скуловая кость (единственная в своем роде структура, характерная для американских обезьян), слезно-носовой канал открывается снаружи глазницы.
Слуховая область
Обезьяны Старого света: тимпаническая пластинка, пирамиды височной кости, внутренняя сонная артерия входит в центр каменистой части; обезьяны Нового Света: слуховые капсулы. Собственный слуховой канал есть у всех обезьян и долгопятов.
Зубная система
Угол нижней челюсти округлен, оба отростка находятся примерно на одном уровне. Симфизеальное сращение половинок нижней челюсти: наличие базальной пластинки ("обезьянья пластинка"), занибающейся вниз и назад на нижнем краю нижнечелюстного симфиза.
Резцы верхних челюстей хорошо развиты, долотовидной формы (лопатообразная у шимпанзе); у задних премоляров тенденция к двухбугорковости.
Зубные формулы.
Американские обезьяны (как у лемуров):
| 2 1 3 3 | ||||
| 2 1 3 3 |
игрунковые – вторичная редукция моляров из-за маленьких размеров челюстей
| 2 1 3 2 | ||||
| 2 1 3 2 |
Обезьяны Старого Света:
| 2 1 2 3 | ||||
| 2 1 2 3 |
Форма коронки
Американские обезьяны: верхние моляры 3-4-бугорковые, нижние 4-бугорковые.
Узконосые: верхние моляры 4-бугорковые, нижние 4-5-бугорковые (как у человека).
Скелет
Позвоночная формула.
Низшие обезьяны: C7 T13 L6-7 S3-4
Человекообразные: C7 T13(12 у орангутана и человека) L3-5 S5-7 (укрепление крестца).
Хвост: американские обезьяны – не менее 14-15 позвонков (коата – свыше 30 позвонков); Старый Свет – редукция, преобладают бесхвостые формы, копчик 1-5.
Ключица: развита хорошо.
Лопатка: акромион развит лучше, чем у низших обезьян, но такого же типа, имеет боковое положение, лопаточный гребень параллелен позвоночнику, сочленовная впадина направлена вперед.
Грудина: палочковидная сегментированная, у человекообразных обезьян несегментированная, в виде плоской пластинки.
Таз: одинаковой формы у всех приматов, высокие крылья, длинное тело подвздошной кости, длинный лобковый симфиз; различия на уровне четвероногого хождения и прямохождения.
Конечности: плечевая кость – надмыщелковое отверстие характерно для низших американских обезьян; бедро – редко наличие третьего вертела.
Пальцевая формула: 3 4 5 2 1
Мозг
Более крупные размеры, чем у полуобезьян; мозг гирэнцефалический (размер зависит не только от уровня развития, но и от размера животного; у игрунки мозг маленький, более гладкий). Несколько борозд (обезьянья борозда – теменно-затылочная).
Органы чувств
Цветное зрение, в отличие от низших обезьян.
Редукция обоняния.
Замкнутый узор на пальцах, развитое осязание.
Ногти, у игрунок когти (вторично из-за древесного образа жизни).
Внутренние органы
Плацента гемохориального типа, дисковидная, опадающая.
Имплантация бластоцисты (у низших может просто прилегать).
Матка простая.
Одна пара грудных желез.
Кольца трахеи неполные.
Размножение
Сезонность размножения утрачена, обычно один детеныш (2-3 у игрунок, но редко, реже, чем у человека).
Тенденция к удлинению периода детства.
Половое созревание: 6 (гиббон) – 12-14 лет (♂ гориллы).
Общее удлинение жизненного цикла: 20-30 лет у низших обезьян, 45-50 у шимпанзе, до 120 у человека.
Систематика человекообразных обезьян.
subfamilia Hominoidea
familia Hylobatidae(гиббонообразные)
Hylobates (6 видов), Symphalangus (1 вид, разнопал)
familia Pongidae (орангутаны)
Pongo pygmaeus (1 вид):
Подвиды:
Pongo pygmaeus pygmaeus (орангутан калимантанский)
Pongo pygmaeus abelei (орангутан Абеля)
Pan troglodytes (шимпанзе)
Подвиды:
Pan troglodytes troglodites (шимпанзе обыкновенный)
Pan troglodytes verus (шимпанзе истинный)
Pan troglodytes schweinfurtii (шимпанзе швернунский)
Pan paniscus (шимпанзе карликовый)
Gorilla gorilla
Подвиды:
Gorilla gorilla gorilla ( горилла береговая, западный вид, Центральная Африка, 400-500 видов)
Gorilla gorilla manuema=graueri ( горилла равнинная, несколько тысяч видов, слабо изучены, есть предложения перенести в ранг вида)
Gorilla gorilla beringei (хорошо изучены)
Палеонтология высших приматов.
