Навигация
УРАЛЬСКАЯ ГОСУДАРСТВЕННАЯ МЕДИЦИНСКАЯ АКАДЕМИЯ
Кафедра биоорганической и биологической химии
КУРСОВАЯ РАБОТА НА ТЕМУ:
Биологическое окисление.
Исполнители: студентки
педиатрического
факультета 223 группы
Заруба Н.С., Чащина Е.Е.
Руководитель: доцент,
к.м.н. Трубачев С.Д.
Рецензент:
Екатеринбург 2002.
Содержание.
I. Введение………………………………………………………………...3
II. Общие представления о биологическом окислении.
Окислительно-восстановительные системы и потенциалы……..3
III. Пути использования кислорода в клетке……………………………...5
- Оксидазный путь использования кислорода. Митохондрии.
Ферменты, их локализация и значение в процессах окисления…….5
IV. Этапы утилизации энергии питательных веществ…………………...6
V. Окислительное фосфорилирование……………………………………9
- Хемиосмотическая теория Митчелла……..……….………………..9
- Редокс – цепь окислительного фосфорилирования………………10
VI. Цикл Кребса…………………………………………………………21
- Открытие ЦТК……………………………………………………..22
- Реакции, ферменты. Регуляция…………………………………...23
VII. Макроэргические соединения и связи……………………………...29
VIII. Витамин РР. Участие в процессах окисления…………………….30
IX. Микросомальное окисление…………………………………………31
- Монооксигеназные реакции………………………………………31
- Диоксигеназные реакции………………………………………….32
- Цитохромы…………………………………………………………32
X. Пероксидазный путь использования кислорода…………………..33
XI. Ферментативная антиоксидантная защита…………………………34
- Супероксиддисмутаза, каталазы, пероксидазы………………….34
XII. Неферментативная антиоксидантная защита………………………35
- Витамины С, Е и Р…………………………………………….…...35
XIII. Заключение…………………………………………………………..38
XIV. Список литературы…………………………………………………..39
Введение.
В химии окисление определяется как удаление электронов, а восстановление - как присоединение электронов; это можно проиллюстрировать на примере окисления ферро-иона в ферри-ион:
Fe2+-e → Fe3+
Отсюда следует, что окисление всегда сопровождается восстановлением акцептора электронов. Этот принцип окислительно-восстановительных процессов в равной мере применим к биохимическим системам и характеризует природу процессов биологического окисления.
Хотя некоторые бактерии (анаэробы) живут в отсутствие кислорода, жизнь высших животных полностью зависит от снабжения кислородом. Кислород, главным образом, используется в процессе дыхания – последнее можно определить как процесс улавливания клеточной энергии в виде АТФ при протекании контролируемого присоединения кислорода с водородом с образованием воды. Кроме того, молекулярный кислород включается в различные субстраты при участии ферментов, называемых оксигеназами. Многие лекарства, посторонние для организма вещества, канцерогены (ксенобиотики) атакуются ферментами этого класса, которые в совокупности получили название цитохрома Р450.
Гипоксические нарушения метаболизма клетки занимают ведущее место в патогенезе критических состояний. Главную роль в формировании необратимости патологических процессов приписывают крайним проявлениям расстройства клеточного метаболизма. Адекватное обеспечение клетки кислородом является основным условием сохранения ее жизнеспособности.[12,1992]
Введением кислорода можно спасти жизнь больных, у которых нарушено дыхание или кровообращение. В ряде случаев успешно применяется терапия кислородом под высоким давлением; следует однако отметить, что интенсивная или продолжительная терапия кислородом под высоким давлением может вызвать кислородное отравление.[2,1994]
При написании данной работы перед нами стояла цель: изучить биологическое окисление и его значение в жизнедеятельности клетки и организма в целом. Для этого мы рассмотрели:
- использование кислорода клеткой;
- источники энергии клетки – цикл лимонной кислоты (цикл Кребса), окислительное фосфорилирование;
- микросомальное окисление;
- антиоксидантную защиту
Общие представления о биологическом окислении.
Окислительно-восстановительные системы и потенциалы.
Источник энергии, используемый для выполнения всех видов работ (химической, механической, электрической и осмотической) – это энергия химической связи. Высвобождение энергии углеводов, жиров, белков и других органических соединений происходит при их окислительно-восстановительном распаде. Высвобожденная энергия затрачивается на синтез АТФ.
Изменение свободной энергии, характеризующее реакции окисления и восстановления, пропорционально способности реактантов отдавать или принимать электроны. Следовательно, изменение свободной энергии окислительно-восстановительного процесса можно характеризовать не только величиной DG0', но и величиной окислительно-восстановительного потенциала системы (Ео). Обычно окислительно-восстановительный потенциал системы сравнивают с потенциалом водородного электрода, принимая последний за ноль, 0В при рН=0. Однако для биологических систем удобнее использовать окислительно-восстановительный потенциал при рН=7,0 (Ео'); при таком рН потенциал водородного электрода равен -0,42В.[10,1993]
Пользуясь таблицей 1, можно предсказать, в каком направлении пойдет поток электронов при сопряжении одной окислительно-восстановительной системы.
