Навигация
Виды биологической продуктивности лесов
62874
знака
12
таблиц
4
изображения
2.7. Виды биологической продуктивности лесов
Потенциальная продуктивность является одной из главных показателей эталонных насаждений, характеризуется такими показателями как видовой состав. Хозяйственная ценность, объединяемая в итоге в понятие «хозяйственной продуктивности». Под понятием «потенциальной продуктивности» понимается максимально возможная продуктивность для данных климатических и почвенных условий, будь то первичная биологическая продуктивность (т/га) или продуктивность запаса стволовой древесины (м3/га). Обычно последняя наиболее существенная составляющая первичной продуктивности является предметом изучения лесоводов.
Методы изучения потенциальной продуктивности по В.С. Чуенкову можно подразделить на три основные группы: лесоводственно-таксационные, лесотипологические и климатологические. Наиболее распространены лесоводственно-таксационные, с помощью которых изучают закономерности строения и роста насаждений, являющееся основой последующего моделирования этих параметров. Итоги многолетних исследований продуктивности лесов обобщены на Всесоюзной конференции по формированию максимально продуктивных эталонных насаждений, которая проводилась в Каунасе в 1979 году. Большинство работ, представленных на конференции выполнено именно лесоводственно-таксационными методами. Эти методы применимы для конкретых условий произрастания, в то время как климатологические методы оценки применимы для крупных территорий. Климатологические методы определения продуктивности основаны на зависимости потенциальной продуктивности от климатологических факторов. Обычно несколько гидротермических показателей климата путём арифметических операций комбинируют в один комплексный показатель, называемый климатическим индексом прироста, который указывают с годичным приростом древостоя. Например, климатический индекс ‘ I ’ Дж. Векка включает в себя осадки ‘ N ’, среднюю температуру воздуха ‘ T ’ и число дней с осадками менее 0,1 мм ‘ n ‘ в течении мая-июля, число дней в году с положительными температурами ‘ Z ‘ и имеет следующий вид:
i = ( N/T+10 * n/92) * (Z-60/100).
С. Патерсон вводит в 1956 году более удобные в использовании средние годовые показатели климата, которые можно найти в климатических справочниках.
I = Tv/Ta * N/1 * G/12 * E/100,
где Tv – средняя температура самого тёплового месяца;
Ta – амплитуда температур самого тёплого и самого холодного
месяцев;
N – годовое количество осадков;
G – продолжительность периода с температурами более +7С;
Е – фактор Миланковича равный Rp/Rs, где:
Rp – суммарная радиация на полюсе;
Rs - суммарная радиация в данном месте.
Индекс Патерсона часто используется в исследовании потенциальной продуктивности лесов.
Годичная продуктивность лесных насаждений определяется путём взятия модельных деревьев, у которых оценивается масса разных фракций (древесина, кора, ветви, листва, генеративные органы, корни).
2.8. Основные экологические и биологические факторы, определяющие продуктивность лесов (на примере лесов севера)
Всестороннее выяснение взаимосвязей и закономерностей роста и развития древесных растений в естественных фитоценозах даёт возможность проектировать и создавать биогеоценозы оптимального состава и структуры, способствующие повышению продуктивности древостоев.
Сосновые и еловые леса наиболее распространенной зеленомошной группы типов леса в условиях северной и средней тайги северо-востока европейской части РФ образуют, как правило, смешанные по составу древостои. Для них характерна невысокая полнота (0,5-0,7), IV, V классы бонитета. В насаждениях естественного происхождения наблюдается большая дифференциация деревьев по возрасту, диаметру и высоте. Эта особенность заметно выражена в еловых фитоценозах. В каждом типе соснового и елового леса создаются характерные эдафические, фитоклиматические условия. В связи с этим из сложного комплекса факторов внешней среды необходимо выявить и дать количественную характеристику тем из них, которые оказывают наиболее существенное прямое и косвенное влияние на жизнедеятельность растений и интенсивность образования органической массы биогеоценоза.
Солнечная радиация является наиболее важным экологическим фактором. Суммарная её величина, поступающая к пологу древостоев северной и средней тайги (390-420 кал/см2*сут), близка к наблюдаемой в южной тайге и даже в зоне широколиственных лесов, что обусловлено продолжительностью светового дня. Следовательно, на севере таежной зоны условия поступления солнечной радиации в течении вегетационного периода не оказывает значительных ограничений на формирование более продуктивных насаждений. Но следует учесть, что малая продолжительность вегетационного периода полностью не компенсируется длительностью дня. Благодаря большим межкроновым просветам в хвойных лесах солнечная радиация хорошо проникает вглубь полога, что приводит к большему прогреванию верхней половины кронового пространства. Это является важной специфической чертой, свойственной лесным биогеоценозам, развивающимся в условиях общего недостатка тепла.