До сих пор ведутся споры о прародине высших приматов. Сушествует несколько теорий на этот счет.
Тарзиоидная теория.
Предположение о происхождении высших обезьян от неспециализированной группы долгопятов.
В Северной Африке еще в XIX веке обнаружены находки, показывающие специализацию долгопята(шорниус, тертониус).Датировка - 50 млн лет, т. е. уже в нижнем эоцене шла специализация по пути долгопятообразных форм.
Недавняя находка в Китае - эосимиас, крохотный примат с некоторыми чертами сходства с высшими приматами(по симфизу нижней челюсти). Датировка - 40-45 млн лет. Представлен несколькими видами. Есть более полные находки - цептепикус.
На основе этих данных видвинуто предположение о восточноазиатском центре происхождения высших приматов. Эта теория имеет место из-за раннегот возраста находок(слои этого возраста насыщены ими).
В Бирме обнаружены остатки Amphipithecus. Его характеризует прочный подбородочный симфиз, высокое положение подбородочного отверстия, пятибугорковые моляры на нижней челюсти. Это примитивный антропоид. Датировка – 40-44 млн лет.
Из вышесказанного можно сделать вывод, что отхождение линии высших приматов происходит в эоцене.
Северо-африканский центр.
Давние раскопки в Фаюке(недалеко от Каира), Египет.
Датировки - верхний-нижний эоцен.
Нижние горизонты - растительность открытого, сухого типа, здесь найдено очень мало приматов. По калий-аргоновому методу получается широкий размах датировок - 37-40 млн лет(если принимать последнюю цифру, то данная теория будет близка к азиатскому типу происхождения).
Представители(в основном, зубные фрагменты):
олигопитек (самый древний)
катопитек (верхний эоцен)
протеопитек (верхний эоцен)
Катопитек имеет сходство с низшими и американскими приматами в строении слуховой области, а в зубной области приближается к высшим ( 2 1 1 3). Олигопитек проявляет черты сходства с адаписами, а протеопитек - с американскими обезьянами (является прототипом общего предка).
Верхние горизонты - жаркий, влажный климат, появление лесных обезьян.
Выделяют 2 семейства: парапитеки и проплиопитеки.
Парапитеки.
Это примитивные прокатариновые обезьяны, имеющие черты сходства с американскими обезьянами. В целом занимают менее определенное положение, чем другое семейство.
Апидий (Apidium) - маленькая лесная обезьяна (челюсть – 35 мм), размером с саймири. Древесная лазающая форма, но одновременно и прыгающая(сросшиеся нижние концы берцовых костей). Слуховая область построена по типу американских обезьян, зубная формула - 2 1 2 3 (собственно парапитек, вариаций много, может быть 2 1 3 3, как у просимий), В общем похожи на дриопитеков.Среди парапитеков есть более крупные представители, похожие на карликовую мартышку. Они были найдены в 60-х годах прошлого столетия Саймосом Элвином и были вначале отнесены к церкопитекоидам. Это слабо специализированная группа и ее сходство с мартышкообразными можно отнести на счет одинаковых ниш обитания.
Проплиопитековые.
Примитивные гоминоиды.
Представители:
Египтопитек (зубы и череп - возможный предок)
Эолопитек (предок гиббона)
Собственно плиопитек
Американский центр происхождения (Южная Америка, бассейн Амазонки)
Первые находки - в Боливии и Аргентине, они гораздо моложе находок в других центрах.
Браниссела(Branissela boliviana) 26 млн лет, как и самые поздние северо-африканские формы). Предок бранисселы не найден.
Предполагаемые пути миграции: Африка – Южная Атлантика – Южная Америка (основная гипотиза). Восточный путь миграции (от азиатского центра): Восточая Сибирь – Северная Америка – Панамский перешеек – Южная Америка. Нет палеонтологического подтверждения этого пути миграции.