Таблица 1. Стандартные потенциалы некоторых окислительно-восстановительных систем.[10,1993]
| Система | Ео¢/ Вольт |
| Кислород/вода | +0,82 |
| Цитохром a: Fe3+/Fe2+ | +0,29 |
| Цитохром с: Fe3+/Fe2+ | +0,22 |
| Убихинон: окисл./восстан. | +0,10 |
| Цитохром b:Fe3+/Fe2+ | +0,03 |
| Фумарат/сукцинат | +0,03 |
| Флавопротеин: окисл./восстан. | -0,12 |
| Оксалоацетат/малат | -0,17 |
| Пируват/лактат | -0,19 |
| Ацетоацетат/гидрооксибутират | -0,27 |
| Липоат: окисл./восстан. | -0,29 |
| НАД+/НАДН | -0,32 |
| H+/H2 | -0,42 |
| Сукцинат/альфакетоглутарат | -0,67 |
Пути использования кислорода в клетке.
Существует три пути использования кислорода в клетке, которые характеризуются следующими реакциями:
1) оксидазный путь (90% поступившего кислорода восстанавливается до Н2О при участии фермента цитохромоксидазы)
02+4е+4Н+ → 2Н2О
2) оксигеназный путь (включение в субстрат одного атома кислорода - монооксигеназный путь, двух атомов кислорода -диоксигеназный путь) -монооксигеназный путь
-диоксигеназный путь
3) свободно-радикальный путь (идет без участия ферментов и АТФ не образуется).
Оксидазный путь использования кислорода. Митохондрии. Ферменты, их локализация и значение в процессе окисления.
Митохондрии справедливо называют "энергетическими станциями" клетки, поскольку именно в этих органеллах в основном происходит улавливание энергии, поставляемой окислительными процессами. Митохондриальную систему сопряжения окислительных процессов с генерацией высокоэнергетического интермедиатора АТФ называют окислительным фосфорилированием.
Митохондрии имеют наружную мембрану, проницаемую для большинства метаболитов, и избирательно проницаемую внутреннюю мембрану с множеством складок (крист), выступающих в сторону матрикса (внутреннего пространства митохондрий). Наружная мембрана может быть удалена путем обработки дигитонином; она характеризуется наличием моноаминоксидазы и некоторых других ферментов (например, ацил-КоА-синтетазы, глицерофосфат-ацилтрансферазы, моноацилглицерофосфат-ацилтрансферазы, фосфолипазы А2). В межмембранном пространстве находятся аденилаткиназа и креатинкиназа. Во внутренней мембране локализован фосфолипид кардиолипин.
В матриксе находятся растворимые ферменты цикла лимонной кислоты и ферменты b-окисления жирных кислот, в связи с этим возникает необходимость в механизмах транспорта метаболитов и нуклеотидов через внутреннюю мембрану. Сукцинатдегидрогеназа локализована на внутренней поверхности внутренней митохондриальной мембраны, где она передает восстановительные эквиваленты дыхательной цепи на уровне убихинона (минуя первую окислительно-восстановительную петлю). 3-гидроксибутиратдегид рогеназа локализована на матриксной стороне внутренней митохондриальной мембраны. Глицерол-3-фосфат-дегидрогеназа находится на наружной поверхности внутренней мембраны, где она участвует в функционировании глицерофосфатного челночного механизма.[10,1993]
Этапы утилизации энергии питательных веществ.
Утилизация энергии питательных веществ - сложный процесс, который протекает в три стадии, согласно следующей схеме:
Схема 1. Стадии катаболизма питательных веществ.[1,1994]
На стадии 1 крупные молекулы полимеров распадаются на мономерные субъединицы: белки на аминокислоты, полисахариды на сахара, а жиры на жирные кислоты и холестеоролы. Этот предварительный процесс, называемый пищеварением, осуществляется главным образом вне клеток под действием ферментов, секретируемых в полость пищеварительного тракта. На стадии 2 образовавшиеся небольшие молекулы поступают в клетки и подвергаются дальнейшему расщеплению в цитоплазме. Большая часть углеродных и водородных атомов сахаров превращается в пируват, который, проникнув в митохондрии, образует там ацетильную группу химически активного соединения ацетилкофермента А (ацетил-СоА). Большое количество ацетил-СоА образуется также при окислении жирных кислот. На стадии 3 происходит полное расщепление ацетильной группы ацетил-СоА до СО2 и Н2О. Именно на этой заключительной стадии образуется большая часть АТФ. В серии сопряженных химических реакций больше половины той энергии, которую, согласно теоретическим расчетам, можно извлечь из углеводов и жиров при окислении их до Н2О и СО2, используется для осуществления энергетически невыгодной реакции Фн + АДФ ® АТФ. Поскольку остальная часть энергии, высвобождающейся при окислении, выделяется клеткой в виде тепла, результатом образования АТФ является общее возрастание неупорядоченности Вселенной, что полностью соответствует второму закону термодинамики.