Пропускание солнечной радиации древесным ярусом хвойных фитоценозов северной тайги составляет 29-44%, что в 3-4 раза выше по сравнению с ельниками и сосняками зоны елово-широколиственных лесов. Полог сосняка пропускает суммарную радиацию на 10-15%, а фотосинтетически активной радиации (ФАР) – в 1,5-2 раза больше, чем темнохвойные фитоценозы. Высокая светопроницаемость полога сосновых лесов позволяет такой теневыносливой породе, как ель, успешно расти под пологом сосны. В результате двухъярусные сосново-еловые древостои средней тайги на плакорах значительно больше поглощают солнечную радиацию (80%) и приближаются по этому показателю к ельникам Московской области. Коэффициенты поглощения ФАР сосняками и ельниками зеленомошной группы типов леса северной тайги Коми АССР составляют 0,63 и 0,80, что соответствует данным В.А. Алексеева (1975) для Мурманской и Архангельской областей, где эти показатели равны 0,60 и 0,79.
В северотаёжных лесах остаточное радиационное тепло почти в равной мере используется на суммарное испарение деятельного слоя и на турбулентный теплообмен. Эта особенность хвойных фитоценозов является следствием низких температур воздуха, меньшей интенсивности физиологических процессов, главным образом транспирации. В условиях же Валдая и Московской области основную долю остаточного радиационного тепла составляет расход на суммарное испарение леса.
Один из основных экологических факторов, отрицательно влияющих на рост и развитие древесной растительности, - невысокая температура воздуха, которая определяет прогрев почвы и интенсивность транспирации. Тем самым создается неблагоприятный режим температуры, влажности и аэрации в ризосфере лесных фитоценозов. Период, когда возможны активный рост и накопление органической массы надземными органами древостоев, короткий и составляет 30-40 и 70-80 дней соответственно в северной и средней тайге. В условиях северной тайги по сравнению со средней в дневное время суток в 4 раза чаще наблюдаются температуры воздуха ниже 10С. После весеннего снеготаяния почвы прогреваются медленно. В северной тайге глее-подзолистые почвы сосняка чернично-зеленомошного на глубине 0,2м нагреваются до температуры 5С лишь во второй декаде июня. Температура торфянисто-подзолисто-глееватых почв ельника чернично-зеленомошного на этой глубине достигает той же величины к началу июля. В условиях средней тайги температура подзолистых почв в лесах зеленомошной группы типов леса несколько выше, но они также характеризуются небольшими тепловыми ресурсами. Температурные условия, при которых возможен рост корней, на глубине 0,2м в сосняках наступает к началу июня, а в ельниках – во второй половине июня. Более благоприятный режим для развития древесных растений наблюдается только в самых верхних горизонтах почвы и в течении очень короткого периода. Следовательно, недостаточное прогревание почв препятствует освоению корнями нижней части почвенной толщи, а интенсивность поглощения питательных веществ при низких температурах невелика.
Особенно отрицательно сказываются на жизнедеятельности деревьев ежегодно повторяющиеся резкие похолодания в разгар вегетационного периода (обычно в июне), связанные с поступлением арктических воздушных масс. В эти периоды минимальная температура воздуха в ночные часы в кроновом пространстве может опускаться в северной тайге до –2-4С, а верхнего корнеобитаемого слоя почвы – до –0,5-1,0С. В средней тайге обычно понижение температур воздуха в кронах отмечается до 2-4С, а в верхнем корнеобитаемом горизонте почвы – до 1,5-2С. Такие температурные условия вызывают резкое снижение прироста, а в отдельные годы даже полную приостановку ростовых процессов надземных и подземных органов древесных растений. Особенно сильно резкие перепады температур сказываются на жизнеспособности ели, вызывая массовое отмирание молодых побегов.
Почвы лесных фитоценозов данной группы типов леса очень кислые, не насыщены основаниями. Бедны элементами питания и оглеены. Формирование их в лесных фитоценозах исследуемого региона происходит в условиях повышенной влажности. Влаги в почве достаточно для поддержания жизнедеятельности растений, а в отдельные годы, особенно в условиях северной подзоны тайги, избыток её угнетает ростовые процессы деревьев. В ельниках зеленомошных умеренно влажными были органогенный слой и подзолистый горизонт. Вниз по профилю, начиная с глубины 20см, почва во все сроки наблюдений находилась в состоянии переувлажнения, что приводит к нарушению её кислородного режима и снижению биологической активности почвы, также наблюдаются негативные отклонения в энергетике, синтетической деятельности и обмене веществ корневых систем. Все перечисленные нарушения в почве и метаболизме корней в условиях избыточного увлажнения отрицательно сказываются на росте и развитии корневых систем древесных растений севера.
В северо-таежных хвойных фитоценозах основная масса корней древесных пород распространяется до 30. а в среднетаежных - до 50см. Это явление особенно сильно проявляется в условиях северной подзоны тайги, где в верхнем 10-15 сантиметровом слое сосредоточено более 90% корней. Сосущие корни хвойных древесных пород образуют очень компактные разветвления.