Следующая находка - рунейя из Техаса, с трубчатым слуховым проходом. В Африке кольцевидный слуховой проход долго сохранялся, только через многомлн лет заменился на трубчатый, а в Атлантике наоборот, так что обратные изменения навряд ли возможны.
Далее эволюция американских обезьян шла в условиях изоляции, причем очень однообразных(древесная среда). Они освоили разные экологические ниши, однако виды были ближе друг к другу, чем в Африке, такм образом можно предполагать консервацию черт общего предка(та же зубная формула, наличие прехаллиуса(есть у гиббона и американских обезьян), косточки, имеющей форму четверти сферы (prehallus) - простой сустав первого луча стопы, определяется по суставным площадкам).
В верхнем олигоцене, на 9 млн лет позднее бранисселы, обитали долихоцебус(череп похож на игрунковый) и тремацебус(ночные обезьяны).Тремацебус найден в Аргентине, в более поздних слоях, его выделяли в особый род Homunculus. Его и миоценовую обезьяну Homunculus miocen объединяют в семейство Homunculidae.
Представители: неосаймири( древние предшественники саймири), цебупитеция(близкие к современным питециям, т. е. К сакам). В Бразилии уже много представителей, близких к современным родам, что также наблюдается на Гаити, Ямайке( почти современные роды ревунов).
Древние мартышкообразные – специализированная боковая ветвь, может быть предком не только современных мартышкообразных, но и человекообразных обезьян. Иногда в эту группу включают даже египтопитека(30-26 млн лет, олигоцен). Палеонтологические данные – викториопитек, настоящая мартышкообразная обезьяна, датировка – 20 млн лет, жившая в Танзании, найдена близ озера Виктория, имела билофодонтные зубы, предположительно седалищные мозоли( определено по специфичным утолщениям на тазовых костях ). Биомолекулярные данные – девергенция от общего предка произошла по-видимому 25-20 млн лет назад, а в районе 20-16 млн лет – миграция мартышкообразных, широкое распространение по Европе.
17-18 млн (плиоцен и плейстоцен) появился мезопитек (Mesopithecus, обезьяна, похожая на тонкотела по черепу, на макаку – по посткраниальному скелету). По-видимому, была листоядной формой. Обладал округленным черепом со слабовыраженным рельефом, укороченным лицом (древесный тип черепа). Другие представители: долихопитек.
Гоминоидный африканский ствол.
Египтопитек – возможный предок обезьян до 20 млн лет. Предок дриопитеков. Западный ствол (верхний олигоцен) дриоморфов, предриопитеков (прежде всего по строению зубов). Черты сходства с американскими обезьянами: короткий наружный слуховой проход, прехаллюкс, надмыщелковое отверстие на плечевой кости, мозг, сходный с мозгом низших приматов ( малый объем, небольшие лобные доли).
Нижний миоцен – настоящие дриопитеки (проконсул). Датировка: 24-16 млн лет (до 12 млн). По африканским находкам, исчезают 12-10 млн лет назад. Найден в 1959 году. Это ювенильная особь с сильной деформацией черепа и выраженным прогнатизмом. По новым находкам проконсулов – лицевые соотношения размеров блмже к олобусам.
Кости очень тонкие, отсутствие рельефа на лобной кости, простая форма носового отверстия (у африканских представителей на носовом крае есть предносовой желобок), скуловые дуги неширокие, узкие обезьяние резцы, а клыки крупные. На нижней челюсти – наклон зубов по типу низших приматов (примитивный признак, напоминает туалетную щетку), почти не выражена обезьянняя площадка. Примитивный мозг: переднее положение центральной борозды, лобные доли не увеличены. Сам мозг по величине варьирует: малая форма – африканский, большая форма – ньярасский (167 куб.см, больше, чем у низших узконосых). Посткраниальный скелет: свободная центральная кость запястья(гиббон и американские обезьяны), хорошее развитие первого луча кисти(павиан), пропорции кисти нейтральны, наличие прехаллюкса, короткий слуховой проход. Интермембральный указатель: 87 (как у макак). По способу локомоции – четвероногие формы, жили не только в степях, но и в лесостепи, саванне и хорошо бегали. Место находки – Кения, на сегодняшний день известны сотни находок.
Хонвуд – общий предок щимпанзе и человеческой линии эволюции (“проконсулоидная гипотиза”, 60-е годы).