Благодаря образованию АТФ энергия, первоначально извлеченная путем окисления из углеводов и жиров, преобразуется в более удобную концентрированную форму химической энергии. В растворе, находящемся во внутриклеточном пространстве типичной клетки, имеется примерно 1 млрд. молекул АТФ, гидролиз которых до АДФ и фосфата обеспечивает необходимой энергией множество энергетически невыгодных реакций.
Самым важным этапом стадии 2 катаболизма является гликолиз – последовательность реакций, приводящих к расщеплению глюкозы. При гликолизе молекула глюкозы, содержащая 6 атомов углерода, превращается в 2 молекулы пирувата, содержащие по 3 атома углерода каждая. Для такого превращения требуется 9 последовательных ферментативных реакций, в которых происходит образование ряда промежуточных фосфатсодержащих соединений. (см. рисунок 1.)
Логически рассуждая, последовательность реакций гликолиза можно разделить на три этапа: 1) в реакциях 1-4 (см.рисунок 1) глюкоза превращается в трехуглеродный альдегид глицеральдегид-3-фосфат (для этого превращения нужны две фосфатные группы, а необходимая энергия выделяется при гидролизе АТФ); 2) в реакциях 5-6 альдегидная группа каждой молекулы глицеральдегид-3-фосфата окисляется до карбоксильной, и выделяющаяся при этом энергия расходуется на синтез АТФ из АДФ и Фн; 3) в реакциях 7-9 те две молекулы фосфата, которые присоединились к сахару на первой стадии, переносятся обратно на АДФ, в результате чего образуется АТФ и компенсируются затраты АТФ на этапе 1.
Рисунок 1. Промежуточные продукты гликолиза.[1,1994]
Суммарный выход энергии при гликолизе сводится к синтезу двух молекул АТФ (на одну молекулу глюкозы), которые образовались в реакциях 5 и 6. Таким образом, данные реакции имеют решающее значение для гликолиза. Эти две реакции – единственные во всем процессе, в которых из Фн формируется высокоэнергетическая фосфатная связь. Суммарным результатом этих двух реакций является окисление сахарного альдегида в фосфоглицероловую кислоту, перенос Фн на АДФ с образованием высокоэнергетической связи АТФ и восстановления НАД+ до НАДН.
Для большинства клеток животных гликолиз предшествует стадии 3 катаболизма, т.к. образующаяся при гликолизе молочная кислота быстро поступает в митохондрии, где окисляется до СО2 и Н2О. Тем не менее у анаэробных организмов и тканей, способных работать в анаэробных условиях, гликолиз может стать основным источником клеточного АТФ. В этих случаях молекулы пирувата остаются в цитозоле и превращаются в лактат, который затем выводится из клетки. Дельнейшее превращение пирувата в этих энергодающих реакциях, называемых брожением, требуется для того, чтобы полностью использовать восстановительный потенциал, полученный в реакции 5 гликолиза, и таким путем регенерировать НАД+, необходимый для дальнейшего осуществления гликолиза.[1,1994]
Окислительное фосфорилирование.
Окислительное фосфорилирование позволяет аэробным организмам улавливать значительную долю потенциальной свободной энергии окисления субстратов. Возможное объяснение механизма окислительного фосфорилирования предлагает хемиосмотическая теория. Ряд лекарственных веществ (например, амобарбитал) и ядов (цианид, окись углерода) подавляют окислительное фосфорилирование, обычно с фатальными последствиями. Окислительное фосфорилирование является столь жизненно важным процессом, что нарушение его нормального хода несовместимо с жизнью. Этим можно объяснить, почему обнаружено лишь небольшое количество генетических нарушений, затрагивающих эту систему.
Хотя цикл лимонной кислоты составляет часть аэробного метаболизма, ни в одной из реакций этого цикла, приводящих к образованию НАДН и ФАДH2, молекулярный кислород не принимает прямого участия; это происходит только в завершающей серии катаболических реакций, протекающих на внутренней мембране. Почти вся энергия, получаемая на ранних этапах окисления от сжигания углеводов, жиров и других питательных веществ, вначале запасается в форме высокоэнергетических электронов, переносимых НАДН и ФАДН. Затем эти электроны взаимодействуют с молекулярным кислородом в дыхательной цепи. Taк как большое количество высвобождаемой энергии используется ферментами внутренней мембраны для синтеза АТФ из AДФ и Фн, эти последние реакции называют окислительным фосфорилированием.