Линейный рост вегетативных органов зависит от метеорологических условий текущего года. Решающим фактором является количество тепла, получаемое растением. Видимый рост побегов, корней у сосны и ели начинается поздно: в условиях северной тайги – в 1-2 декаде июня, в отдельные годы – в начале июля, в условиях средней тайги – во 2-3 декаде мая, протекая интенсивно и в сжатые сроки. Максимальный прирост и накопление органической массы древостоем в северной тайге происходят в конце июня – первых двух декадах июля, в средней тайге – в июне, первой половине июля. В это время деревья испытывают наибольшую потребность в минеральных веществах.
Большинство исследователей, изучавших рост подземных органов древесных растений, считает, что для преобладающих видов их характерны два периода максимального роста (весенний и осенний) и два периода покоя, или минимума роста (летний и зимний).
В годы обильных урожаев усиленный приток ассимилянтов к развивающимся шишкам и семенам ограничивает питание камбия, что вызывает сокращение годичного прироста древесины. Считается, что наступление неурожайных лет вслед за годом обильного урожая связано с истощением запасных питательных веществ, на восстановление которых необходим определенный срок. Этим объясняется явление так называемой периодичности урожаев, хотя можно утверждать, что циклический характер плодоношения хвойных определяется как внешними факторами, так и биологическими особенностями вида, и в первую очередь размещением женских шишек и мужских колосков на материнском вегетативном побеге.
В дальнейшем одним из мероприятий, направленных на повышение продуктивности северных ельников, может стать отбор стерильных и малоурожайных гибридных форм с целью введения их в состав искусственных насаждений наряду с другими быстрорастущими и урожайными фенотипами.
Исследование продуцирования органического вещества растительными сообществами представляет значительный интерес как основа для оценки сложных энергетических связей в экосистемах. В результате весьма неблагоприятного сочетания и взаимодействия основных экологических факторов и напряженного протекания биологических процессов в лесных фитоценозах севера таежной зоны развиваются древостои низкой продуктивности. Прирост и общие запасы биомассы древесной растительности в 1,5-3 раза меньше, чем в более южных районах лесной зоны. Полученные данные И.С. Мелеховым (1970) показали, что в условиях северной подзоны тайги сосновые насаждения зеленомошной группы типов леса производительнее, чем еловые. Сосновые древостои по сравнению с еловыми используют лучистую энергию солнца более эффективно, даже в случае, когда они моложе по возрасту, органическая масса их больше, чем у ельников. В еловых насаждениях накапливается больше хвои (листьев) и стволовой коры. Биомасса сосняков отличается более высоким содержанием качественно ценного компонента – древесины ствола. Следовательно, в зеленомошных типах леса северной подзоны тайги наиболее целесообразно воспроизводство сосновых лесов, чем еловых. Необходимо также отметить, что двухъярусный сосново-еловый древостой средней подзоны тайги отличается довольно высоким коэффициентом поглощения и использования ФАР и относительно большим накоплением органического вещества.
Поиск путей повышения продуктивности северных лесов является узловой задачей таёжного лесоведения. В сложных климатических условиях Европейского северо-востока познание биологических особенностей роста и развития основных лесообразующих пород будет иметь ключевое значение при осушении и удобрении лесов или создании прочной лесосеменной базы на селекционно-генетической основе.
Из курса биогеографии мне известно, что:
экологический оптимум вида – это наибольшая продуктивность вида при условии оптимального соотношения экологических факторов.
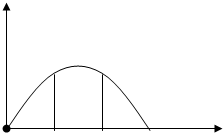
Opt
Min Max
Фитоценозтический оптимум вида – это наибольшая продуктивность вида в условиях конкуренции в растительном сообществе), при оптимальном соотношении экологических факторов.
Похожие работы
... квартал №46. Самой богатой почвой - перегнойно-глеевая песчаная на флювиогляциальных песках, которая занимает наибольшую площадь из всех представленных у нас почв. 5. Влияние почвенного плодородия на продуктивность лесообразующих пород Рациональное ведение лесного хозяйства определяется экологическими требованиями пород лесообразователей к почвенно-грунтовым условиям, обеспечивая при этом ...
... (возвращаемых с спадом) зольных элементов и азота (в кг/га). В целях унификации принята десятибалльная шкала числовых показателей. 3 Лесной тип биологического круговорота 3.1. Различия степных и лесных экосистем Энергетические затраты на первичную продукцию и в целом на биогеоценотические процессы возрастают от среднетаежных растительных формаций к подтайге и широколиственным лесам, а ...
... , песчаные. Каменисто-щебенчатые Вид Сходство доминирующих урочищ Западно-сибирские равнинные возвышенные степные с разнотравными степями на черноземах легкосуглинистых Пример результата классификации ландшафтов южного Подмосковья (по В. А. Николаеву): отдел — наземные; разряд — бореальные; подразряд — умеренно континентальные; семейство — восточно-европейские; класс — ...
... , являющихся защитными и выполняющих водоохранные, почвозащитные и другие полезные функции. В первую очередь резерваты были отведены на территории Катон-Карагайского государственного национального природного парка ЛГР №12-17. Ведение хозяйства в лесных генетических резерватах осуществляется в соответствии с Типовыми положениями, Лесным кодексом Республики Казахстан и другими нормативными актами. ...
0 комментариев