Сейчас описано много родов проконсулов:
Дендропитек (Лимнопитек). Обитал в в Восточной Африке, в нижнем миоцене, имел тонкое грацильное сложение (похож на гиббона).
Плиопитек. Европа, средний миоцен, потомок проплиопитека.
Наиболее полные находки – в так называемом Венском бассейне, находящимся на территории бывшей Чехословакии.
Предположительно, были филлофагами.
По внешнему виду представляли собой мозаику признаков, перестройка в направлении гиббонов еще не произошла. Позвоночная формула – по типу мартышкообразных, L 6-7 S 3. Грудина – понгидного типа, прехаллюкса у плиопитека нет (может, просто кость не найдена).
Новая находка (1999 год ) – человек тысячелетия (millenium man), кости, датированные 6 млн лет. Найдено бедро (предполагают двуногую форму) и плечевую кость (массивная, возможно, древолазы).
Южный Китай – лаккопитек (массивный), гиббонообразная обезьяна, 8 млн лет. Время отделения гиббоидной линии – 18-22 млн лет, 20-16 млн лет – многочисленные гиббоидные формы. В плиоцене и нижнем плейстоцене очень много гиббонов. Миграция: Африка – Европа – Ближний Восток – Восток. Происходит отделение классических, европейских дриопитеков.
Первые находки сделаны во Франции, близ города Сенгоден, в 1856 году. Была найдена нижняя челюсть и плечевая кость без диафизов. Этот представитель описан Ларте и назван фонтанов дриопитек.
По челюсти – это понгид: челюсть U-образная, параллельное расположение дуг зубов, на нижних молярах – специфический узор, называемый дриопитеков узор (часто встречается у шимпанзе и горилл, у человека редко), узор бороздки – Y-образный (Y-6 или Y-5), вместо Е-образного, увеличение моляров идет в сторону третьего ( у человекообразных обезьян М3 меньше М2). Присутствует мелкая морщинистость эмали. Клыки большие, имеются диастемы. У первого нижнего премоляра нет выраженной двубугорковой формы (секториальный тип первого премоляра).
Плечевая кость – небольшие размеры, слабый рельеф, общая грацильность. Датировка – 9-12,5 млн лет. Распространение – Австрия, Германия, Испания.
Европейские формы миоцена.
Грузинская находка, 1939 год, Гремяцкий – удабнопитек (2 зуба, маленький фрагмент премоляра и передний моляр, соединенные породой, 4-бугорковый моляр, увеличение числа корней: 4 у моляра, 3 у премоляра). Этот представительмиоцена или нижнего плейстоцена считается восточным вариантом дриопитека.
Ореопитек.
Были найдены элементы челюстного аппаратакоторые были сначала отнесены к мартышкообразным: удлиненные задние моляры, но ясно видны косые гребни (как у человекообразных).
Почти полный скелет был найден в Таскани (Италия), датировка – 12 млн лет, верхний миоцен. По размерам – как небольшой шимпанзе (1,10–1,20), 40 кг. Пропорции конечностей – брахиальный тип, длинные руки. Интермембральный указатель – 119, головка плечевой кости больше головки бедра. наличие косых гребней на молярах. Наблюдаются черты адаптации к двуногости – расширение тазовых костей (80). На пяточной кости хорошо виден пяточный бугор. И стопа, и таз – понгидного типа. На стопе первый палец противопоставлен остальным: седловидный сустав, но пятка хорошо обозначена. Они обитали в болотистых лесах южной Европы, передвигались по деревьям.
Череп. Большой мозг – 350 см3, коэффициент церебрации – 0,75-0,93, лицевой отдел укорочен, мозговой округлен. Тип – древесный.
Зубная система прогрессивная: нет диастем, маленькие клыки, двубугорковый первый премоляр, передняя часть лица округлена.
Адаптация к двуногости.
Выделяют в отдельное семейство ореопитецид.
Рудапитек.
Найден в Рудабании, Венгрия. Найдено два десятка индивидов, среди которых встречаются не только рудопитеки, но и плеопитеки.
Характерно выступание носовых костей, стенки носовой полости вертикальные. Предположительно могли передвигаться в выпрямленном положении. Датировка – примерно 8 млн лет.
Уранопитек (грекопитек).
Греческая находка – у. македонский.
Признаки сходства с аффарским австралопитеком: слабый надглазничный рельеф, малые клыки. Если это не самка, то черты действительно прогрессивные.