Синтез АТФ в реакциях окислительного фосфорилирования, протекающих в дыхательной цепи, зависит от хемиосмотического процесса. Механизм этого процесса, впервые предложенный в 1961 году, позволил разрешить проблему, давно ставшую перед биологией клетки.
Раньше думали, что энергию для синтеза АТФ в дыхательной цепи обеспечивает такой же механизм, как и при субстратном фосфорилировании: предполагалось, что энергия окисления используется для образования высокоэнергетической связи между фосфатной группой и каким-то промежуточным соединением и, что превращение AДФ в АТФ осуществляется за счет энергии, выделяемой при разрыве этой связи. Однако, несмотря на интенсивные поиски, предполагаемый интермедиат не был обнаружен.
Согласно хемиосмотической гипотезе, вместо богатых энергией промежуточных продуктов существует прямая связь между процессами химическими («хеми...») и транспортными (осмотическими, от греческого osmos - толчок, давление) - хемиосмотическое сопряжение.
Хемиосмотическая гипотеза, предложенная в начале 60-х годов, включала четыре независимых постулата, касавшиеся функции митохондрий:
1. Митохондриальная дыхательная цепь, находящаяся во внутренней мембране, способна перемещать протоны; при прохождении электронов по дыхательной цепи происходит «откачивание» Н+ из матрикса.
2. Митохондриальный АТФ-синтетазный комплекс тоже перемещает протоны через внутреннюю мембрану. Поскольку этот процесс обратим, фермент может не только использовать энергию гидролиза АТФ для переноса Н + через мембрану, но при достаточно большом протонном градиенте протоны начинают «течь» через АТФ-синтетазу в обратном направлении, что сопровождается синтезом АТФ.
3. Внутренняя мембрана митохондрий непроницаема для Н +, ОН- и вообще всех анионов и катионов.
Похожие работы
... электронов, они способствуют использованию НАДН в других восстановительных реакциях, таких как синтез глутамата и глюконеогенез. Биологическое окисление и восстановление. Клеточным дыханием называют совокупность протекающих в каждой клетке ферментативных процессов, в результате которых молекулы углеводов, жирных кислот и аминокислот расщепляются в конечном счете до углекислоты и воды, а ...
... . 5.ПРОЦЕССЫ БИОЛОГИЧЕСКОЙ ОЧИСТКИ Сооружениям биологической очистки отводится главенствующая роль в общем комплексе сооружений канализационной очистной станции. В результате процессов биологической очистки сточная вода может быть очищена от многих органических и некоторых неорганических примесей. Процесс очистки осуществляет сложное сообщество микроорганизмов - бактерий, простейших, ряда ...

... Гензеляйт в 1932 г. вывели уравнения реакций синтеза мочевины, которые представлены в виде цикла, получившего в литературе название орнитинового цикла мочевинообразования Кребса. Следует указать, что в биохимии это была первая циклическая система метаболизма, описание которой почти на 5 лет опеределило открытие Г. Кребсом другого метаболического процесса – цикла трикарбоновых кислот. Дальнейшие ...
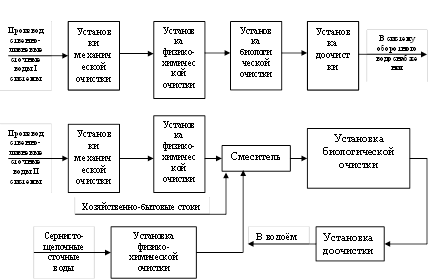

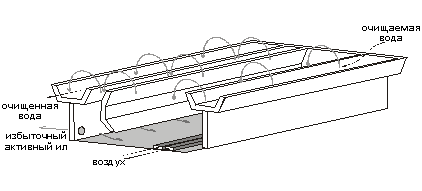
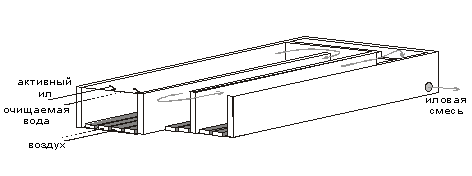
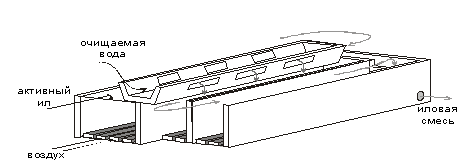
... труб на новые мембранные мелкопузырчатые аэраторы. Для достижения поставленных целей необходимо было решить следующие задачи: Ø Тщательно изучить теоретические основы технологии биохимической очистки сточных вод нефтеперерабатывающих предприятий; Ø Проанализировать имеющуюся технологическую схему очистки сточных вод на предприятии ООО "ЛУКОЙЛ-Пермнефтеоргсинтез"; Ø Выбрать ...
0 комментариев