Восточный ствол дриопитеков.
Представлены находками в предгорье Гималаев. Названы сивапитеками (вначале назывались палеопитеки, 1870-е гг.).
Сивапитеки – орангоидные формы, датированные 14-13 млн лет, возможно, появились раньше. По биомолекулярным данным, линния орангутанов имеет датировки до 16 млн лет. Сейчас орангутан – единственный представитель этого ствола.
В 1930-х годах появились находки рамапитеков. Они отличались от сивапитеков более укороченной мордой, параболической зубной дугой (в отличии от U-образной), малыми клыками, небольшим первым премоляром (тенденция к двубугорковости), отсутствием диастем, толстой зубной эмалью (поздний признак). Они имели зубы наземного типа с толстой шейкой зубов. что говорило о питании жесткой травяной растительностью саванны, орехами.
Рамапитека пытались связать с человеческой ветвью эволюции, представить переходным звеном к австралопитеку. Впрочем. иммунологическая сыворотка разместила человека и шимпанзе дальше всех от рамапитека.
По зубной системе пытались судить о прямохождении, преобразовании кисти, развитом мозге, орудийной деятельности и даже о семейных группах.
Однако по форме зубной дуги нельзя четко различить сива- и рамапитека. Их стали считать особями мужского и женского пола соответственно.
Рамапитекоидная гипотеза продержалась до начала 70-х годов (с 60-х), но по мере поступления новых находок, ее сторонники потерпели фиаско. Были найдены многочисленные находки частей черепа и, в частности, челюстей в юго-западном Китае, Юннань, Люфенг. На их основе был описан род люфенгопитекус.
В 1935 году по нескольким зубам был описан род Гигантопитек, вид Г. Блейка. Это были очень большие, превосходящие человеческие в 6 раз, два третих и один первый моляры.
Сейчас сивапитеки, люфенгопитеки и гигантопитеки объединены в группу рамаморфов.
В 1956-58 годах в пещере китайской провинции Гуанси были найдены огромные скопления зубов и три нижние челюсти, по которым их назвали Гигантопитеки 1, 2, 3. Предположительно, это представители трех разных видов, хотя половой диморфизм также не исключен.
Датировка: 1-7 млн лет назад. Родоначальник предположительно гигантопитек из Беласпура. Он имел крупные размеры (1-3 метра), понгидные черты строения преобладали. Это был массивный (500 кг) вегетарианец (зубной камень с неорганическими тканями растений). Наблюдалось уменьшение передних зубов по отношению к заклыковым, клыки массивные, но из зубного ряда не сильно выступают, небольшая диастема.
Австралопитеки
Первая находка была сделана в 1924 году на территории ЮАР, в восточной части пустыни Калахари, недалеко от Таунга («львиное место»). Это был череп, эндокран и фрагмент нижней челюсти. Вначале считалось, что череп принадлежит обезьяне, что нашло отражение в названии – «австралопитек африканский» Australopithecus africanus (южно-африканская обезьяна, Дарт). Однако, обьем мозга найденного черепа превышал мозг шимпанзе, соотношение мозгового и лицевого отделов черепа также отличалось от обезьяньего. У данного индивида прорезался первый моляр, поэтому приблизительный возраст составляет 5-6 лет (с учетом того, что у человекообразных обезьян первый моляр прорезается раньше, от 3 лет, возраст особи определили как 3-5 лет).
В 30-е годы появились новые находки в карстовых пещерах Стеркфонтейн (более 60 различных объектов), Сварткранс, Кромдраай. Это были остатки посткраниального скелета, в том числе тазовые кости, по которым была определена адаптация австралопитеков к двуногости.Тазовые кости очень похожи на кости современного человека, но они миниатюрнее (сравнение по тазовым костям пигмея), есть ямка подвздошной кости, S-образный подвздошный гребень, укрепление крестцово-подвздошной области (ушковидная поверхность), большая вертлужная впадина, дорзальный наклон пластинки крыла, обеспечивающий большую седалишную вырезку. При моделировке краев кости были выявлены элементы, связанные с фиксацией ноги, разгибанием в тазобедренном суставе. Внутренняя структура кости по методу компьюторной томографии также соответствует двуногости, но походка не была идентична походке шагающего человека. Австралопитек передвигался мелкими шажками и рысцой.
Найденные кости в пещерах различались по массивности, в связи с чем австралопитеков стали делить на плезиантропов и парантропов. Австралопитеки грацильного типа (плезиантропы "Plesianthropus"), более близкие к современному человеку; это находки в Стеркфонтейн и Кромдраай, женская тазовая кость (крылья сильно отогнуты, широкая седалищная вырезка). Парантропы Paranthropus (пещера Сварткранс) – более массивные кости, сагитталный гребень сильно выражен. Для всех австралопитеков характерно уменьшение резцов и клыков, крупные моляры, толстая эмаль. Внутренняя часть рельефа челюсти варьирует: подбородка нет, есть лишь неьольшое утолщение подбородочного симфиза. У человека есть вторая ость (spina mentalis), у обезьян (и у парантропов) – только ямка. Сочленовная впадина близка к шимпанзоидному типу. Лопатка обеспечивала большую свободу движений, свойственную брахиаторам.
Рядом со скелетами были найдены элементы остеодонтокератической культуры (обломки рогов, костей животных), назначение которых не совсем ясно, они могли использоваться для различных целей. Также плезиантропам приписывли применение огня (почернение почвы, считавшиеся вначале остатками золы, позже выяснили, что это соли марганца), названный поэтому Прометеем ("Australopithecus prometheus"); сейчас называют греческим типом африканского австралопитека A. africanus (пещера Макапансгат).
По данным палеоневрологии (изучение эндокрана) картина сети сосудистой оболочки различается у массивного (сильное развитие затылочно-краевого венозного синуса) и у грацильного типов (ближе к человеческому). Предположительно, это адаптивный признак с учетом среды обитания (у детей система охлаждения мозга – как у массивного).
В 30-50 годы о виде австралопитек сложились две основные точки зрения:
род Australopithecus: A. africanus (грацильная форма), Макапансгат, Стеркфонтейн, возможно, Таунг. Датировки: современники питекантропов, 1 млн лет (биостратиграфия). Массивных австралопитеков выделяют в другой род – парантропы (Paranthropus).
Нет достаточного обоснования межродового уровня. Датировка – 2-1 млн лет.
Находки Восточно-Африканского региона.
1959 год. Экспедиция Льюиса Лики. Танзания, Олдувайское ущелье, дно пересохщего солоноватого озера. Самый древний слой – 2 млн-1,7 млн лет.
В 1959 году были найдены остатки посткраниального скелета (голень) и череп, принадлежавшие разным существам. Череп датировался приблизительно 1,8 млн лет. Рядом находились каменные орудия (олдуван).
Восточно-африканский человек ("Zinjanthropus boisei"). Мужчина, 18 лет, гориллообразная внешность, мощные скуловые дуги, сильный рельеф. Кости голени – тонкие, грацильные.
Новые находки – Кения, Эфиопия., 70-е годы, более 200 останков. Западная Туркана – "черный череп", особый вид, А. aethiopicus, возможный предок современного человека.
Грацильный тип тоже встречается: A. afarensis (73-76 годы), Эфиопия, около 40 индивидов.
Самые полные останки – Люси, 1974 год. Женщина, 20-30 лет, форма таза, как у плезиантропа, гипертрофирован по ширине, невозможность ротации головки плода.
Похожие находки были сделаны не только в Эфиопии (Хадар, бывшее озеро). Это двуногие формы (плечебедренный указатель – 84-85), на лопатке – косое положение лопаточной впадины, изогнутость фаланг пальцев ("древесность"), на позвонках – разрастание костной ткани (не патология). Длинноногая форма – шагающая походка.
Мозг: две тенденции морфотипа (объем, в см3): грацильные – 440-450 (Южная Африка), 414 (Восточная Африка), массивные – 514-516. Рост – 1-1,6 м. Коэффициент церебрализации (соотношение веса мозга и тела): австралопитек – 1,1-1,5, современный человек – 2,6-2,7, шимпанзе – 0,9.
Другие виды австалопитеков являются недостаточно хорошо изученными.
A. ramidus
Найден в начале 90-х годов, в Эфиопии (среднее течение реки Аваш). О локомоции нет точных сведений, предположительно, двуногий. Палеонтологами выделяется в отдельный род – Ardipithecus, хотя первоначально они были объединены с австралопитеками. Датировка – 4,1 млн лет.
A. anamensis
Таксономический статус не ясен. Найдена большая берцовая кость. Предположительно, двуногая форма. Возраст – 4.2-3.8(3.5) млн лет. Это самый ранний их представителей австралопитеков.
Родоначальником массивных австалопитеков восточной Африки считают А. aethiopicus (подобные находки известны из Эфиопии). Был найден череп там же , где и A. boisei, но в более раннем слое (возраст - 2,5 млн лет).
Кения, озеро Баринго. "Человек тысячелетия" Orrorin tugenensis. К старой находке коренного зуба (гоминоидного типа, близкого к гоминидам), датированной 6 млн. лет, прибавились плечевая и бедренные кости, зубы (клыки небольшие, коронка высокая – прогрессивные признаки). Найдено бедро (предполагают двуногую форму) и плечевую кость (массивная, возможно, древолазы). Благодаря этим находкам возникло предположение, что род Homo, возможно, более древний, чем считалось раньше (приравнивается по времени к австралопитекам).
Кениантроп (Kenyanthropus platyops)
Сочетание примитивных признаков и черт, сближающих с родом Homo, на черепе. Лицевой скелет напоминает KNM-ER 1470 (Homo rudolfensis), в отношении которого не определен таксономический ранг. Возраст – 1,9 млн лет. Сначала причислили к H. habilis (олдувайским), потом стали рассматривать как кенийский вариант (продвинутый). Это первые общепризнанный предок Homo. Зубы: верхние моляры трехбугорковые. Фрагмент нижней челюсти: передняя часть зубной дуги уменьшена. Относят к ранним, примитивным плейстоценовым формам.
H. habilisПервые находки – 1960 год, таксон был выделен в 1964. Таксон постоянно изменяется, новые находки неодназначны. Первый хабилис – ювенильная форма, 11-12 лет, рядом орудия.
К олдувайским хабилисам относят череп KNM-ER 1813, KNM-ER 1470 выделен из вида Homo habilis и назван "человек рудольфский" (Homo rudolfensis).
Аналогичные находки сделаны в Азии: пещера Лонггуппо (Китай), датировка – 2-2,8 млн лет. Это нижняя челюсть с молярами, похожими на аналогичные зубы у habilis, а также 2 каменных артефакта со следами возможной обработки. Эта культура названа преолдувай
Похожие работы
... из Африки. Люди современного физического типа появились в Восточной Азии 75-150 тыс лет назад. Молекулярно-генетические данные также не подтверждают эту гипотезу. Факторы антропогенеза Осмысливая происхождение человека, Ч. Дарвин придавал большое значение таким факторам антропогенеза, как изменчивость телесных и психических свойств предков человека, использование ими тех или иных органов, ...
... , венцом эволюции материи - космогенеза – является антропогенез. На определенном этапе антропогенеза возникает ноосфера - мыслящая оболочка планеты с отделением мыслящего духа от своей материальной основы (Шарден Тейяр П., 1965; Вернадский В.И., 1977; Алексеев В.П., 1984). Согласно концепции крупного отечественного философа Н.Н. Моисеева, Вселенная представляет собой суперсистему, включающую в ...
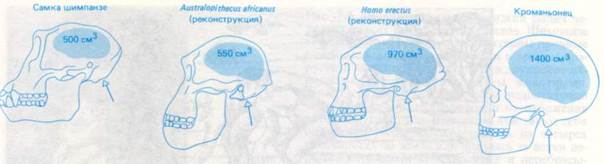
... - известен только по нескольким зубам и фрагментам челюсти, возраст которых определяется в 9-14 млн. лет назад. Неизвестно, был ли он прямоходящим. Рис. 3 Австралопитек. Начальные стадии антропогенеза начались 2,5 - 3 млн. лет назад с возникновением австралопитека африканского (Australopithecus Africanus), который был прямоходящим, обладал развитым мозгом и изготавливал орудия труда. Однако, ...
... отбора" (1859 г.) этой формулировки нет. Даже спустя 12 лет, в работе "Происхождение человека и половой отбор", осмотрительный Дарвин пишет лишь об эволюции вообще, совершенно не касаясь "промежуточного звена". О происхождении же человека от обезьяны заявили в 1863 г. дарвинисты Фохт, Гексли и Геккель. Именно Геккель говорил о "недостающем звене" между высшими приматами и человеком, имея в виду ...
0 комментариев